
Biofilm Development on Carbon Steel by Iron Reducing Bacterium Shewanella putrefaciens and Their Role in Corrosion



Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Bacterial Culture
2.2. Test Solutions Preparation
2.3. Carbon Steel Sample Preparation
2.4. Iron Quantification
2.5. Microscopic and Biofilm Cross-Sectional Analysis
2.5.1. Development of Biofilms on Carbon Steel for Microscopic Imaging and FIB-SEM Analysis
2.5.2. Biofilm Fixation and Environmental Scanning Electron Microscopy (ESEM) Imaging
2.5.3. Focused Ion Beam—Scanning Electron Microscopy (FIB-SEM)
2.5.4. Attenuated Total Reflectance—Fourier Transform Infrared (ATR-FTIR) Spectroscopy
2.6. Electrochemical Measurements
3. Results and Discussion
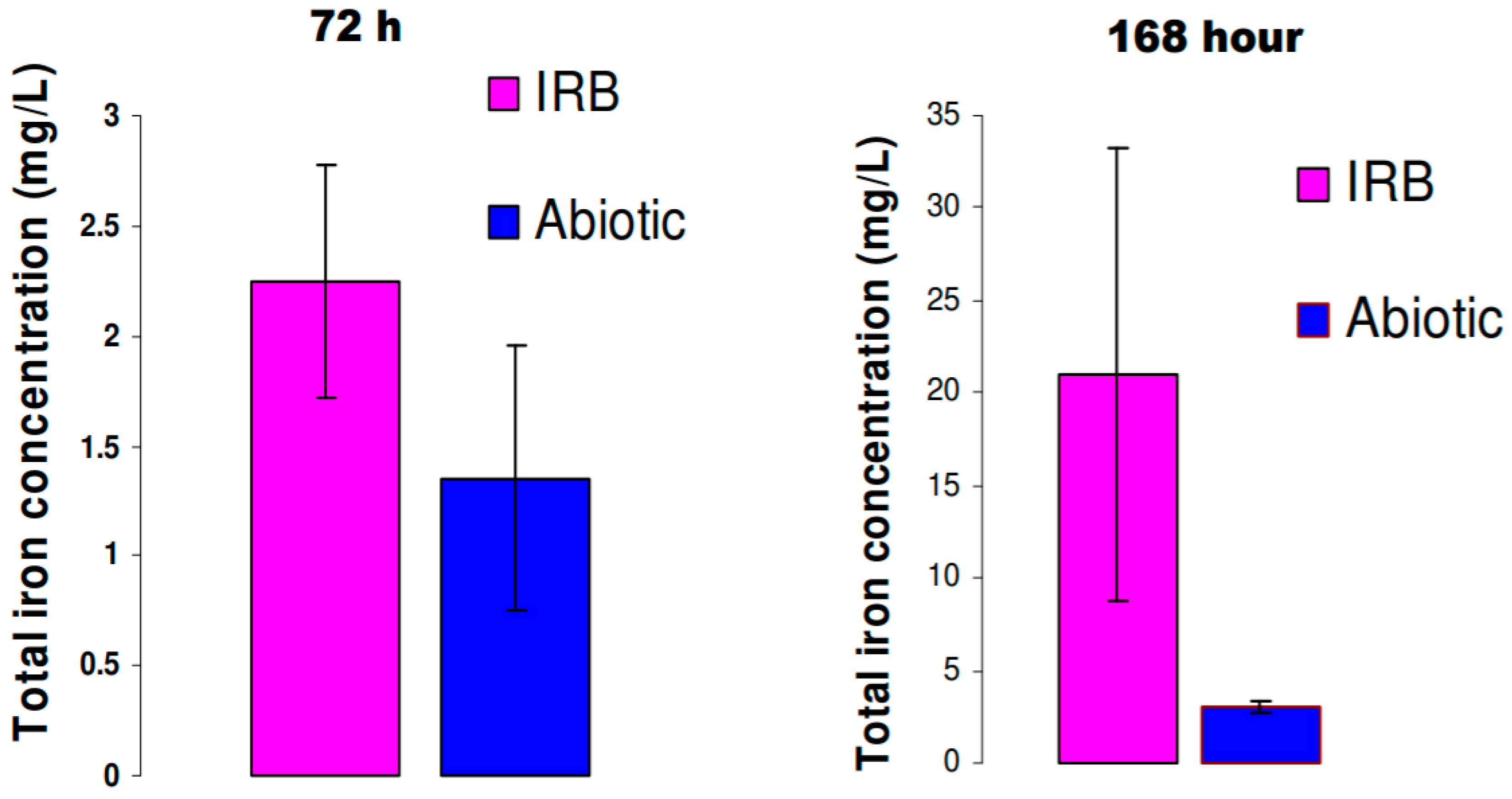
3.1. Iron and Sulfide Measurements in Solution
3.2. Biofilm Development and Electrochemical Characterization of Carbon Steel in Biotic Postgate C Solution
3.2.1. Biofilm Development in Commonly Used Postgate C Solution with IRB
3.2.2. Electrochemical Behaviour of Carbon STEEL in Postgate C Medium Inoculated with IRB
3.3. Effect of Removal of Yeast Extract from Postgate C (Modified Postgate C) on IRB Activity and Biofilm Development
3.3.1. Biofilm Development in Modified Postgate C Solution with IRB
3.3.2. Electrochemical Behaviour of Steel in Modified Postgate C Medium with IRB
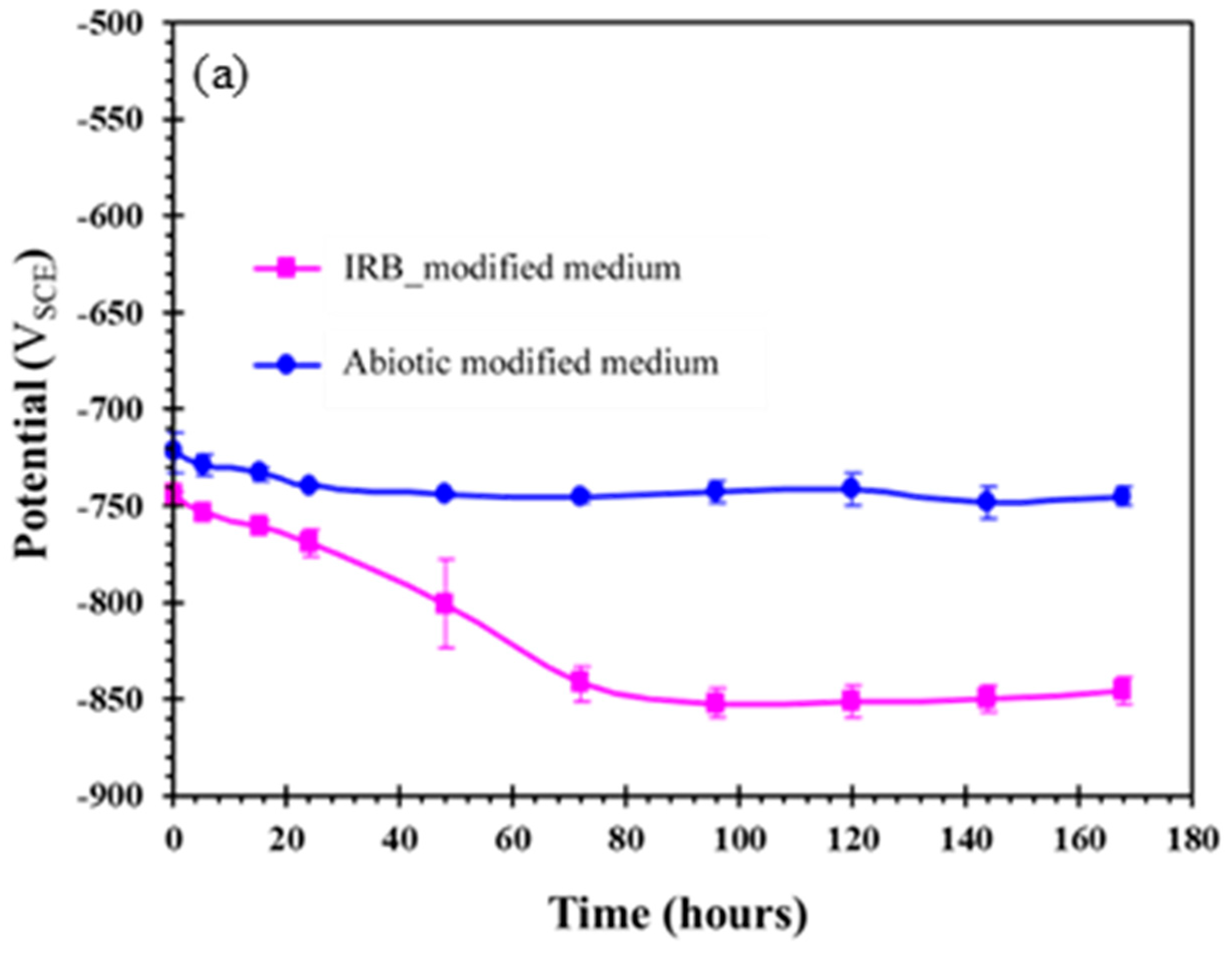
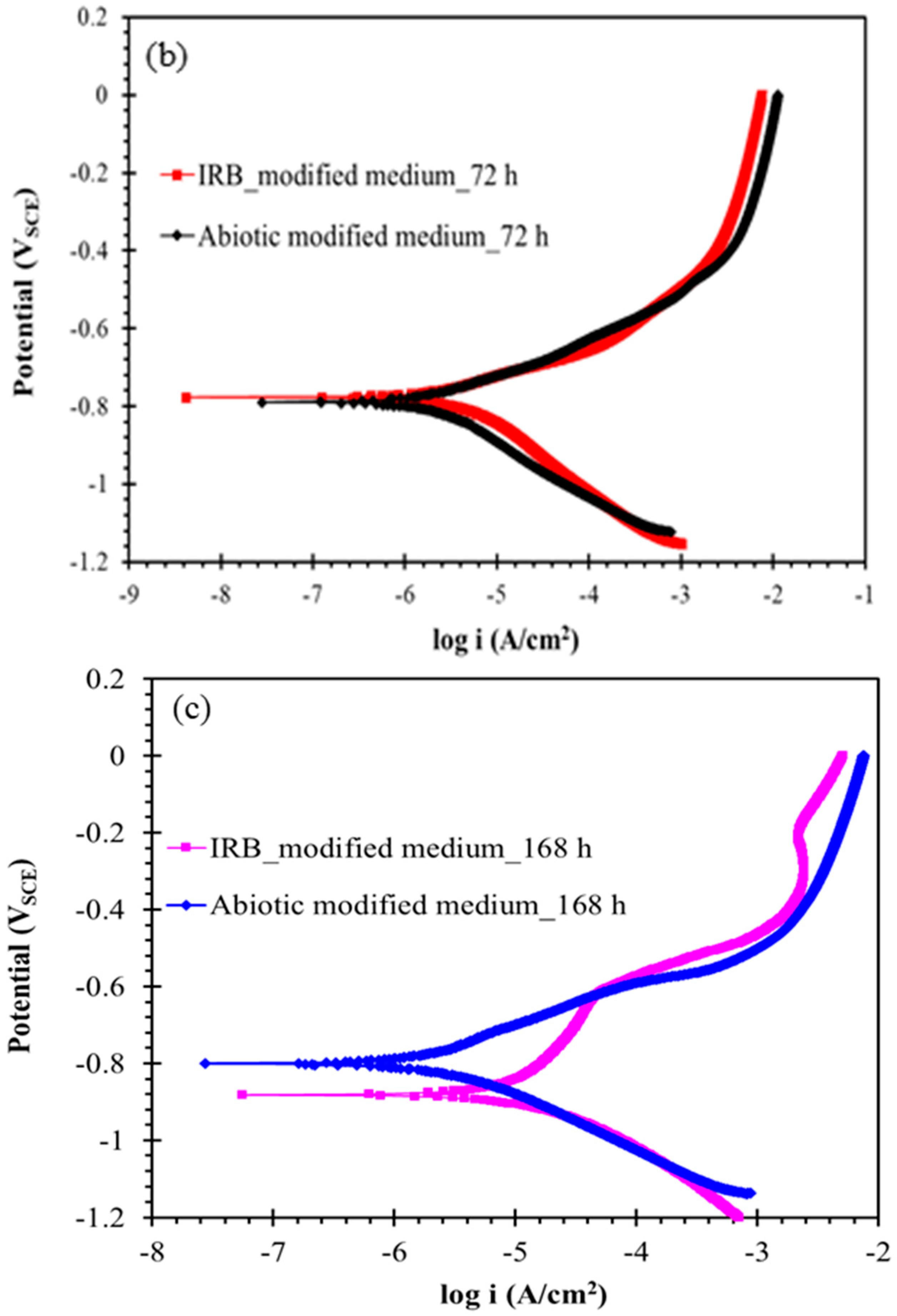
3.4. Biofilm Development and Electrochemical Characterization of Carbon Steel in Biotic Postgate C Solution without Organic Nutrients (i.e., Inorganic Medium)
3.4.1. Biofilm Development on Carbon Steel Exposed to IRB in Inorganic Postgate C Solution
3.4.2. Electrochemical Behaviour Carbon Steel Exposed to Inorganic Postgate C Solution with IRB
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dawuda, A.-W.; Taleb-Berrouane, M.; Khan, F. A probabilistic model to estimate microbiologically influenced corrosion rate. Process Saf. Environ. Prot. 2021, 148, 908–926. [Google Scholar] [CrossRef]
- Javaherdashti, R. Microbiologically influenced corrosion (MIC). In Microbiologically Influenced Corrosion; Springer: London, UK, 2017; pp. 29–71. [Google Scholar]
- Little, B.J.; Lee, J.S. Microbiologically Influenced Corrosion; John Wiley & Sons: Hoboken, NJ, USA, 2007; Volume 3. [Google Scholar]
- Sequeira, C.A.; Tiller, A. Microbial Corrosion-1; Elsevier Applied Science: London, UK, 1988. [Google Scholar]
- Padilla-Viveros, A.; Garcia-Ochoa, E.; Alazard, D. Comparative electrochemical noise study of the corrosion process of carbon steel by the sulfate-reducing bacterium Desulfovibrio alaskensis under nutritionally rich and oligotrophic culture conditions. Electrochim. Acta 2006, 51, 3841–3847. [Google Scholar] [CrossRef]
- Sun, Z.; Moradi, M.; Chen, Y.; Bagheri, R.; Guo, P.; Yang, L.; Song, Z.; Xu, C. Simulation of the marine environment using bioreactor for investigation of 2507 duplex stainless steel corrosion in the presence of marine isolated Bacillus Vietnamensis bacterium. Mater. Chem. Phys. 2018, 208, 149–156. [Google Scholar] [CrossRef]
- Enning, D.; Garrelfs, J. Corrosion of Iron by Sulfate-Reducing Bacteria: New Views of an Old Problem. Appl. Environ. Microbiol. 2014, 80, 1226–1236. [Google Scholar] [CrossRef]
- Little, B.J.; Lee, J.S. Microbiologically influenced corrosion. In Kirk-Othmer Encyclopedia of Chemical Technology; Wiley: Hoboken, NJ, USA, 2000; pp. 1–42. [Google Scholar]
- Usher, K.M.; Kaksonen, A.H.; Cole, I.; Marney, D. Critical review: Microbially influenced corrosion of buried carbon steel pipes. Int. Biodeterior. Biodegrad. 2014, 93, 84–106. [Google Scholar] [CrossRef]
- Herrera, L.K.; Videla, H.A. Role of iron-reducing bacteria in corrosion and protection of carbon steel. Int. Biodeterior. Biodegrad. 2009, 63, 891–895. [Google Scholar] [CrossRef]
- Gaylarde, C.C.; Videla, H.A. Localised corrosion induced by a marine vibrio. Int. Biodeterior. 1987, 23, 91–104. [Google Scholar] [CrossRef]
- Little, B.; Wagner, P.; Hart, K.; Ray, R.; Lavoie, D.; Nealson, K.; Aguilar, C. The role of biomineralization in microbiologically influenced corrosion. Biogeochemistry 1998, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kalajahi, S.T.; Rasekh, B.; Yazdian, F.; Neshati, J.; Taghavi, L. Corrosion behaviour of X60 steel in the presence of sulphate-reducing bacteria (SRB) and iron-reducing bacteria (IRB) in seawater. Corros. Eng. Sci. Technol. 2021, 56, 543–552. [Google Scholar] [CrossRef]
- Perego, P.; Fabiano, B. Microbiological Induced Corrosion and Inhibition. In Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 1–21. [Google Scholar]
- Dubiel, M.; Hsu, C.H.; Chien, C.C.; Mansfeld, F.; Newman, D.K. Microbial Iron Respiration Can Protect Steel from Corrosion. Appl. Environ. Microbiol. 2002, 68, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.K.; Newman, D.K. Microbial iron respiration: Impacts on corrosion processes. Appl. Microbiol. Biotechnol. 2003, 62, 134–139. [Google Scholar] [CrossRef] [PubMed]
- AlAbbas, F.M.; Bhola, S.M.; Spear, J.R.; Olson, D.L.; Mishra, B. The shielding effect of wild type iron reducing bacterial flora on the corrosion of linepipe steel. Eng. Fail. Anal. 2013, 33, 222–235. [Google Scholar] [CrossRef]
- Sharma, M.; Voordouw, G. MIC detection and assessment: A holistic approach. In Microbiologically Influenced Corrosion in the Upstream Oil and Gas Industry; CRC Press: Boca Raton, FL, USA, 2017; pp. 177–212. [Google Scholar]
- Videla, H.A.; Herrera, L. Biocorrosion in oil recovery systems: Prevention and protection-An update. Edición Espec. 2007, 30, 272–279. [Google Scholar]
- Kim, H.J.; Park, H.S.; Hyun, M.S.; Chang, I.S.; Kim, M.; Kim, B.H. A mediator-less microbial fuel cell using a metal reducing bacterium, Shewanella putrefaciens. Enzym. Microb. Technol. 2002, 30, 145–152. [Google Scholar] [CrossRef]
- O’Loughlin, E.J. Effects of electron transfer mediators on the bioreduction of lepidocrocite (γ-FeOOH) by Shewanella putrefaciens CN32. Environ. Sci. Technol. 2008, 42, 6876–6882. [Google Scholar] [CrossRef]
- Glasauer, S.; Weidler, P.G.; Langley, S.; Beveridge, T.J. Controls on Fe reduction and mineral formation by a subsurface bacterium. Geochim. Cosmochim. Acta 2003, 67, 1277–1288. [Google Scholar] [CrossRef]
- Igarashi, K.; Kato, S. Reductive Transformation of Fe (III) (oxyhydr)Oxides by Mesophilic Homoacetogens in the Genus Sporomusa. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Frankel, R.B.; Bazylinski, D.A. Biologically Induced Mineralization by Bacteria. Rev. Mineral. Geochem. 2003, 54, 95–114. [Google Scholar] [CrossRef]
- Fredrickson, J.K.; Zachara, J.M.; Kennedy, D.W.; Dong, H.; Onstott, T.C.; Hinman, N.W.; Li, S.-M. Biogenic iron mineralization accompanying the dissimilatory reduction of hydrous ferric oxide by a groundwater bacterium. Geochim. Cosmochim. Acta 1998, 62, 3239–3257. [Google Scholar] [CrossRef]
- Postgate, J.R. The Sulphate-Reducing Bacteria; CUP Archive: Cambridge, UK, 1979. [Google Scholar]
- Al-Saadi, S.; Banerjee, P.C.; Raman, R.S. Corrosion of bare and silane-coated mild steel in chloride medium with and without sulphate reducing bacteria. Prog. Org. Coat. 2017, 111, 231–239. [Google Scholar] [CrossRef]
- Al-Saadi, S.; Raman, R.S.; Panter, C. A Two-Step Silane Coating Incorporated with Quaternary Ammonium Silane for Mitigation of Microbial Corrosion of Mild Steel. ACS Omega 2021, 6, 16913–16923. [Google Scholar] [CrossRef] [PubMed]
- Al-Saadi, S.; Raman, R.S. A long aliphatic chain functional silane for corrosion and microbial corrosion resistance of steel. Prog. Org. Coat. 2018, 127, 27–36. [Google Scholar] [CrossRef]
- Cheng, Y.F.; Rairdan, B.R.; Luo, J.L. Features of electrochemical noise generated during pitting of inhibited A516-70 carbon steel in chloride solutions. J. Appl. Electrochem. 1998, 28, 1371–1375. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Blankemeier, A.R. Characterization of Pseudomonas Fluorescens Biofilm. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2011. [Google Scholar]
- Simpson, W.J. Isolation and Characterisation of Thermophilic Anaerobes from Bass Strait oil Production Waters. Master’s Thesis, Monash University, Clayton, Australia, 1999. [Google Scholar] [CrossRef]
- Hamilton, W.A. Microbially Influenced Corrosion as a Model System for the Study of Metal Microbe Interactions: A Unifying Electron Transfer Hypothesis. Biofouling 2003, 19, 65–76. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, E.J.; Boyanov, M.I.; Gorski, C.A.; Scherer, M.M.; Kemner, K.M. Effects of Fe(III) Oxide Mineralogy and Phosphate on Fe(II) Secondary Mineral Formation during Microbial Iron Reduction. Minerals 2021, 11, 149. [Google Scholar] [CrossRef]
- Bonneville, S.; Behrends, T.; Van Cappellen, P.; Hyacinthe, C.; Röling, W.F. Reduction of Fe(III) colloids by Shewanella putrefaciens: A kinetic model. Geochim. Cosmochim. Acta 2006, 70, 5842–5854. [Google Scholar] [CrossRef]
- Lies, D.P.; Hernandez, M.E.; Kappler, A.; Mielke, R.E.; Gralnick, J.A.; Newman, D.K. Shewanella oneidensis MR-1 Uses Overlapping Pathways for Iron Reduction at a Distance and by Direct Contact under Conditions Relevant for Biofilms. Appl. Environ. Microbiol. 2005, 71, 4414–4426. [Google Scholar] [CrossRef]
- Fuller, S.J.; McMillan, D.; Renz, M.B.; Schmidt, M.; Burke, I.T.; Stewart, D.I. Extracellular Electron Transport-Mediated Fe (III) Reduction by a Community of Alkaliphilic Bacteria That Use Flavins as Electron Shuttles. Appl. Environ. Microbiol. 2014, 80, 128–137. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P.; Lonergan, D.J. Hydrogen and Formate Oxidation Coupled to Dissimilatory Reduction of Iron or Manganese by Alteromonas putrefaciens. Appl. Environ. Microbiol. 1989, 55, 700–706. [Google Scholar] [CrossRef]
- Pinney, N.; Morgan, D. Ab initio study of structurally bound water at cation vacancy sites in Fe- and Al-oxyhydroxide materials. Geochim. Cosmochim. Acta 2013, 114, 94–111. [Google Scholar] [CrossRef]
- Ona Nguema, G.; Abdelmoula, M.; Jorand, F.; Benali, O.; Géhin, A.; Block, J.C.; Génin, J.M. Microbial reduction of lepidocrocite gamma-FeOOH by Shewanella putrefaciens: The formation of green rust. In Industrial Applications of the Mössbauer Effect; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Bosch, A.; Serra, D.; Prieto, C.; Schmitt, J.; Naumann, D.; Yantorno, O. Characterization of Bordetella pertussis growing as biofilm by chemical analysis and FT-IR spectroscopy. Appl. Microbiol. Biotechnol. 2005, 71, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, J.; Flemming, H.-C. FTIR-spectroscopy in microbial and material analysis. Int. Biodeterior. Biodegrad. 1998, 41, 1–11. [Google Scholar] [CrossRef]
- Socrates, G. Infrared and Raman Characteristic Group Frequencies Tables and Charts; John Wiley & Sons Ltd.: Chichester, UK, 2001. [Google Scholar]
- Furukawa, Y.; Dale, J.R. The surface properties of Shewanella putrefaciens 200 and S. oneidensis MR-1: The effect of pH and terminal electron acceptors. Geochem. Trans. 2013, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Omoike, A.; Chorover, J.; Kwon, K.D.; Kubicki, J. DAdhesion of Bacterial Exopolymers to α-FeOOH: Inner-Sphere Complexation of Phosphodiester Groups. Langmuir 2004, 20, 11108–11114. [Google Scholar] [CrossRef]
- McGill, I.R.; McEnaney, B.; Smith, D.C. Crystal structure of green rust formed by corrosion of cast iron. Nature 1976, 259, 200–201. [Google Scholar] [CrossRef]
- Refait, P.; Memet, J.-B.; Bon, C.; Sabot, R.; Génin, J.-M. Formation of the Fe (II)–Fe (III) hydroxysulphate green rust during marine corrosion of steel. Corros. Sci. 2003, 45, 833–845. [Google Scholar] [CrossRef]
- Meyers, R.A. Encyclopedia of Molecular Cell Biology and Molecular Medicine; Wiley Online Library: Weinheim, Germany, 2004. [Google Scholar]
- Abdel-Khalek, N.A.; Selim, K.A.; Amir, A.; Farahat, M.M.; El-Hendawy, H.H. Enhancement of Selective Bioflocculation of (Quartz- PyrolusiteHematite) System Using Paenibacillus polymyxa Bacteria. Bio Tech. 2015, 89, 36712–36717. [Google Scholar]
- Natarajan, K.; Deo, N. Role of bacterial interaction and bioreagents in iron ore flotation. Int. J. Miner. Process. 2001, 62, 143–157. [Google Scholar] [CrossRef]
- Beech, I.B.; Sunner, J. Biocorrosion: Towards understanding interactions between biofilms and metals. Curr. Opin. Biotechnol. 2004, 15, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Sherar, B.W.A.; Keech, P.G.; Qin, Z.; King, F.; Shoesmith, D.W. Nominally Anaerobic Corrosion of Carbon Steel in Near-Neutral pH Saline Environments. Corrosion 2010, 66, 045001–045001-11. [Google Scholar] [CrossRef]
- Nealson, K.H.; Myers, C.R. Iron reduction by bacteria: A potential role in the genesis of banded iron formations. Am. J. Sci. 1990, 290, 35–45. [Google Scholar]
- Berthelin, J.; Ona-Nguema, G.; Stemmler, S.; Quantin, C.; Abdelmoula, M.; Jorand, F. Bioreduction of ferric species and biogenesis of green rusts in soils. Comptes Rendus Geosci. 2006, 338, 447–455. [Google Scholar] [CrossRef]
- Hedrich, S.; Johnson, D.B. Aerobic and anaerobic oxidation of hydrogen by acidophilic bacteria. FEMS Microbiol. Lett. 2013, 349, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Gram, L. Siderophore-Mediated Iron Sequestering by Shewanella putrefaciens. Appl. Environ. Microbiol. 1994, 60, 2132–2136. [Google Scholar] [CrossRef] [PubMed]
- Kane, R.D.; Cayard, M.S. Roles of H2S in the Behavior of Engineering Alloys: A Review of Literature and Experience. In Proceedings of the Corrosion-National Association OF Corrosion Engineers Annual Conference, San Diego, CA, USA, 22–27 March 1998. [Google Scholar]
- Jack, T.R. Biological Corrosion Failures; ASM International: Russell, OH, USA, 2002. [Google Scholar]
- Pickering, H.W. Important Early Developments and Current Understanding of the IR Mechanism of Localized Corrosion. J. Electrochem. Soc. 2003, 150, K1. [Google Scholar] [CrossRef]
- Ghahari, S.M. In Situ Synchrotron X-ray Characterisation and Modelling of Pitting Corrosion of Stainless Steel. Ph.D. Thesis, University of Birmingham, Birmingham, UK, 2012. [Google Scholar]
- Esaklul, K.A. Handbook of Case Histories in Failure Analysis; ASM International: Almere, The Netherlands, 1992. [Google Scholar]
- Hansel, C.M.; Benner, S.G.; Neiss, J.; Dohnalkova, A.; Kukkadapu, R.K.; Fendorf, S. Secondary mineralization pathways induced by dissimilatory iron reduction of ferrihydrite under advective flow. Geochim. Cosmochim. Acta 2003, 67, 2977–2992. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemicals | Postgate C | Modified Postgate C | Inorganic Postgate C |
|---|---|---|---|
| Na2SO4 a | 4.5 | 4.5 | 4.5 |
| CaCl2·2H2O a | 0.06 | 0.06 | 0.06 |
| Lactic acid b | 4.8 | 4.8 | - |
| Sodium citrate a | 0.3 | 0.3 | - |
| NH4Cl a | 1.0 | 1.0 | 1.0 |
| K2HPO4 a | 0.5 | 0.5 | 0.5 |
| MgSO4·7H2O a | 2.0 | 2.0 | 2.0 |
| Yeast extract a | 1.0 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Welikala, S.; Al-Saadi, S.; Gates, W.P.; Panter, C.; Raman, R.K.S. Biofilm Development on Carbon Steel by Iron Reducing Bacterium Shewanella putrefaciens and Their Role in Corrosion. Metals 2022, 12, 1005. https://doi.org/10.3390/met12061005
Welikala S, Al-Saadi S, Gates WP, Panter C, Raman RKS. Biofilm Development on Carbon Steel by Iron Reducing Bacterium Shewanella putrefaciens and Their Role in Corrosion. Metals. 2022; 12(6):1005. https://doi.org/10.3390/met12061005
Chicago/Turabian StyleWelikala, Sachie, Saad Al-Saadi, Will P. Gates, Christopher Panter, and R. K. Singh Raman. 2022. "Biofilm Development on Carbon Steel by Iron Reducing Bacterium Shewanella putrefaciens and Their Role in Corrosion" Metals 12, no. 6: 1005. https://doi.org/10.3390/met12061005
APA StyleWelikala, S., Al-Saadi, S., Gates, W. P., Panter, C., & Raman, R. K. S. (2022). Biofilm Development on Carbon Steel by Iron Reducing Bacterium Shewanella putrefaciens and Their Role in Corrosion. Metals, 12(6), 1005. https://doi.org/10.3390/met12061005