
The Yorktown Formation: Improved Stratigraphy, Chronology, and Paleoclimate Interpretations from the U.S. Mid-Atlantic Coastal Plain



Abstract
:1. Introduction
2. Yorktown Formation Stratigraphy
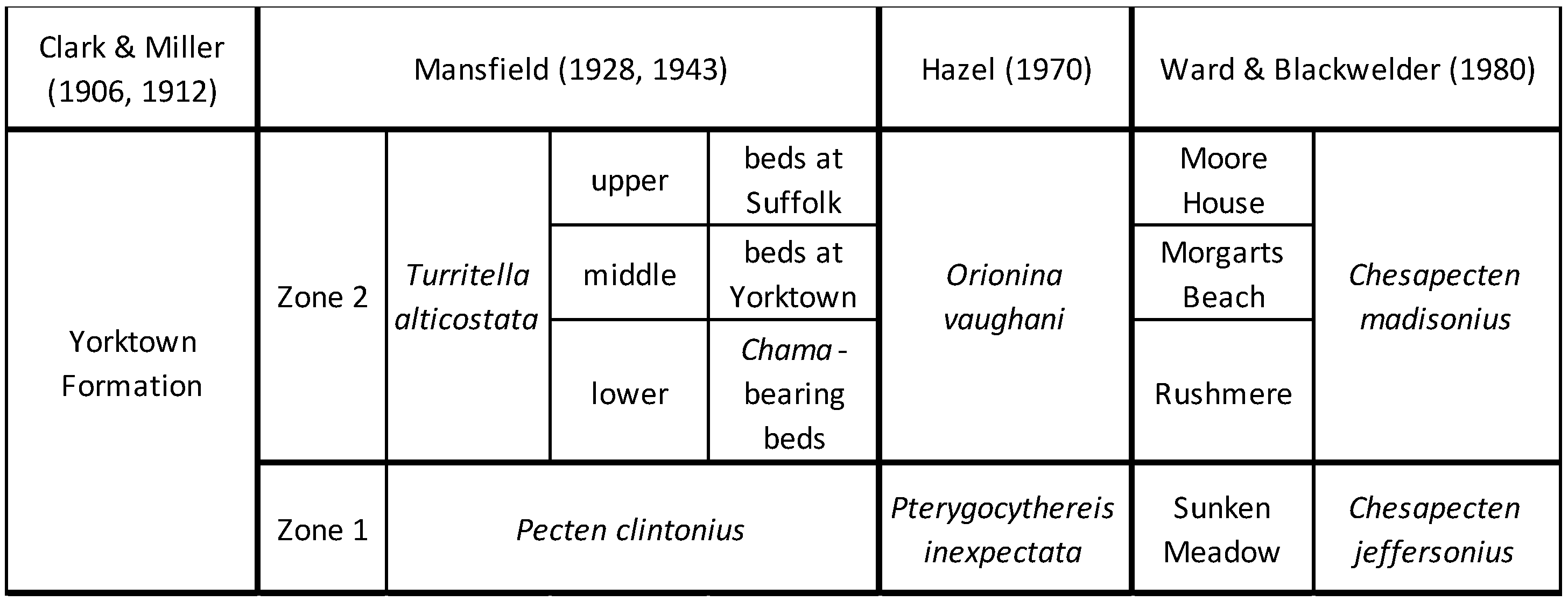
2.1. Previous Stratigraphic Work
2.2. Chronology
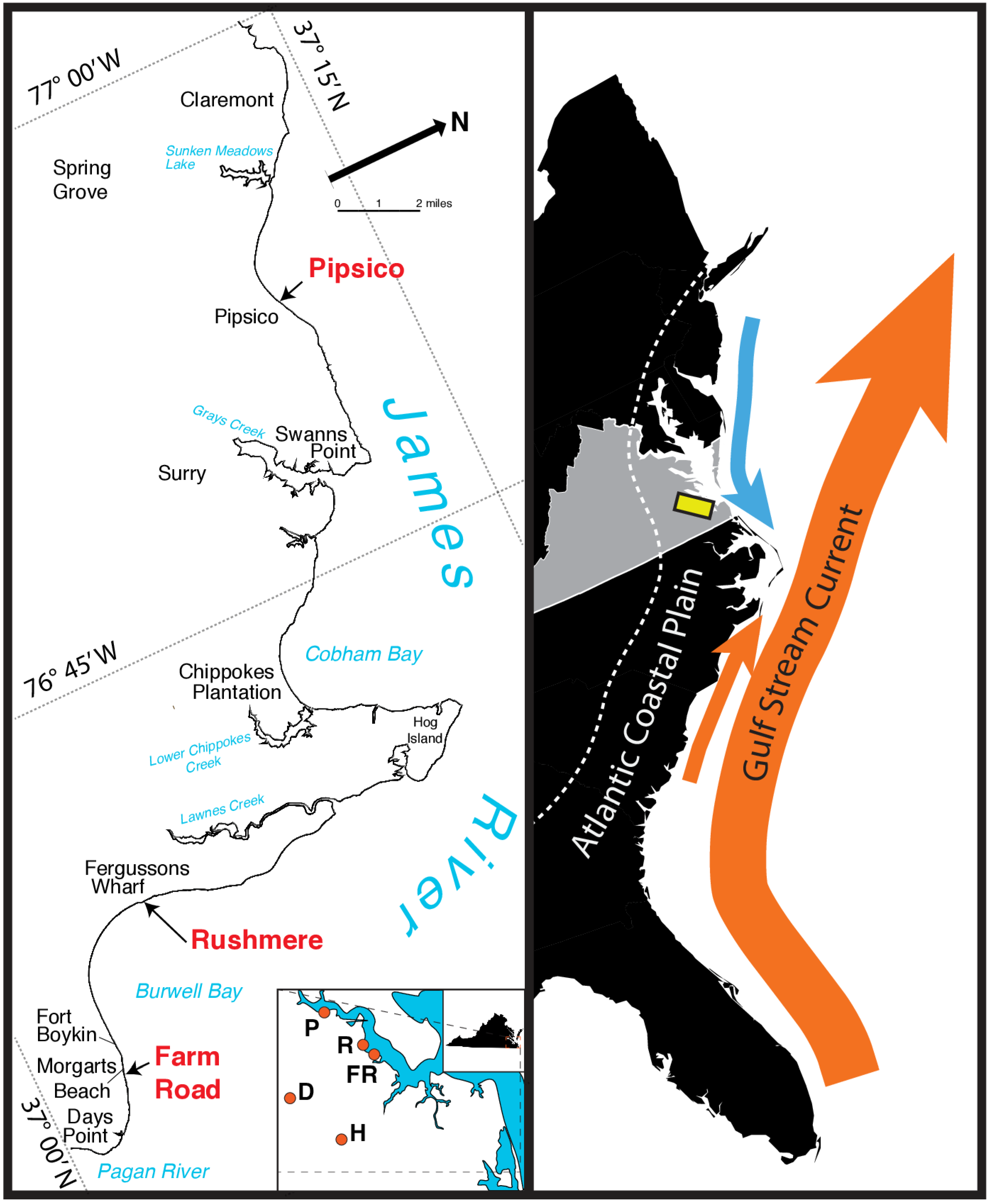
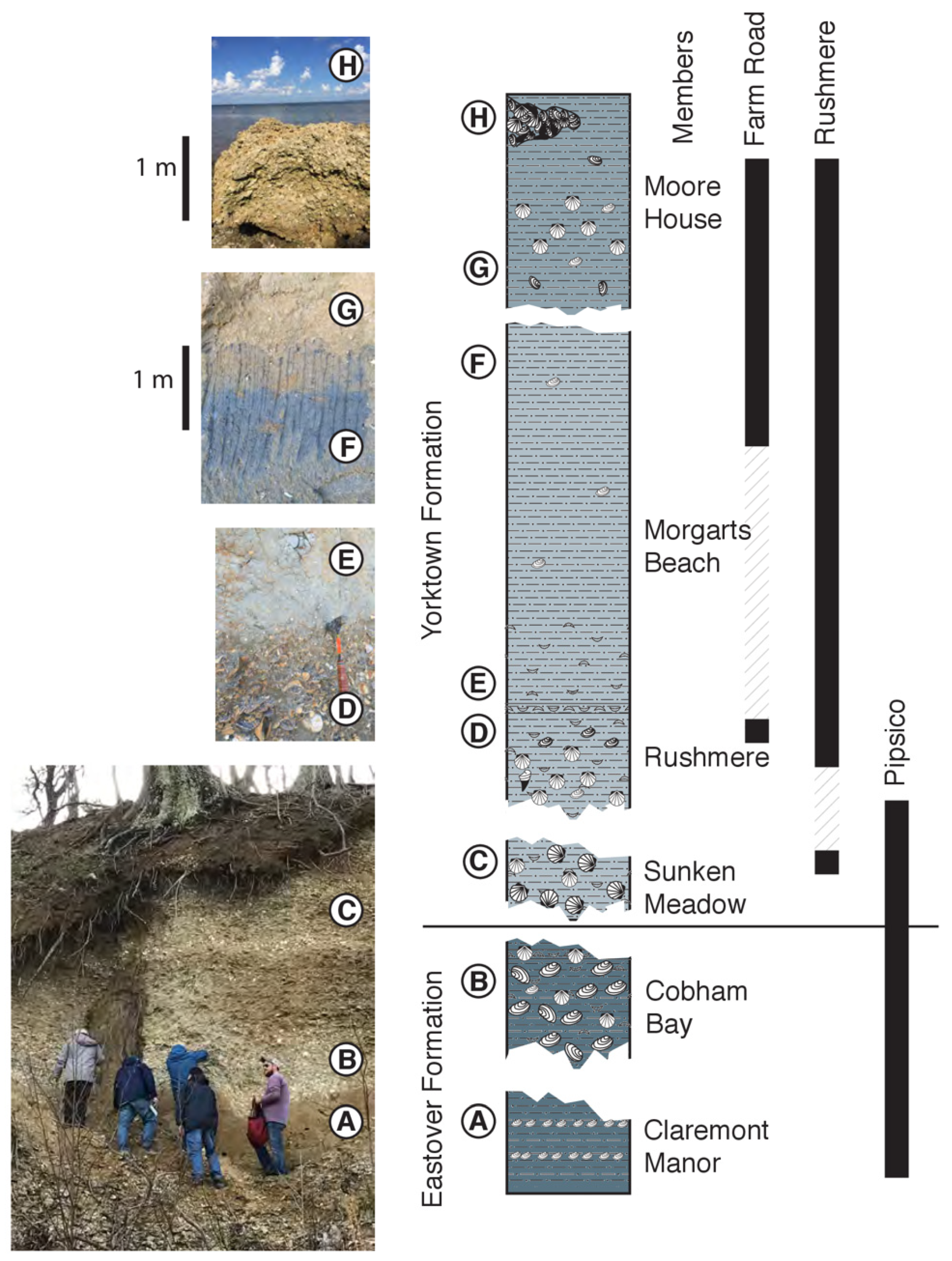
2.3. James River Composite Section
3. Materials and Methods
3.1. Alkenones
3.2. Planktonic Foraminifera
4. Results
4.1. Alkenone SST and Productivity
4.2. Faunal Analysis
5. Discussion
5.1. Chronology
5.2. Mid-Piacenzian Conditions on the MACP
5.3. Paleoclimate Interpretations from Yorktown Formation Data
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, B.; Midgley, B.M. IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Hayhoe, K.; Edmonds, J.; Kopp, R.E.; LeGrande, A.N.; Sanderson, B.M.; Wehner, M.F.; Wuebbles, D.J. Climate models, scenarios, and projections. In Climate Science Special Report: Fourth National Climate Assessment, Volume I; Wuebbles, D.J., Fahey, D.W., Hibbard, K.A., Dokken, D.J., Stewart, B.C., Maycock, T.K., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2017; pp. 133–160. [Google Scholar]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Ginevra, Switzerland, 2021.
- Haywood, A.M.; Dowsett, H.J.; Dolan, A.M. Integrating geological archives and climate models for the mid-Pliocene warm period. Nat. Commun. 2016, 7, 10646. [Google Scholar] [CrossRef] [PubMed]
- Dowsett, H.; Robinson, M.; Haywood, A.; Salzmann, U.; Hill, D.; Sohl, L.; Chandler, M.; Williams, M.; Foley, K.; Stoll, D. The PRISM3D paleoenvironmental reconstruction. Stratigraphy 2010, 7, 123–139. [Google Scholar]
- De la Vega, E.; Chalk, T.B.; Wilson, P.A.; Bysani, R.P.; Foster, G.L. Atmospheric CO2 during the mid-piacenzian warm period and the M2 glaciation. Sci. Rep. 2020, 10, 11002. [Google Scholar] [CrossRef]
- Haywood, A.M.; Tindall, J.C.; Dowsett, H.J.; Dolan, A.M.; Foley, K.M.; Hunter, S.J.; Hill, D.J.; Chan, W.L.; Abe-Ouchi, A.; Stepanek, C.; et al. The Pliocene model intercomparison project phase 2: Large-scale climate features and climate sensitivity. Clim. Past 2020, 16, 2095–2123. [Google Scholar] [CrossRef]
- Hazel, J.E. Paleoclimatology of the Yorktown Formation (Upper Miocene and Lower Pliocene) of Virginia and North Carolinas. Cent. Rech. Pau-SNPA Bull. 1971, 5, 361–375. [Google Scholar]
- Ward, L.W.; Blackwelder, B.W. Stratigraphic Revision of Upper Miocene and LOWER Pliocene Beds of the Chesapeake Group, Middle Atlantic Coastal Plain; U.S. Geological Survey Bull.: Reston, VA, USA, 1980; Volume 1482-D, p. 61.
- Gibson, T.G. Stratigraphy of miocene through Lower Pleistocene strata of the United States central Atlantic coastal plain. In Geology and Paleontology of the Lee Creek Mine, North Carolina; I: Smithsonian Contributions to Paleobiology; Ray, C.E., Ed.; Smithsonian Institution Offices: Washington, DC, USA, 1983; Volume 53, pp. 35–80. [Google Scholar]
- Snyder, S.W.; Mauger, L.L.; Akers, W.H. Planktonic foraminifera and biostratigraphy of the Yorktown Formation, Lee Creek Mine. In Geology and Paleontology of the Lee Creek Mine, North Carolina, I; Ray, C.E., Ed.; Smithsonian: Washington, DC, USA, 1983; Volume 53, pp. 455–481. [Google Scholar]
- Cronin, T.M. Pliocene shallow water paleoceanography of the North Atlantic ocean based on marine ostracodes. Quat. Sci. Rev. 1991, 10, 175–188. [Google Scholar] [CrossRef]
- Krantz, D.E. A chronology of Pliocene sea-level fluctuations: The U.S. Middle Atlantic coastal plain record. Quat. Sci. Rev. 1991, 10, 163–174. [Google Scholar] [CrossRef]
- Dowsett, H.J.; Wiggs, L.B. Planktonic foraminiferal assemblage of the Yorktown Formation, Virginia, USA. Micropaleontology 1992, 38, 75–86. [Google Scholar] [CrossRef]
- Campbell, L.D. Pliocene Molluscs from the Yorktown and Chowan River Formations in Virginia; Commonwealth of Virginia, Department of Mines, Minerals, and Energy, Division of Mineral Resources: Washington, DC, USA, 1993; Volume 127, p. 259.
- Williams, M.; Haywood, A.M.; Harper, E.M.; Johnson, A.L.A.; Knowles, T.; Leng, M.J.; Lunt, D.J.; Okamura, B.; Taylor, P.D.; Zalasiewics, J. Pliocene climate and seasonality in North Atlantic shelf seas. Philos. Trans. R. Soc. A 2009, 367, 85–108. [Google Scholar] [CrossRef]
- Johnson, A.L.A.; Valentine, A.; Leng, M.J.; Sloane, H.J.; SchÖNe, B.R.; Balson, P.S. Isotopic temperatures from the early and mid-Pliocene of the US middle Atlantic coastal plain, and their implications for the cause of regional marine climate change. Palaios 2017, 32, 250–269. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, H.J.; Robinson, M.M.; Foley, K.M.; Herbert, T.D.; Otto-Bliesner, B.L.; Spivey, W. The mid-Piacenzian of the North Atlantic Ocean. Stratigraphy 2019, 16, 119–144. [Google Scholar] [CrossRef]
- Johnson, A.L.A.; Valentine, A.M.; Leng, M.J.; SchÖNe, B.R.; Sloane, H.J. Life history, environment and extinction of the scallop carolinapecten eboreus (conrad) in the plio-pleistocene of the U.S. eastern seaboard. Palaios 2019, 34, 49–70. [Google Scholar] [CrossRef]
- Kier, P.M. Upper Miocene Echinoids from the Yorktown Formation of Virginia and Their Environmental Significance; Smithsonian Institution: Washington, DC, USA, 1972. [Google Scholar]
- Dowsett, H.; Dolan, A.; Rowley, D.; Moucha, R.; Forte, A.M.; Mitrovica, J.X.; Pound, M.; Salzmann, U.; Robinson, M.; Chandler, M.; et al. The PRISM4 (mid-Piacenzian) paleoenvironmental reconstruction. Clim. Past 2016, 12, 1519–1538. [Google Scholar] [CrossRef] [Green Version]
- Colquhoun, D.J. The geology and physiography of the Orangeburg Scarp. In Southeastern Section of the Geological Society of America, Centennial Field Guide; Neathery, T.L., Ed.; Geological Society of America: Boulder, CO, USA, 1986; Volume 6, pp. 321–322. [Google Scholar]
- Dowsett, H.J.; Cronin, T.M. High eustatic sea level during the middle Pliocene: Evidence from the southeastern U.S. Atlantic Coastal Plain. Geology 1990, 18, 435–438. [Google Scholar] [CrossRef]
- Rowley, D.B.; Forte, A.M.; Moucha, R.; Mitrovica, J.X.; Simmons, N.A.; Grand, S.P. Dynamic topography change of the Eastern United States since 3 million years ago. Science 2013, 340, 1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, L.W.; Allmon, W.D. History of paleontology in Virginia: 1607–2007. Bull. Am. Paleontol. 2019, 397, 196. [Google Scholar]
- Lyell, C. Travels in North America: With Geological Observations on the United States, Canada, and Nova Scotia; J. Murray: London, UK, 1845; Volume 2. [Google Scholar]
- Dana, J.D. Manual of Geology, 4th ed.; Ivison, Blakeman, Taylor: New York, NY, USA, 1895. [Google Scholar]
- Dana, J.D. Manual of Geology, 3rd ed.; Ivison, Blakeman, Taylor: New York, NY, USA, 1880; p. 912. [Google Scholar]
- Dana, J.D. Manual of Geology: Treating of the Principles of the Science, with Special Reference to American Geological History, for the Use of Colleges, Academies, and Schools of Science; T. Bliss & Company: Philadelphia, PA, USA, 1862; p. 798. [Google Scholar]
- Dall, W.H.; Harris, G.D. Correlation papers Neocene. U. S. Geol. Surv. Bull. 1892, 84, 349. [Google Scholar]
- Clark, W.B. A brief summary of the geology of the Virginia Coastal Plain. Va. Geol. Surv. Bull. 1906, 2, 11–24. [Google Scholar]
- Clark, W.B.; Miller, B.L.; Berry, E.W.; Watson, T.L. The Physiography and Geology of the Coastal Plain Province of Virginia; University of Virginia: Charlottesville, VA, USA, 1912. [Google Scholar]
- Mansfield, W.C. New fossil mollusks from the Miocene of Virginia and North Carolina, with a brief outline of the divisions of the Chesa-peake group. U. S. Natl. Mus. Proc. 1928, 74, 1–2. [Google Scholar] [CrossRef]
- Mansfield, W.C. Stratigraphy of the Miocene of Virginia and the Miocene and Pliocene of North Carolina: Mollusca from the Miocene and lower Pliocene of Virginia and North Carolina: Part 1. Pelecypoda. With a summary of the stratigraphy. U. S. Geol. Surv. Prof. Pap. 1943, 199-A, 1–178. [Google Scholar]
- Hazel, J.E. Atlantic continental shelf and slope of the United States—Ostracode zoogeography in the southern Nova Scotian and northern Virginian faunal provinces. U. S. Geol. Surv. Prof. Pap. 1970, 529E, 1–21. [Google Scholar]
- Hazel, J.E. Ostracode biostratigraphy of the Yorktown Formation (upper Miocene and lower Pliocene) of Virginia and North Carolina. U. S. Geol. Surv. Prof. Pap. 1971, 704, 13. [Google Scholar]
- Ward, L.W. Geology and Paleontology of the James River: Richmond to Hampton Roads; Virginia Museum of Natural History: Martinsville, VA, USA, 2008. [Google Scholar]
- Cronin, T.M.; Dowsett, H.J.; Gosnell, L.B.; Ross, R.M. Pliocene Marine Micropaleontology of Southeastern Virginia and Northeastern North Carolina; US Geological Survey Open-File Rep.; US Geological Survey: Reston, VA, USA, 1989. [CrossRef] [Green Version]
- Blow, W.H. Late middle Eocene to recent planktonic foraminiferal biostratigraphy. In Proceedings of the 1st International Conference on Planktonic Microfossils, Geneva, Switzerland, 1 January 1969; Bronniman, P., Renz, H.H., Eds.; E. J. Brill: Leiden, The Netherlands, 1969; pp. 199–422. [Google Scholar]
- Berggren, W.A. The pliocene time scale: Calibration of planktonic foraminiferal and calcareous nannoplankton zones. Nature 1973, 243, 391–397. [Google Scholar] [CrossRef]
- Ogg, J.G. Chapter 5—Geomagnetic polarity time scale. In The Geologic Time Scale; Elsevier: Boston, MA, USA, 2012; pp. 85–113. [Google Scholar]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography 2005, 20. [Google Scholar] [CrossRef] [Green Version]
- De Schepper, S.; Head, M.J.; Groeneveld, J. North Atlantic Current variability through marine isotope stage M2 (circa 3.3 Ma) during the mid-Pliocene. Paleoceanography 2009, 24, PA4206. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, H.J.; Foley, K.M.; Robinson, M.M.; Herbert, T.D. PRISM Late Pliocene (Piacenzian) Alkenone—Derived SST Data; U.S. Geological Survey: Reston, VA, USA, 2017. [CrossRef]
- Volkman, J.K. Ecological and environmental factors affecting alkenone distributions in seawater and sediments. Geochem. Geophys. Geosyst. 2000, 1, 1036. [Google Scholar] [CrossRef]
- Herbert, T.D. Review of alkenone calibrations (culture, water column, and sediments). Geochem. Geophys. Geosyst. 2001, 2, 1005. [Google Scholar] [CrossRef]
- Herbert, T.D. Alkenone palaeotemperature determinations. In The Oceans and Marine Geochemistry; Elderfield, H., Holland, H.D., Turekian, K.K., Eds.; Treatise on geochemistry Elsevier-Pergamon: Oxford, UK, 2004; Volume 6, pp. 391–432. [Google Scholar]
- Lawrence, K.T.; Herbert, T.D.; Dekens, P.S.; Ravelo, A.C. The application of the alkenone organic proxy to the study of Plio-Pleistocene climate. In Deep-Time Perspectives on Climate Change: Marrying the Signal from Computer Models and Biological Proxies; Williams, M., Haywood, A.M., Gregory, F.J., Schmidt, D.N., Eds.; Micropalaeontological Society (Special Publication), Geological Society of London: London, UK, 2007; pp. 539–562. [Google Scholar]
- Müller, P.J.; Kirst, G.; Ruhland, G.; von Storch, I.; Rosell-MelÈ, A. Calibration of the alkenone paleotemperature index U37K’ based on core-tops from the eastern South Atlantic and the global ocean (60° N–60° S). Geochim. Cosmochim. Acta 1998, 62, 1757–1772. [Google Scholar] [CrossRef]
- Bolton, C.T.; Lawrence, K.T.; Gibbs, S.J.; Wilson, P.A.; Cleaveland, L.C.; Herbert, T.D. Glacial-interglacial productivity changes recorded by alkenones and microfossils in late Pliocene eastern equatorial Pacific and Atlantic upwelling zones. Earth Planet. Sci. Lett. 2010, 295, 401–411. [Google Scholar] [CrossRef]
- Bolton, C.T.; Lawrence, K.T.; Gibbs, S.J.; Wilson, P.A.; Herbert, T.D. Biotic and geochemical evidence for a global latitudinal shift in ocean biogeochemistry and export productivity during the late Pliocene. Earth Planet. Sci. Lett. 2011, 308, 200–210. [Google Scholar] [CrossRef]
- Howe, H.V. Use of soap in the preparation of samples for micropaleontologic study. J. Paleontol. 1941, 15, 691. [Google Scholar]
- Parker, F.L. Planktonic foraminiferal species in Pacific sediments. Micropaleontology 1962, 8, 219–254. [Google Scholar] [CrossRef]
- Parker, F.L. Late Tertiary biostratigraphy (planktonic foraminifera) of tropical Indo-Pacific deep-sea cores. Bull. Am. Paleontol. 1967, 52, 115–208. [Google Scholar]
- Dowsett, H.J.; Robinson, M.M. Mid-Pliocene planktic foraminifer assemblage of the North Atlantic Ocean. Micropaleontology 2007, 53, 105–126. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Buzas, M.A.; Gibson, T.G. Species diversity: Benthonic foraminifera in western North Atlantic. Science 1969, 163, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Collwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples, Version 9. User’s Guide and Application. Available online: http://purl.oclc.org/estimates (accessed on 1 January 2019).
- Dowsett, H.J.; Robinson, M.M.; Foley, K.M. Estimating Piacenzian Sea Surface Temperature Using an Alkenone-Calibrated Transfer Function; U.S. Geological Survey Science Investigate Report; U.S. Geological Survey: Reston, VA, USA, 2021; Volume 17. [CrossRef]
- Dowsett, H.J.; Spivey, W. Planktonic Foraminifer Census Data from Type Section of Yorktown Formation at Rushmere, Virginia; U. S. Geological Survey Data Release: Reston, VA, USA, 2021. [CrossRef]
- Cronin, T.M.; Bybell, L.M.; Poore, R.Z.; Blackwelder, B.W.; Liddicoat, J.C.; Hazel, J.E. Age and correlation of emerged pliocene and pleistocene deposits, U.S. Atlantic Coastal Plain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1984, 47, 21–51. [Google Scholar] [CrossRef]
- Shackleton, N.J.; Hall, M.A. Oxygen and carbon isotope stratigraphy of Deep Sea Drilling Project Hole 552A: Plio-Pleistocene glacial history. Initial. Rep. Deep. Sea Drill. Proj. 1985, 81, 599–609. [Google Scholar] [CrossRef]
- Prell, W.L. Pliocene stable isotope and carbonate stratigraphy (Holes 572C and 573A): Paleoceanographic data bearing on the question of Pliocene glaciation. Initial. Rep. Deep. Sea Drill. Proj. 1985, 85, 723–734. [Google Scholar] [CrossRef]
- Hodell, D.A.; Kennett, J.P. Late Miocene—Early pliocene stratigraphy and paleoceanography of the South Atlantic and Southwest Pacific Oceans: A synthesis. Paleoceanography 1986, 1, 285–311. [Google Scholar] [CrossRef]
- Keigwin, L.D. Pliocene stable-isotope record of Deep Sea Drilling Project Site 606: Sequential events of 18O enrichment beginning at 3.1 Ma. Initial. Rep. Deep. Sea Drill. Proj. 1987, 94, 911–917. [Google Scholar] [CrossRef]
- Raymo, M.E.; Ruddiman, W.F.; Backman, J.; Clement, B.M.; Martinson, D.G. Late pliocene variation in Northern Hemisphere Ice Sheets and North Atlantic deep water circulation. Paleoceanography 1989, 4, 413–446. [Google Scholar] [CrossRef]
- Curry, W.B.; Miller, K.G. Oxygen and carbon isotopic variation in Pliocene benthic foraminifers of the equatorial Atlantic. Proc. Ocean. Drill. Program Sci. Results 1989, 108, 157–166. [Google Scholar]
- Lisiecki, L.E.; Raymo, M.E. Pliocene-Pleistocene climate evolution: Trends and transitions in glacial cycle dynamics. Quat. Sci. Rev. 2007, 26. [Google Scholar] [CrossRef] [Green Version]
- Haywood, A.M.; Dowsett, H.J.; Dolan, A.M.; Rowley, D.; Abe-Ouchi, A.; Otto-Bliesner, B.; Chandler, M.A.; Hunter, S.J.; Lunt, D.J.; Pound, M.; et al. The Pliocene Model Intercomparison Project (PlioMIP) Phase 2: Scientific objectives and experimental design. Clim. Past 2016, 12, 663–675. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, H.J.; Foley, K.M.; Stoll, D.K.; Chandler, M.A.; Sohl, L.E.; Bentsen, M.; Otto-Bliesner, B.L.; Bragg, F.J.; Chan, W.-L.; Contoux, C.; et al. Sea surface temperature of the mid-Piacenzian Ocean: A data-model comparison. Sci. Rep. 2013, 3, 1–8. [Google Scholar] [CrossRef]
- Salzmann, U.; Dolan, A.M.; Haywood, A.M.; Chan, W.-L.; Voss, J.; Hill, D.J.; Abe-Ouchi, A.; Otto-Bliesner, B.; Bragg, F.J.; Chandler, M.A.; et al. Challenges in quantifying Pliocene terrestrial warming revealed by data-model discord. Nat. Clim. Chang. 2013, 3, 969–974. [Google Scholar] [CrossRef]
- Foley, K.M.; Dowsett, H.J. Community Sourced Mid-Piacenzian Sea Surface Temperature (SST) Data; U.S. Geological Survey Data Release: Reston, VA, USA, 2019. [CrossRef]
- Chandan, D.; Peltier, W.R. Regional and global climate for the mid-Pliocene using the University of Toronto version of CCSM4 and PlioMIP2 boundary conditions. Clim. Past 2017, 13, 919–942. [Google Scholar] [CrossRef] [Green Version]
- Chandan, D.; Peltier, W.R. On the mechanisms of warming the mid-Pliocene and the inference of a hierarchy of climate sensitivities with relevance to the understanding of climate futures. Clim. Past 2018, 14, 825–856. [Google Scholar] [CrossRef] [Green Version]
- Hunter, S.J.; Haywood, A.M.; Dolan, A.M.; Tindall, J.C. The HadCM3 contribution to PlioMIP Phase 2 Part 1: Core and Tier 1 experiments. Clim. Past Discuss. 2019, 2019, 1–38. [Google Scholar] [CrossRef]
- Kamae, Y.; Yoshida, K.; Ueda, H. Sensitivity of Pliocene climate simulations in MRI-CGCM2.3 to respective boundary conditions. Clim. Past Discuss. 2016, 2016, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, X.; Guo, C.; Otterå, O.H.; Nisancioglu, K.H.; Tan, N.; Contoux, C.; Ramstein, G.; Feng, R.; Otto-Bliesner, B.L.; et al. Mid-Pliocene Atlantic meridional overturning circulation simulated in PlioMIP2. Clim. Past 2021, 17, 529–543. [Google Scholar] [CrossRef]
- Tan, N.; Contoux, C.; Ramstein, G.; Sun, Y.; Dumas, C.; Sepulchre, P.; Guo, Z. Modeling a modern-like pCO2 warm period (Marine Isotope Stage KM5c) with two versions of an Institut Pierre Simon Laplace atmosphere–ocean coupled general circulation model. Clim. Past 2020, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Stepanek, C.; Samakinwa, E.; Knorr, G.; Lohmann, G. Contribution of the coupled atmosphere–ocean–sea ice–vegetation model COSMOS to the PlioMIP2. Clim. Past 2020, 16, 2275–2323. [Google Scholar] [CrossRef]
- Samakinwa, E.; Stepanek, C.; Lohmann, G. Sensitivity of mid-Pliocene climate to changes in orbital forcing and PlioMIP’s boundary conditions. Clim. Past 2020, 16, 1643–1665. [Google Scholar] [CrossRef]
- Oldeman, A.M.; Baatsen, M.L.J.; von der Heydt, A.S.; Dijkstra, H.A.; Tindall, J.C.; Abe-Ouchi, A.; Booth, A.R.; Brady, E.C.; Chan, W.L.; Chandan, D.; et al. Reduced El Niño variability in the mid-Pliocene according to the PlioMIP2 ensemble. Clim. Past Discuss. 2021, 2021, 1–35. [Google Scholar] [CrossRef]
- Li, X.; Guo, C.; Zhang, Z.; Otterå, O.H.; Zhang, R. PlioMIP2 simulations with NorESM-L and NorESM1-F. Clim. Past 2020, 16, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Zhang, Q.; Li, Q.; Feng, R.; Haywood, A.M.; Tindall, J.C.; Hunter, S.J.; Otto-Bliesner, B.L.; Brady, E.C.; Rosenbloom, N.; et al. Evaluating the large-scale hydrological cycle response within the PlioMIP2 ensemble. Clim. Past Discuss. 2021, 2021, 1–32. [Google Scholar] [CrossRef]
- de Nooijer, W.; Zhang, Q.; Li, Q.; Zhang, Q.; Li, X.; Zhang, Z.; Guo, C.; Nisancioglu, K.H.; Haywood, A.M.; Tindall, J.C.; et al. Evaluation of Arctic warming in mid-Pliocene climate simulations. Clim. Past 2020, 16, 2325–2341. [Google Scholar] [CrossRef]
- Chan, W.L.; Abe-Ouchi, A. Pliocene Model Intercomparison Project (PlioMIP2) simulations using the Model for Interdisciplinary Research on Climate (MIROC4m). Clim. Past 2020, 16, 1523–1545. [Google Scholar] [CrossRef]
- Berntell, E.; Zhang, Q.; Li, Q.; Haywood, A.M.; Tindall, J.C.; Hunter, S.J.; Zhang, Z.; Li, X.; Guo, C.; Nisancioglu, K.H.; et al. Mid-Pliocene West African Monsoon rainfall as simulated in the PlioMIP2 ensemble. Clim. Past 2021, 17, 1777–1794. [Google Scholar] [CrossRef]
- Hunter, S.J.; Haywood, A.M.; Dolan, A.M.; Tindall, J.C. The HadCM3 contribution to PlioMIP phase 2. Clim. Past 2019, 15, 1691–1713. [Google Scholar] [CrossRef] [Green Version]
- Krantz, D.E. Mollusk-isotope records of plio-pleistocene marine paleoclimate, U.S. Middle Atlantic Coastal Plain. Palaios 1990, 5, 317–335. [Google Scholar] [CrossRef]
- Winkelstern, I.A.N.; Surge, D.; Hudley, J.W. Mulltiproxy sclerochronological evidence for Plio-Pleistocene regional warmth: United States Mid-Atlantic coastal plain. Palaios 2013, 28, 649–660. [Google Scholar] [CrossRef]
- Seidov, D.; Baranova, O.K.; Johnson, D.R.; Boyer, T.P.; Mishonov, A.V.; Parsons, A.R. Northwest Atlantic Regional Climatology; NOAA Atlas NESDIS 80, Tech.; NOAA: Silver Spring, MD, USA, 2016. [Google Scholar] [CrossRef]
- Huang, B.; Thorne, P.W.; Banzan, V.F.; Boyer, T.; Chepurin, G.; Lawrimore, J.H.; Menne, M.J.; Smith, T.M.; Vose, R.S.; Zhang, H.-M. NOAA Extended Reconstructed Sea Surface Temperature (ERSST), Version 5; NOAA National Centers for Environmental Information: Washington, DC, USA, 2017. [Google Scholar] [CrossRef]
- Bova, S.; Rosenthal, Y.; Liu, Z.; Godad, S.P.; Yan, M. Seasonal origin of the thermal maxima at the Holocene and the last interglacial. Nature 2021, 589, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Dowsett, H.J.; Robinson, M.M.; Stoll, D.K.; Foley, K.M.; Johnson, A.L.A.; Williams, M.; Riesselman, C.R. The PRISM (Pliocene Palaeoclimate) reconstruction: Time for a paradigm shift. Philos. Trans. R. Soc. 2013, 371, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Lyell, C. On the Miocene Tertiary Strata of Maryland, Virginia, and of North and South Carolina. Q. J. Geol. Soc. 1845, 1, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Cronin, T.M.; Dowsett, H.J. A quantitative micropaleontologic method for shallow marine peleoclimatology: Application to Pliocene deposits of the western North Atlantic Ocean. Mar. Micropaleontol. 1990, 16, 117–147. [Google Scholar] [CrossRef]
- Cronin, T.M. Evolution of marine climates of the U.S. Atlantic Coast during the past four million years. Philos. Trans. R. Society. Ser. B Biol. Sci. 1988, 318, 661–678. [Google Scholar]
- Ward, L.W.; Bailey, R.H.; Carter, J.G. Pliocene and early Pleistocene stratigraphy, depositional history, and molluscan paleobiogeography of the coastal plain. In The geology of the Carolinas: Carolina Geological Society Fiftieth Anniversary Volume; Horton, J.W., Zullo, V.A., Eds.; University of Tennessee Press: Knoxville, TN, USA, 1991; pp. 274–289. [Google Scholar]
- Johnson, A.L.A.; Hickson, J.A.; Bird, A.; Schône, B.R.; Balson, P.S.; Heaton, T.H.E.; Williams, M. Comparative sclerochronology of modern and mid-Pliocene (c. 3.5 Ma) Aequipecten opercularis (Mollusca, Bivalvia): An insight into past and future climate change in the north-east Atlantic region. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 284, 164–179. [Google Scholar] [CrossRef]
- Valentine, A.; Johnson, A.L.; Leng, M.J.; Sloane, H.J.; Balson, P.S. Isotopic evidence of cool winter conditions in the mid-Piacenzian (Pliocene) of the southern North Sea Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 309, 9–16. [Google Scholar] [CrossRef]
- Cronin, T.M.; Dowsett, H.J. Biotic and oceanographic response to the Pliocene closing of the Central American isthmus. In Evolution and Environment in Tropical America; Jackson, J.B.C., Budd, A.F., Coates, A.G., Eds.; University of Chicago Press: Chicago, IL, USA, 1996; pp. 76–104. [Google Scholar]
- Dowsett, H.J.; Cronin, T.M.; Poore, R.Z.; Thompson, R.S.; Whatley, R.C.; Wood, A.M. Micropaleontological Evidence for Increased Meridional Heat Transport in the North Atlantic Ocean During the Pliocene. Science 1992, 258, 1133–1135. [Google Scholar] [CrossRef] [PubMed]
- Haug, G.H.; Tiedemann, R. Effect of the formation of the Isthmus of Panama on Atlantic Ocean thermohaline circulation. Nature 1998, 393, 673–676. [Google Scholar] [CrossRef]
- Haug, G.; Tiedemann, R.; Zahn, R.; Ravelo, A. Role of Panama uplift on oceanic freshwater balance. Geology 2001, 29, 207–210. [Google Scholar] [CrossRef]
- Montes, C.; Cardona, A.; Jaramillo, C.; Pardo, A.; Silva, J.C.; Valencia, V.; Ayala, C.; Pérez-Angel, L.C.; Rodriguez-Parra, L.A.; Ramirez, V. Middle miocene closure of the Central American seaway. Science 2015, 348, 226–229. [Google Scholar] [CrossRef] [Green Version]
- Coates, A.; Jackson, J.B.C.; Collins, L.S.; Cronin, T.M.; Dowsett, H.J.; Bybell, L.M.; Jung, P.; Obando, J.A. Closure of the Isthmus of Panama: The near-shore marine record of Costa Rica and western Panama. Geol. Soc. Am. Bull. 1992, 104, 814–828. [Google Scholar] [CrossRef]
- Ward, L.W.; Gilinsky, N.L. Molluscan Assemblages of the Chowan River Formation; Virginia Museum of Natural History: Martinsville, VA, USA, 1993. [Google Scholar]
- Knowles, T.; Taylor, P.D.; Williams, M.; Haywood, A.M.; Okamura, B. Pliocene seasonality across the North Atlantic inferred from cheilostome bryozoans. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 277, 226–235. [Google Scholar] [CrossRef]
- Groeneveld, J. Effect of the Pliocene Closure of the Panamanian Gateway on Caribbean and East Pacific Sea Surface Temperatures and Salinities by Applying Combined Mg/Ca and δ18O Measurements (5.6–2.2 Ma); University of Kiel: Kiel, Germany, 2005. [Google Scholar]
- Bartoli, G.; Sarnthein, M.; Weinelt, M.; Erlenkeuser, H.; Garbe-Schonberg, D.; Lea, D.W. Final closure of Panama and the onset of northern hemisphere glaciation. Earth Planet. Sci. Lett. 2005, 237, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.M.; Dowsett, H.J.; Dwyer, G.S.; Lawrence, K.T. Reevaluation of mid-Pliocene North Atlantic sea surface temperatures. Paleoceanography 2008, 23. [Google Scholar] [CrossRef] [Green Version]
- Dowsett, H.J.; Chandler, M.A.; Robinson, M.M. Surface temperatures of the Mid-Pliocene North Atlantic Ocean: Implications for future climate. Philos. Trans. R. Soc. A 2009, 367, 69–84. [Google Scholar] [CrossRef]
- Sarnthein, M.; Bartoli, G.; Prange, M.; Schmittner, A.; Schneider, B.; Weinelt, M.; Andersen, N.; Garbe-Sch√∂nberg, D. Mid-Pliocene shifts in ocean overturning circulation and the onset of Quaternary-style climates. Clim. Past 2009, 5, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Herbert, T.D.; Peterson, L.C.; Lawrence, K.T.; Liu, Z. Tropical Ocean Temperatures Over the Past 3.5 Million Years. Science 2010, 328, 1530–1534. [Google Scholar] [CrossRef]
- Lawrence, K.T.; Herbert, T.D.; Brown, C.M.; Raymo, M.E.; Haywood, A.M. High-amplitude variations in North Atlantic sea surface temperature during the early Pliocene warm period. Paleoceanography 2009, 24, PA2218. [Google Scholar] [CrossRef]
- Robinson, M.M. New quantitative evidence of extreme warmth in the Pliocene Arctic. Stratigraphy 2009, 6, 265–275. [Google Scholar]
- De Schepper, S.; Groeneveld, J.; Naafs, B.D.A.; Van Renterghem, C.; Hennissen, J.; Head, M.J.; Louwye, S.; Fabian, K. Northern Hemisphere glaciation during the globally warm early Late Pliocene. PLoS ONE 2013, 8, e81508. [Google Scholar] [CrossRef]
- Lunt, D.; Valdes, P.; Haywood, A.; Rutt, I. Closure of the Panama Seaway during the Pliocene: Implications for climate and Northern Hemisphere glaciation. Clim. Dyn. 2008, 30, 1–18. [Google Scholar] [CrossRef]
- Marincovich, L.; Gladenkov, A.Y. Evidence for an early opening of the Bering Strait. Nature 1999, 397, 149–151. [Google Scholar] [CrossRef]
- Marincovich, L.; Gladenkov, A.Y. New evidence for the age of Bering Strait. Quat. Sci. Rev. 2001, 20, 329–335. [Google Scholar] [CrossRef]
- Cronin, T.M.; Whatley, R.; Wood, A.; Tsukagoshi, A.; Ikeya, N.; Brouwers, E.M.; Briggs, W.M., Jr. Microfaunal Evidence for Elevated Pliocene Temperatures in the Arctic Ocean. Paleoceanography 1993, 8, 161–173. [Google Scholar] [CrossRef]
- Matthiessen, J.; Knies, J.; Vogt, C.; Stein, R. Pliocene palaeoceanography of the Arctic Ocean and subarctic seas. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2009, 367, 21–48. [Google Scholar] [CrossRef]
- Hopkins, D.M. Cenozoic History of the Bering Land Bridge. Science 1959, 129, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.M. The Cenozoic History of Beringia—A Synthesis; Stanford University Press: Stanford, CA, USA, 1967; pp. 451–484. [Google Scholar]
- Nelson, C.H.; Hopkins, D.; Scholl, D. Tectonic Setting and Cenozoic Sedimentary History of the Bering Sea. In Marine Geology and Oceanography of the Arctic Seas; Herman, Y., Ed.; Springer: Berlin/Heidelberg, Germany, 1974; pp. 119–140. [Google Scholar]
- Hu, A.; Meehl, G.A.; Han, W.; Otto-Bliestner, B.; Abe-Ouchi, A.; Rosenbloom, N. Effects of the Bering Strait closure on AMOC and global climate under different background climates. Prog. Oceanogr. 2015, 132, 174–196. [Google Scholar] [CrossRef] [Green Version]
- Feng, R.; Otto-Bliesner, B.L.; Fletcher, T.; Tabor, C.; Ballantyne, A.P.; Brady, E. Amplified Late Pliocene Terrestrial Warmth in Northern High Latitudes from Greater Radiative Forcing and Closed Arctic Ocean Gateways; Elsevier: Amsterdam, The Netherlands, 2017; Volume 466, pp. 129–138. [Google Scholar]
- Otto-Bliesner, B.; Jahn, A.; Feng, R.; Brady, E.; Hu, A.; Löfverström, M. Changes in arctic gateways amplify North Atlantic Warming in the Late Pliocene: Arctic gateways and pliocene climate. Geophys. Res. Lett. 2017, 44, 957–964. [Google Scholar] [CrossRef]
- Gonçalves Neto, A.; Langan, J.A.; Palter, J.B. Changes in the Gulf Stream preceded rapid warming of the Northwest Atlantic Shelf. Commun. Earth Environ. 2021, 2, 74. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Designation | Latitude | Longitude | Members Present |
|---|---|---|---|---|
| Farm Road | – | 37.033 | 76.604 | Rushmere 1, Morgarts Beach, Moore House |
| Rushmere | – | 37.067 | 76.669 | Sunken Meadow 1, Rushmere, Morgarts Beach, Moore House |
| Pipsico | – | 37.202 | 76.882 | Claremont Manor, Cobham Bay, Sunken Meadow, Rushmere |
| Holland core | DEQ 161-592 | 36.682 | 76.781 | Eastover Fm. 2, Rushmere, Morgarts Beach |
| Dory core | DEQ 187-262 | 36.856 | 77.038 | Claremont Manor, Cobham Bay, Rushmere, Morgarts Beach |
| Sample | Member | Elevation (cm) | Dentoglobigerina altispira | Globigerina bulloides | Globigerina falconensis | Globigerina umbilicata | Globigerinella calida | Globigerinella siphonifera | Globigerinita glutinata | Globigerinoides conglobatus | Globigerinoides obliquus | Globigerinoides ruber | Globoconella puncticulata | Globorotalia crassaformis | Globorotalia hirsuta | Globorotalia scitula | Globorotalia tumida | Globoturborotalita apertura | Globoturborotalita woodi | Neogloboquadrina acostaensis | Neogloboquadrina atlantica | Neogloboquadrina humerosa | Neogloboquadrina incompta | Neogloboquadrina pachyderma | Orbulina universa | Sphaeroidinellopsis | Trilobatus sacculifer | Total | Planktics/gm | S | H’ | E |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 12MR10 | Morgarts Beach | 510 | 4 | 3 | 7 | 0.03 | 2 | 0.68 | 0.99 | |||||||||||||||||||||||
| 17HD158 | 387 | 1 | 60 | 3 | 2 | 17 | 28 | 28 | 1 | 7 | 10 | 1 | 2 | 1 | 20 | 181 | 0.72 | 14 | 1.98 | 0.52 | ||||||||||||
| 17HD153 | 299 | 95 | 1 | 1 | 4 | 23 | 43 | 2 | 1 | 3 | 8 | 13 | 3 | 3 | 2 | 202 | 0.81 | 14 | 1.67 | 0.38 | ||||||||||||
| 17HD151 | 259 | 85 | 3 | 3 | 1 | 1 | 29 | 1 | 15 | 2 | 1 | 1 | 4 | 18 | 164 | 0.66 | 13 | 1.55 | 0.36 | |||||||||||||
| 17HD147 | 187 | 3 | 171 | 1 | 2 | 2 | 59 | 3 | 42 | 10 | 6 | 2 | 27 | 3 | 3 | 3 | 36 | 373 | 1.49 | 16 | 1.77 | 0.37 | ||||||||||
| 12HD11 | 100 | 1 | 74 | 2 | 71 | 27 | 51 | 4 | 1 | 5 | 1 | 4 | 1 | 3 | 13 | 1 | 22 | 281 | 1.12 | 16 | 1.95 | 0.44 | ||||||||||
| 17HD141 | Rushmere | 80 | 107 | 2 | 1 | 5 | 23 | 2 | 26 | 5 | 2 | 1 | 2 | 6 | 8 | 3 | 3 | 2 | 45 | 243 | 0.97 | 17 | 1.85 | 0.37 | ||||||||
| 12HD10 | 65 | 36 | 3 | 5 | 28 | 10 | 11 | 2 | 2 | 3 | 3 | 1 | 1 | 43 | 148 | 0.59 | 13 | 1.93 | 0.53 | |||||||||||||
| 17HD140 | 62 | 63 | 1 | 1 | 16 | 8 | 4 | 1 | 2 | 2 | 2 | 100 | 0.40 | 10 | 1.29 | 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dowsett, H.J.; Robinson, M.M.; Foley, K.M.; Herbert, T.D. The Yorktown Formation: Improved Stratigraphy, Chronology, and Paleoclimate Interpretations from the U.S. Mid-Atlantic Coastal Plain. Geosciences 2021, 11, 486. https://doi.org/10.3390/geosciences11120486
Dowsett HJ, Robinson MM, Foley KM, Herbert TD. The Yorktown Formation: Improved Stratigraphy, Chronology, and Paleoclimate Interpretations from the U.S. Mid-Atlantic Coastal Plain. Geosciences. 2021; 11(12):486. https://doi.org/10.3390/geosciences11120486
Chicago/Turabian StyleDowsett, Harry J., Marci M. Robinson, Kevin M. Foley, and Timothy D. Herbert. 2021. "The Yorktown Formation: Improved Stratigraphy, Chronology, and Paleoclimate Interpretations from the U.S. Mid-Atlantic Coastal Plain" Geosciences 11, no. 12: 486. https://doi.org/10.3390/geosciences11120486
APA StyleDowsett, H. J., Robinson, M. M., Foley, K. M., & Herbert, T. D. (2021). The Yorktown Formation: Improved Stratigraphy, Chronology, and Paleoclimate Interpretations from the U.S. Mid-Atlantic Coastal Plain. Geosciences, 11(12), 486. https://doi.org/10.3390/geosciences11120486