
Comprehensive Approach for the Genetic Diagnosis of Patients with Waardenburg Syndrome

 , , , , ,
, , , , ,  , and
, and 
Abstract
:1. Introduction
2. Methods
2.1. WES Variant Prioritization Process
2.2. Copy Number Variants Analysis
2.2.1. WES RAW Analysis
2.2.2. MLPA Assays
3. Results
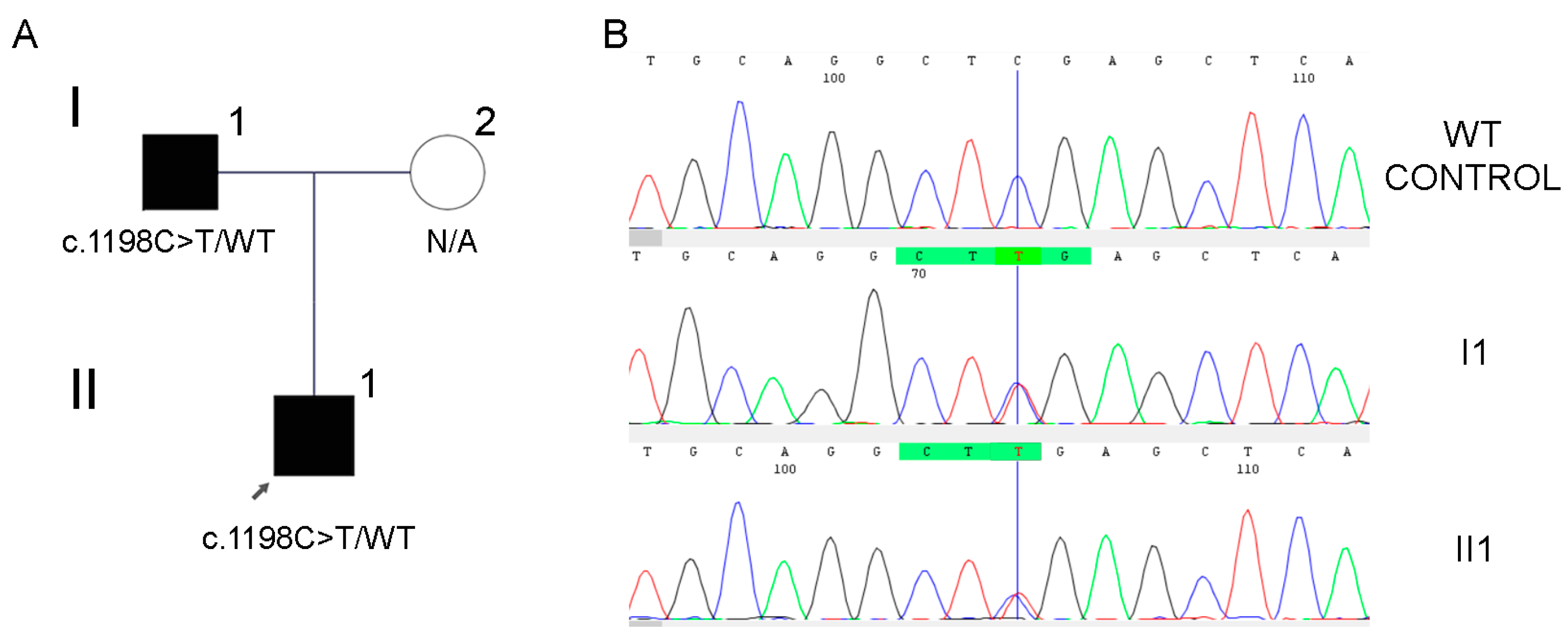
3.1. Case #1
3.2. Case #2
3.3. Case #3
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaman, A.; Capper, R.; Baddoo, W. Waardenburg Syndrome: More Common than You Think! Clin. Otolaryngol. 2015, 40, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Ahmed Jan, N.; Mui, R.K.; Masood, S. Waardenburg Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Saleem, M.D. Biology of Human Melanocyte Development, Piebaldism, and Waardenburg Syndrome. Pediatr. Dermatol. 2019, 36, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zheng, X.; Kong, X.; Chai, Y.; Wang, Y.; Qian, H.; Yang, B.; Wu, C.; Chu, J.; Yang, T. A Homozygous MITF Mutation Leads to Familial Waardenburg Syndrome Type 4. Am. J. Med. Genet. A 2019, 179, 243–248. [Google Scholar] [CrossRef]
- Ringer, J. Identification of Waardenburg Syndrome and the Management of Hearing Loss and Associated Sequelae: A Review for the Pediatric Nurse Practitioner. J. Pediatr. Health Care 2019, 33, 694–701. [Google Scholar] [CrossRef]
- Milunsky, J.M. Waardenburg Syndrome Type I. In GeneReviews; Adam, M.P., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington, Seattle: Seattle, WA, USA, 2001. [Google Scholar]
- Vanstrum, E.B.; Castellanos, C.X.; Ziltzer, R.S.; Ulloa, R.; Moen, R.; Choi, J.S.; Cortessis, V.K. Cochlear Implantation in Waardenburg Syndrome: Systematic Review and Meta-Analysis. Int. J. Pediatr. Otorhinolaryngol. 2023, 175, 111738. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Song, J.; He, C.; Cai, X.; Yuan, K.; Mei, L.; Feng, Y. Genetic Insights, Disease Mechanisms, and Biological Therapeutics for Waardenburg Syndrome. Gene Ther. 2022, 29, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Pingault, V.; Ente, D.; Dastot-Le Moal, F.; Goossens, M.; Marlin, S.; Bondurand, N. Review and Update of Mutations Causing Waardenburg Syndrome. Hum. Mutat. 2010, 31, 391–406. [Google Scholar] [CrossRef]
- Ren, S.; Chen, X.; Kong, X.; Chen, Y.; Wu, Q.; Jiao, Z.; Shi, H. Identification of Six Novel Variants in Waardenburg Syndrome Type II by next-Generation Sequencing. Mol. Genet. Genom. Med. 2020, 8, e1128. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Available online: https://www.who.int/ (accessed on 24 August 2023).
- Morton, C.C.; Nance, W.E. Newborn Hearing Screening--a Silent Revolution. N. Engl. J. Med. 2006, 354, 2151–2164. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid Isolation of High Molecular Weight Plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef]
- Buonfiglio, P.I.; Bruque, C.D.; Lotersztein, V.; Luce, L.; Giliberto, F.; Menazzi, S.; Francipane, L.; Paoli, B.; Goldschmidt, E.; Elgoyhen, A.B.; et al. Predicting Pathogenicity for Novel Hearing Loss Mutations Based on Genetic and Protein Structure Approaches. Sci. Rep. 2022, 12, 301. [Google Scholar] [CrossRef] [PubMed]
- Riggs, E.R.; Andersen, E.F.; Cherry, A.M.; Kantarci, S.; Kearney, H.; Patel, A.; Raca, G.; Ritter, D.I.; South, S.T.; Thorland, E.C.; et al. Technical Standards for the Interpretation and Reporting of Constitutional Copy-Number Variants: A Joint Consensus Recommendation of the American College of Medical Genetics and Genomics (ACMG) and the Clinical Genome Resource (ClinGen). Genet. Med. 2020, 22, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and Guidelines for the Interpretation of Sequence Variants: A Joint Consensus Recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Oza, A.M.; DiStefano, M.T.; Hemphill, S.E.; Cushman, B.J.; Grant, A.R.; Siegert, R.K.; Shen, J.; Chapin, A.; Boczek, N.J.; Schimmenti, L.A.; et al. Expert Specification of the ACMG/AMP Variant Interpretation Guidelines for Genetic Hearing Loss. Hum. Mutat. 2018, 39, 1593–1613. [Google Scholar] [CrossRef]
- Tavtigian, S.V.; Harrison, S.M.; Boucher, K.M.; Biesecker, L.G. Fitting a Naturally Scaled Point System to the ACMG/AMP Variant Classification Guidelines. Hum. Mutat. 2020, 41, 1734–1737. [Google Scholar] [CrossRef]
- Fowler, A. DECoN: A Detection and Visualization Tool for Exonic Copy Number Variants. Methods Mol. Biol. 2022, 2493, 77–88. [Google Scholar]
- den Dunnen, J.T.; Dalgleish, R.; Maglott, D.R.; Hart, R.K.; Greenblatt, M.S.; McGowan-Jordan, J.; Roux, A.-F.; Smith, T.; Antonarakis, S.E.; Taschner, P.E.M. HGVS Recommendations for the Description of Sequence Variants: 2016 Update. Hum. Mutat. 2016, 37, 564–569. [Google Scholar] [CrossRef]
- Jalilian, N.; Tabatabaiefar, M.A.; Bahrami, T.; Karbasi, G.; Bahramian, M.H.; Salimpoor, A.; Noori-Daloii, M.R. A Novel Pathogenic Variant in the MITF Gene Segregating with a Unique Spectrum of Ocular Findings in an Extended Iranian Waardenburg Syndrome Kindred. Mol. Syndromol. 2017, 8, 195–200. [Google Scholar] [CrossRef]
- Rauschendorf, M.-A.; Zimmer, A.D.; Laut, A.; Demmer, P.; Rösler, B.; Happle, R.; Sartori, S.; Fischer, J. Homozygous Intronic MITF Mutation Causes Severe Waardenburg Syndrome Type 2A. Pigment Cell Melanoma Res. 2019, 32, 85–91. [Google Scholar] [CrossRef]
- George, A.; Zand, D.J.; Hufnagel, R.B.; Sharma, R.; Sergeev, Y.V.; Legare, J.M.; Rice, G.M.; Scott Schwoerer, J.A.; Rius, M.; Tetri, L.; et al. Biallelic Mutations in MITF Cause Coloboma, Osteopetrosis, Microphthalmia, Macrocephaly, Albinism, and Deafness. Am. J. Hum. Genet. 2016, 99, 1388–1394. [Google Scholar] [CrossRef]
- Ideura, M.; Nishio, S.-Y.; Moteki, H.; Takumi, Y.; Miyagawa, M.; Sato, T.; Kobayashi, Y.; Ohyama, K.; Oda, K.; Matsui, T.; et al. Comprehensive Analysis of Syndromic Hearing Loss Patients in Japan. Sci. Rep. 2019, 9, 11976. [Google Scholar] [CrossRef]
- Li, W.; Mei, L.; Chen, H.; Cai, X.; Liu, Y.; Men, M.; Liu, X.Z.; Yan, D.; Ling, J.; Feng, Y. New Genotypes and Phenotypes in Patients with 3 Subtypes of Waardenburg Syndrome Identified by Diagnostic Next-Generation Sequencing. Neural Plast. 2019, 2019, 7143458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, Q.-D.; Zhao, L.-P.; Ma, J.; Zhang, T.-S.; Pang, J.-X.; Li, Y.-F.; Wang, M.-F.; Wang, A.-P.; Tang, L.; et al. A Novel Variant in MITF in a Child from Yunnan-Guizhou Plateau with Autosomal Dominant Inheritance of Nonsyndromic Hearing Loss: A Case Report. Mol. Med. Rep. 2018, 17, 6054–6058. [Google Scholar] [CrossRef] [PubMed]
- Thongpradit, S.; Jinawath, N.; Javed, A.; Noojarern, S.; Khongkraparn, A.; Tim-Aroon, T.; Lertsukprasert, K.; Suktitipat, B.; Jensen, L.T.; Wattanasirichaigoon, D. MITF Variants Cause Nonsyndromic Sensorineural Hearing Loss with Autosomal Recessive Inheritance. Sci. Rep. 2020, 10, 12712. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Shi, J.; Pang, X.; Hu, Y.; Wang, X.; Wu, H.; Yang, T. Molecular Etiology and Genotype-Phenotype Correlation of Chinese Han Deaf Patients with Type I and Type II Waardenburg Syndrome. Sci. Rep. 2016, 6, 35498. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Gao, X.; Su, Y.; Han, M.; Gao, B.; Guo, C.; Kang, D.; Huang, S.; Yuan, Y.; et al. Analysis of Genotype-Phenotype Relationships in 90 Chinese Probands with Waardenburg Syndrome. Hum. Genet. 2022, 141, 839–852. [Google Scholar] [CrossRef]
- Pandya, A.; Xia, X.J.; Landa, B.L.; Arnos, K.S.; Israel, J.; Lloyd, J.; James, A.L.; Diehl, S.R.; Blanton, S.H.; Nance, W.E. Phenotypic Variation in Waardenburg Syndrome: Mutational Heterogeneity, Modifier Genes or Polygenic Background? Hum. Mol. Genet. 1996, 5, 497–502. [Google Scholar] [CrossRef]
- Kingdom, R.; Wright, C.F. Incomplete Penetrance and Variable Expressivity: From Clinical Studies to Population Cohorts. Front. Genet. 2022, 13, 920390. [Google Scholar] [CrossRef]
- Velasco, G.; Francastel, C. Genetics Meets DNA Methylation in Rare Diseases. Clin. Genet. 2019, 95, 210–220. [Google Scholar] [CrossRef]
- Sun, F.; Xiao, M.; Ji, D.; Zheng, F.; Shi, T. Deciphering Potential Causative Factors for Undiagnosed Waardenburg Syndrome through Multi-Data Integration. Orphanet J. Rare Dis. 2024, 19, 226. [Google Scholar] [CrossRef]
- Song, J.; Feng, Y.; Acke, F.R.; Coucke, P.; Vleminckx, K.; Dhooge, I.J. Hearing Loss in Waardenburg Syndrome: A Systematic Review. Clin. Genet. 2016, 89, 416–425. [Google Scholar] [CrossRef]
- Milunsky, J.M.; Maher, T.A.; Ito, M.; Milunsky, A. The Value of MLPA in Waardenburg Syndrome. Genet. Test. 2007, 11, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Bondurand, N.; Dastot-Le Moal, F.; Stanchina, L.; Collot, N.; Baral, V.; Marlin, S.; Attie-Bitach, T.; Giurgea, I.; Skopinski, L.; Reardon, W.; et al. Deletions at the SOX10 Gene Locus Cause Waardenburg Syndrome Types 2 and 4. Am. J. Hum. Genet. 2007, 81, 1169–1185. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jiang, L.; Xie, Z.; Mei, L.; He, C.; Hu, Z.; Xia, K.; Feng, Y. Novel Mutations of PAX3, MITF, and SOX10 Genes in Chinese Patients with Type I or Type II Waardenburg Syndrome. Biochem. Biophys. Res. Commun. 2010, 397, 70–74. [Google Scholar] [CrossRef]
- Siomou, E.; Manolakos, E.; Petersen, M.; Thomaidis, L.; Gyftodimou, Y.; Orru, S.; Papoulidis, I. A 725 Kb Deletion at 22q13.1 Chromosomal Region Including SOX10 Gene in a Boy with a Neurologic Variant of Waardenburg Syndrome Type 2. Eur. J. Med. Genet. 2012, 55, 641–645. [Google Scholar] [CrossRef]
- Chen, S.; Francioli, L.C.; Goodrich, J.K.; Collins, R.L.; Kanai, M.; Wang, Q.; Alföldi, J.; Watts, N.A.; Vittal, C.; Gauthier, L.D.; et al. A Genomic Mutational Constraint Map Using Variation in 76,156 Human Genomes. Nature 2024, 625, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of Protein-Coding Genetic Variation in 60,706 Humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef]
- Talevich, E.; Shain, A.H.; Botton, T.; Bastian, B.C. CNVkit: Genome-Wide Copy Number Detection and Visualization from Targeted DNA Sequencing. PLoS Comput. Biol. 2016, 12, e1004873. [Google Scholar] [CrossRef]
- McCarroll, S.A.; Altshuler, D.M. Copy-Number Variation and Association Studies of Human Disease. Nat. Genet. 2007, 39, S37–S42. [Google Scholar] [CrossRef]
- Singh, A.K.; Olsen, M.F.; Lavik, L.A.S.; Vold, T.; Drabløs, F.; Sjursen, W. Detecting Copy Number Variation in next Generation Sequencing Data from Diagnostic Gene Panels. BMC Med. Genom. 2021, 14, 214. [Google Scholar] [CrossRef]
- Kiani, R.; Gangadharan, S.K.; Miller, H. Case Report: Association of Waardenburg Syndrome with Intellectual Disability, Autistic Spectrum Disorder and Unprovoked Aggressive Outbursts: A New Behavioural Phenotype? Br. J. Dev. Disabil. 2007, 53, 53–62. [Google Scholar] [CrossRef]
- George, F.S.A.; Sam, L.E.; Tanwar, M.; Wall, L. Association of Autism Spectrum Disorder with Waardenburg Syndrome in a Toddler. BMJ Case Rep. 2023, 16, e254741. [Google Scholar] [CrossRef] [PubMed]
- Cantani, A.; Bamonte, G.; Tacconi, M.L. Mental Retardation and EEG Abnormalities in Waardenburg’s Syndrome: Two Case Reports (EEG Anomalies in Waardenburg’s Syndrome). Padiatr. Padol. 1989, 24, 137–140. [Google Scholar]
- Xu, G.Y.; Hao, Q.Q.; Zhong, L.L.; Ren, W.; Yan, Y.; Liu, R.Y.; Li, J.N.; Guo, W.W.; Zhao, H.; Yang, S.M. 10 mutation is relevant to inner ear malformation in patients with Waardenburg syndrome. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 2016, 51, 832–837. [Google Scholar] [PubMed]
- Elmaleh-Bergès, M.; Baumann, C.; Noël-Pétroff, N.; Sekkal, A.; Couloigner, V.; Devriendt, K.; Wilson, M.; Marlin, S.; Sebag, G.; Pingault, V. Spectrum of Temporal Bone Abnormalities in Patients with Waardenburg Syndrome and SOX10 Mutations. AJNR Am. J. Neuroradiol. 2013, 34, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Barnett, C.P.; Mendoza-Londono, R.; Blaser, S.; Gillis, J.; Dupuis, L.; Levin, A.V.; Chiang, P.W.; Spector, E.; Reardon, W. Aplasia of Cochlear Nerves and Olfactory Bulbs in Association with SOX10 Mutation. Am. J. Med. Genet. A 2009, 149A, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Lovett, A.; Eastwood, M.; Metcalfe, C.; Muzaffar, J.; Monksfield, P.; Bance, M. Outcomes of Cochlear Implantation in Early-Deafened Patients with Waardenburg Syndrome: A Systematic Review and Narrative Synthesis. Laryngoscope Investig. Otolaryngol. 2023, 8, 1094–1107. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Gene | WS Subtype | Genotype (RefSeq NM) | Proband’s Clinical Features | Classification According to ACMG and HL-VCEP |
|---|---|---|---|---|---|
| 1 | MITF | Type II | c.1198C>T (NM_001354604.2) p.(Arg400*) | White forelock Congenital profound sensorineural hearing loss Cochlear implants | Pathogenic (PM2_Supporting, PVS1_Strong, PS4_Supporting, PP1_Strong, PP4) 11 points = 11 P-0B |
| 2 | PAX3 | Type I | del-Ex5 # (NM_181458.4) | Dystopia canthorum Hypoplastic blue eyes Congenital profound sensorineural hearing loss Developmental disorder | Pathogenic Total score: 1.3 |
| 3 | EIF3L, MICALL1, C22orf23, POLR2F, MIR6820, MIR4534, SOX10 | Type II | seq[GRCh38] del(22)(22p13.1) NC_000022.11:g.(?_37849420)_(37988853_?)del | Heterochromia iridis Congenital profound sensorineural hearing loss | Pathogenic Total score: 1.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buonfiglio, P.I.; Izquierdo, A.; Pace, M.V.; Grinberg, S.; Lotersztein, V.; Brun, P.; Bruque, C.D.; Elgoyhen, A.B.; Dalamón, V. Comprehensive Approach for the Genetic Diagnosis of Patients with Waardenburg Syndrome. J. Pers. Med. 2024, 14, 906. https://doi.org/10.3390/jpm14090906
Buonfiglio PI, Izquierdo A, Pace MV, Grinberg S, Lotersztein V, Brun P, Bruque CD, Elgoyhen AB, Dalamón V. Comprehensive Approach for the Genetic Diagnosis of Patients with Waardenburg Syndrome. Journal of Personalized Medicine. 2024; 14(9):906. https://doi.org/10.3390/jpm14090906
Chicago/Turabian StyleBuonfiglio, Paula Inés, Agustín Izquierdo, Mariela Vanina Pace, Sofia Grinberg, Vanesa Lotersztein, Paloma Brun, Carlos David Bruque, Ana Belén Elgoyhen, and Viviana Dalamón. 2024. "Comprehensive Approach for the Genetic Diagnosis of Patients with Waardenburg Syndrome" Journal of Personalized Medicine 14, no. 9: 906. https://doi.org/10.3390/jpm14090906