
Globorotalia truncatulinoides in the Mediterranean Basin during the Middle–Late Holocene: Bio-Chronological and Oceanographic Indicator

 , , , and
, , , and
Abstract
:1. Introduction
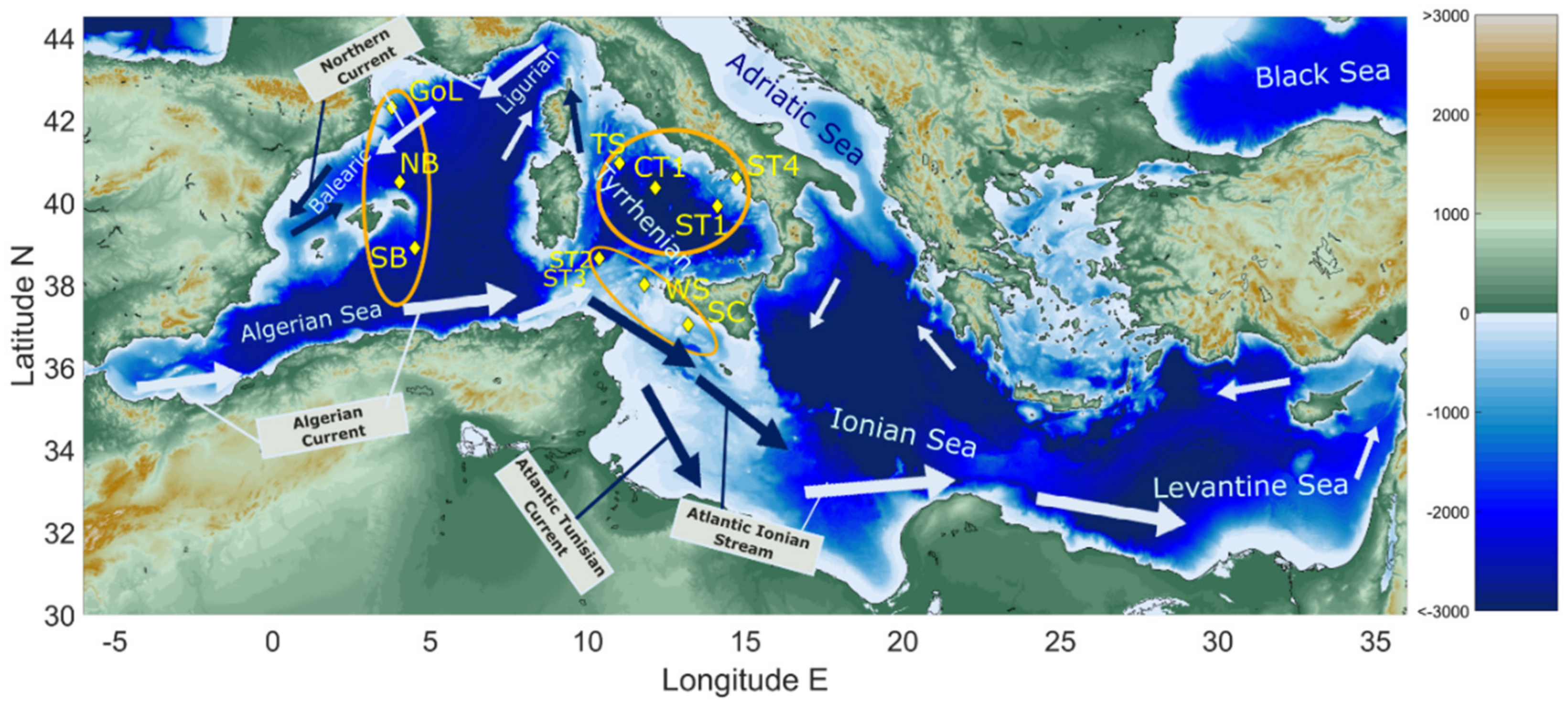
2. Oceanography of the Study Area: Present-Day Conditions
3. Material and Methods
4. G. truncatulinoides: Habitat and Ecology
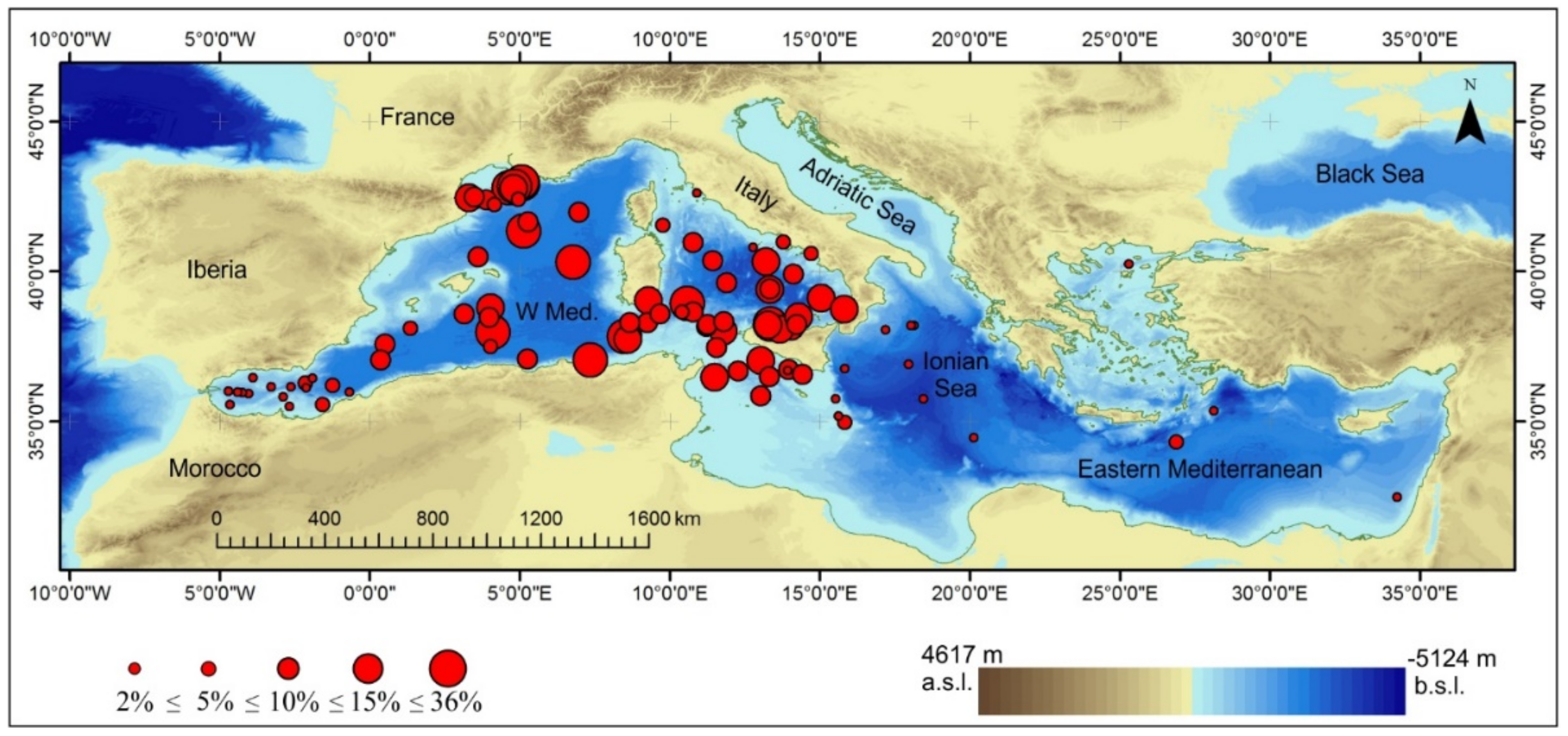
5. Chronological Distribution of Globorotalia truncatulinoides
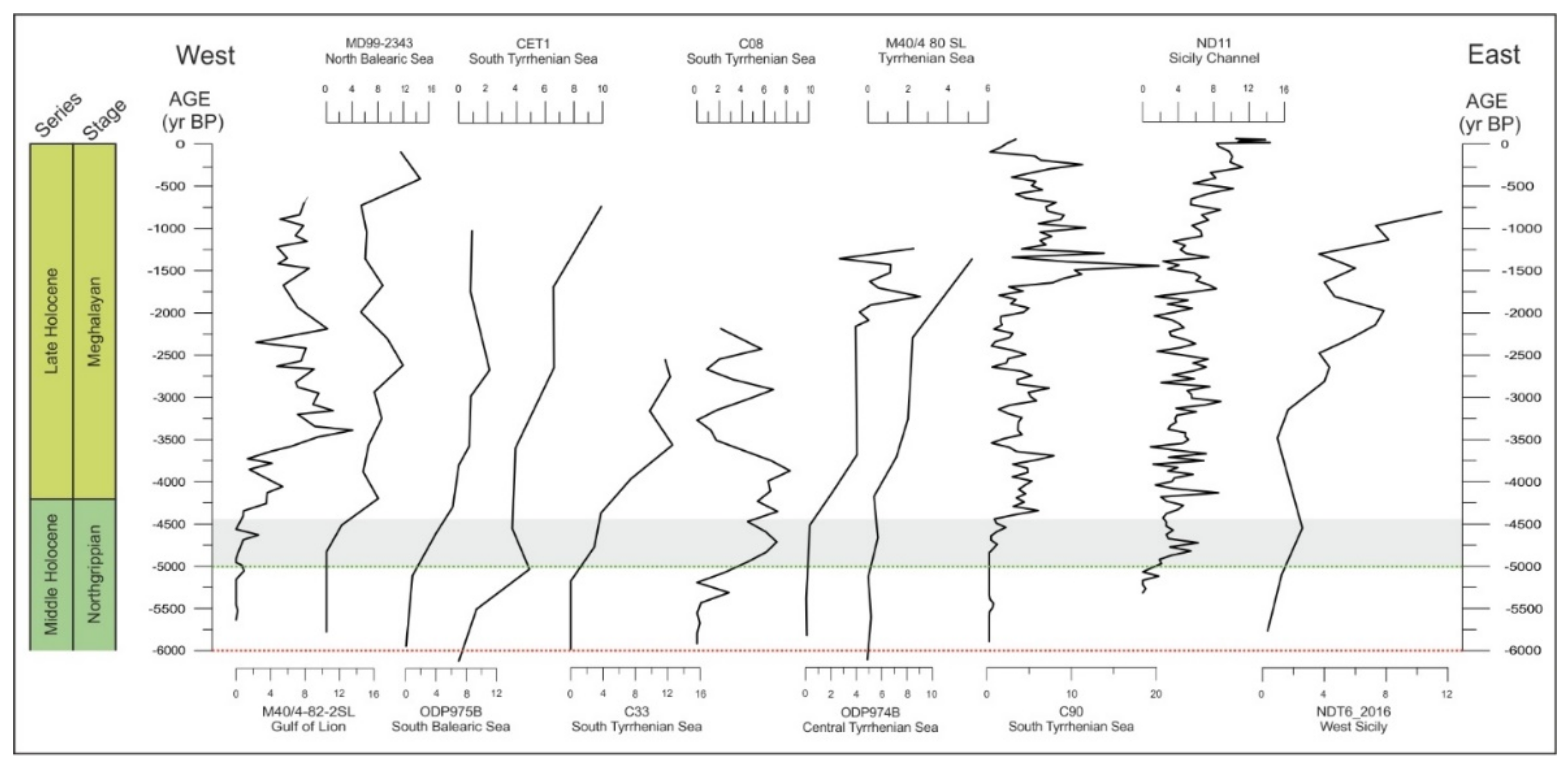
6. Results and Discussion
6.1. Globorotalia truncatulinoides: A Bio-Chronological Indicator
6.2. Globorotalia truncatulinoides: A Palaeoceanographic Tool
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aze, T.; Ezard, T.H.G.; Purvis, A.; Coxall, H.K.; Stewart, D.R.M.; Wade, B.S.; Pearson, P.N. A phylogeny of Cenozoic macroperforate planktonic foraminifera from fossil data. Biol. Rev. 2011, 63, 900–927. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.; Sprovieri, R.; Di Sefano, E.; Raffi, I. Globorotalia truncatulinoides (d’Orbigny) in the Mediterranean upper Pliocene geologic record. Micropaleontology 1984, 30, 121–137. [Google Scholar] [CrossRef]
- Walker, M.; Head, M.J.; Berkelhammer, M.; Björck, S.; Cheng, H.; Cwynar, L.; Fisher, D.; Gkinis, V.; Long, A.; Lowe, J.; et al. Formal ratification of the subdivision of the Holocene Series/Epoch (Quaternary System/Period): Two new Global Boundary Stratotype Sections and Points (GSSPs) and three new stages/subseries. Episodes 2018, 41, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Cacho, I.; Grimalt, J.O.; Canals, M.; Sbaffi, L.; Shackleton, N.J.; Schönfeld, J.; Zahn, R. Variability of the western Mediterranean Sea surface temperature during the last 25,000 years and its connection with the Northern Hemisphere climatic changes. Paleoceanogr. Paleoceanogr. 2001, 16, 40–52. [Google Scholar] [CrossRef]
- Margaritelli, G.; Vallefuoco, M.; di Rita, F.; Capotondi, L.; Bellucci, L.G.; Insinga, D.D.; Petrosino, P.; Bonomo, S.; Cacho, I.; Cascella, A.; et al. Climate events from a shallow water marine record of the Central Tyrrhenian during the last four millennia. Glob. Planet. Change 2016, 142, 53–72. [Google Scholar] [CrossRef]
- Margaritelli, G.; Cisneros, M.; Cacho, I.; Capotondi, L.; Vallefuoco, M.; Rettori, R.; Lirer, F. Climatic variability over the last 3000 years in the central–western Mediterranean Sea (Menorca Basin) detected by planktonic foraminifera and stable isotope records. Glob. Planet. Change 2018, 169, 179–187. [Google Scholar] [CrossRef]
- Margaritelli, G.; Cacho, I.; Català, A.; Barra, M.; Bellucci, L.G.; Lubritto, C.; Rettori, R.; Lirer, F. Persistent warm Mediterranean surface waters during the Roman period. Sci. Rep. 2020, 10, 10431. [Google Scholar] [CrossRef]
- Lirer, F.; Antonio, C.; Claudia, C.; Elena, T.; Francisco, S.; Salvatorini, G.; Turco, E.; Cosentino, C.; Sierro, F.J.; Caruso, A. Mediterranean Neogene planktonic foraminifer biozonation and biochronology. Earth Sci. Rev. 2019, 196, 102869. [Google Scholar] [CrossRef]
- Rohling, E.G.; Mayewski, P.A.; Abu-Zied, R.H.; Casford, J.S.L.; Hayes, A. Holocene atmosphere-ocean interactions: Records from Greenland and the Aegean Sea. Clim. Dyn. 2002, 18, 587–593. [Google Scholar]
- Trias-Navarro, S.; Cacho, I.; de la Fuente, M.; Pena, L.D.; Frigola, J.; Lirer, F.; Caruso, A. Surface hydrographic changes at the western flank of the Sicily Channel associated with the last sapropel. Glob. Planet. Change 2021, 204, 103582. [Google Scholar] [CrossRef]
- Magny, M.; Miramont, C.; Sivan, O. Assessment of the impact of climate and anthropogenic factors on Holocene Mediterranean vegetation in Europe on the basis of palaeohydrological records. Palaeogeography 2002, 186, 47–59. [Google Scholar] [CrossRef]
- Lirer, F.; Sprovieri, M.; Vallefuoco, M.; Ferraro, L.; Pelosi, N.; Giordano, L.; Capotondi, L. Planktonic foraminifera as bio-indicators for monitoring the climatic changes that have occurred over the past 2000 years in the south-eastern Tyrrhenian Sea. Integrativ. Zool. 2014, 9, 542–554. [Google Scholar] [CrossRef]
- Berger, J.F.; Lespez, L.; Kuzucuoğlu, C.; Glais, A.; Hourani, F.; Barra, A.; Guilaine, J. Interactions between climate change and human activities during the early to mid-Holocene in the eastern Mediterranean basins. Clim. Past. 2016, 12, 1847–1877. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.; Head, M.J.; Lowe, J.; Berkelhammer, M.; BjÖrck, S.; Cheng, H.; Cwynar, L.C.; Fisher, D.; Gkinis, V.; Long, A.; et al. Subdividing the Holocene Series/Epoch: Formalization of stages/ages and subseries/subepochs, and designation of GSSPs and auxiliary stratotypes. JQS 2019, 34, 3–173. [Google Scholar] [CrossRef]
- Ran, M.; Chen, L. The 4.2 ka BP climatic event and its cultural response. Quat. Int. Vol. 2019, 521, 158–167. [Google Scholar]
- Mayewski, P.A.; Rohling, E.; Stager, C.; Karlén, W.; Maasch, K.A.; Meeker, L.D.; Meyerson, E.A.; Gasse, F.; van Kreveld, S.; Holmgren, K.; et al. Holocene climate variability. Quat. Res. 2004, 62, 243–255. [Google Scholar] [CrossRef]
- Staubwasser, M.; Weiss, H. Holocene climate and cultural evolution in Late Prehistoric–Early Historic West Asia. Quat. Res. 2006, 66, 372–387. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.J.C.; Berkelhammer, M.; Bjorck, S.; Cwynar, L.C.; Fisher, D.A.; Long, A.J.; Lower, J.J.; Newnham, R.M.; Rasmussen, S.O.; Weiss, H. Formal subdivision of the Holocene series/epoch: A discussion paper by a working group of INTIMATE (integration of ice-core, marine and terrestrial records) and the subcommission on quaternary stratigraphy (International Commission on Stratigraphy). J. Quat. Sci. 2012, 27, 649–659. [Google Scholar] [CrossRef]
- Magri, D.; Parra, I. Late Quaternary western Mediterranean pollen records and African winds. Earth Planet. Sci. Lett. 2002, 200, 401–408. [Google Scholar] [CrossRef]
- Sangiorgi, F.; Capotondi, C.; Combourieu Nebout, N.; Vigliotti, L.; Brinkhuis, H.; Giunta, S.; Lotter, A.F.; Morigi, C.; Negri, A.; Reichart, G.J. Holocene seasonal sea-surface temperature variations in the southern Adriatic Sea inferred from a multiproxy approach. J. Quat. Sci. 2003, 18, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Drysdale, R.; Zanchetta, G.; Hellstrom, J.; Maas, R.; Fallick, A.; Pickett, M.; Cartwright, I.; Piccini, L. Late Holocene drought responsible for the collapse of Old-World civilizations is recorded in an Italian cave flowstone. Geology 2006, 34, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Di Rita, F.; Magri, D. The 4.2 ka event in the vegetation record of the central Mediterranean. Clim. Past. 2019, 15, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Combourieu-Nebout, N.; Bertini, A.; Russo-Ermolli, E.; Peyron, O.; Klotz, S.; Montade, V.; Fauquette, S.; Allen, J.; Fusco, F.; Goring, S.; et al. Climate changes in the central Mediterranean and Italian vegetation dynamics since the Pliocene. Rev. Palaeobot. Palynol. 2015, 218, 127–147. [Google Scholar] [CrossRef]
- Zanchetta, G.; Regattieri, E.; Isola, I.; Drysdale, R.N.; Bini, M.; Baneschi, I.; Hellstrom, J.C. The so-called “4.2 event” in the Central Mediterranean and its climatic teleconnections. Alp. Mediterr. Quat. 2016, 29, 5–17. [Google Scholar]
- Di Rita, F.; Lirer, F.; Bonomo, S.; Cascella, A.; Ferraro, L.; Florindo, F.; Insinga, D.D.; Lurcock, P.C.; Margaritelli, G.; Petrosino, P.; et al. Late Holocene forest dynamics in the Gulf of Gaeta (central Mediterranean) in relation to NAO variability and human impact. Quat. Sci. Rev. 2018, 179, 137–152. [Google Scholar] [CrossRef]
- Bini, M.; Zanchetta, G.; Perșoiu, A.; Cartier, R.; Català, A.; Cacho, I.; Dean, R.J.; Di Rita, F.; Drysdale, R.N.; Finnè, M.; et al. The 4.2 ka BP Event in the Mediterranean Region: An overview. Clim. Past 2019, 15, 555–577. [Google Scholar] [CrossRef] [Green Version]
- Català, A.; Cacho, I.; Frigola, J.; Pena, L.D.; Lirer, F. Holocene hydrography evolution in the Alboran Sea: A multi-record and multi-proxy comparison. Clim. Past 2019, 15, 927–942. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, S.; Sulli, A.; Di Stefano, E.; Giaramita, L.; Incarbona, A.; Mortyn, P.G.; Sprovieri, M.; Sprovieri, R.; Tonielli, R.; Vallefuoco, M.; et al. Late Quaternary palaeoenvironmental reconstruction of sediment drift accumulation in the Malta Graben (central Mediterranean Sea). Geo-Mar. Lett. 2018, 38, 241–258. [Google Scholar] [CrossRef]
- Lirer, F.; Sprovieri, M.; Ferraro, L.; Vallefuoco, M.; Capotondi, L.; Cascella, A.; Petrosino, P.; Insinga, D.D.; Pelosi, N.; Tamburrino, S.; et al. Integrated stratigraphy for the late Quaternary in the eastern Tyrrhenian Sea. Quat. Int. 2013, 292, 71–85. [Google Scholar] [CrossRef]
- Margaritelli, G.; Lirer, F.; Schroeder, K.; Alberico, I.; Dentici, M.P.; Caruso, A. Globorotalia truncatulinoides in Central—Western Mediterranean Sea during the Little Ice Age. Mar. Micropaleontol. 2020, 161, 101921. [Google Scholar] [CrossRef]
- Robinson, A.R.; Golnaraghi, M. The Physical and Dynamical Oceanography of the Mediterranean. In Ocean Processes in Climate Dynamics: Global and Mediterranean Examples; Malanotte-Rizzoli, P., Robinson, A.R., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 255–306. [Google Scholar]
- POEM Group. General circulation of the Eastern Mediterranean. Earth Sci. Rev. 1992, 32, 285–309. [Google Scholar] [CrossRef]
- Millot, C. Circulation in the Western Mediterranean Sea. J. Mar. Syst. 1999, 20, 423–442. [Google Scholar] [CrossRef] [Green Version]
- Béranger, K.; Mortier, L.; Gasparini, G.P.; Gervasio, L.; Astraldi, M.; Crépona, M. The dynamics of the Sicily Strait: A comprehensive study from observations and models. Deep-Sea Res. II 2004, 51, 411–440. [Google Scholar] [CrossRef]
- Vetrano, A.; Napolitano, E.; Iacono, R.; Schroeder, K.; Gasparini, G.P. Tyrrhenian Sea circulation and water mass fluxes in spring 2004: Observations and model results. J. Geophys. Res. 2010, 115, C06023. [Google Scholar] [CrossRef]
- D’Ortenzio, F.; Ribera d’Alcalà, M. On the trophic regimes of the Mediterranean Sea: A satellite analysis. Biogeosciences 2009, 6, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, K. Current Systems in the Mediterranean Sea. In Encyclopedia of Ocean Sciences, 3rd ed.; Cochran, J.K., Bokuniewicz, J.H., Yager, L.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 3, pp. 219–227. ISBN 978-0-12-813081-0. [Google Scholar]
- Kallel, N.; Paterne, M.; Labeyrie, L.D.; Duplessy, J.C.; Arnold, M. Temperature and salinity records of the Tyrrhenian Sea during the last 18,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 135, 97–108. [Google Scholar] [CrossRef]
- Kucera, M.; Weinelt, M.; Kiefer, T.; Pflaumann, U.; Hayes, A.; Weinelt, M.; Chen, M.; Mix, A.C.; Barrows, T.T.; Cortijo, E.; et al. Reconstruction of sea-surface temperatures from assemblages of planktonic foraminifera: Multi-technique approach based on geographically constrained calibration datasets and its application to glacial Atlantic and Pacific Oceans. Quat. Sci. Rev. 2005, 24, 951–998. [Google Scholar] [CrossRef]
- Rohling, E.J.; Hayes, A.; De Rijk, S.; Kroon, D.; Zachariasse, W.J.; Eisma, D. Abrupt cold spells in the northwest Mediterranean. Paleoceanography 1998, 13, 316–322. [Google Scholar] [CrossRef]
- Thunel, R.C. Distribution of recent planktonic foraminifera in surface sediments of the Mediterranean Sea. Mar. Micropaleont 1978, 3, 147–173. [Google Scholar] [CrossRef]
- Pujol, C.; Vergnaud Grazzini, C. Distribution patterns of live planktic foraminifers as related to regional hydrography and productive systems of the Mediterranean Sea. Mar. Micropaleontol. 1995, 25, 187–217. [Google Scholar] [CrossRef]
- Frigola, J.; Moreno, A.; Cacho, I.; Canals, M.; Sierro, F.J.; Flores, J.A.; Grimalt, J.O.; Hodell, D.A.; Curtis, J.H. Holocene climate variability in the western Mediterranean region from a deep water sediment record. Paleoceanography 2007, 22, PA2209. [Google Scholar] [CrossRef] [Green Version]
- Broggy, T.C. Planktonic foraminiferal response to the Last Glacial Termination and their application to Holocene biostratigraphy in the western Mediterranean Sea. PhD Thesis, Department of Geography, Mary Immaculate College, University of Limerick, Limerick, Ireland, 2011. [Google Scholar]
- Morabito, S.; Petrosino, P.; Milia, A.; Sprovieri, M.; Tamburrino, S. A multidisciplinary approach for reconstructing the stratigraphic framework of the last 40 ka in a bathyal area of the eastern Tyrrhenian Sea. Glob. Planet. Change 2014, 123, 121–138. [Google Scholar] [CrossRef]
- Di Stefano, A.; Foresi, L.M.; Incarbona, A.; Sprovieri, M.; Vallefuoco, M.; Iorio, M.; Pelosi, N.; Di Stefano, E.; Sangiorgi, P.; Budillon, F. Mediterranean coccolith ecobiostratigraphy since the penultimate Glacial (the last 145,000 years) and ecobioevent traceability. Mar. Micropaleontol. 2015, 115, 24–38. [Google Scholar] [CrossRef]
- Budillon, F.; Lirer, F.; Iorio, M.; Macrì, P.; Sagnotti, L.; Vallefuoco, M.; Ferraro, L.; Garziglia, S.; Innangi, S.; Sahabi, M.; et al. Integrated stratigraphic reconstruction for the last 80 kyr in a deep sector of the Sardinia Channel (Western Mediterranean). Deep-Sea Res. II 2009, 56, 725–737. [Google Scholar] [CrossRef] [Green Version]
- Capotondi, L.; Soroldoni, E.; Principato, M.S.; Corselli, C. Late Quaternary planktonic foraminiferal distributions: Problems related to size fraction. In Proceedings of the First Italian Meeting on Environmental Micropaleontology, Urbino, Italy, 4–6 June 2002; Coccioni, R., Galeotti, S., Lirer, F., Eds.; Grzybowski Foundation Special Publication: Kraków, Poland, 2004. [Google Scholar]
- Houpert, L.; Testor, P.; De Madron, X.D. Gridded climatology of the Mixed Layer (Depth and Temperature), the bottom of the Seasonal Thermocline (Depth and Temperature), and the upper-ocean Heat Storage Rate for the Mediterranean Sea. SEANOE 2015. Available online: http://nora.nerc.ac.uk/id/eprint/522296 (accessed on 9 May 2022).
- Houpert, L.; Testor, P.; de Madron, X.D.; Somot, S.; D’Ortenzio, F.; Estournel, C.; Lavigne, H. Seasonal cycle of the mixed layer, the seasonal thermocline and the upper ocean heat storage rate in the Mediterranean Sea derived from observations. Prog. Oceanogr. 2015, 132, 333–352. [Google Scholar] [CrossRef]
- Emiliani, C. Depth habitats of some species of pelagic foraminifera as indicated by oxygen isotope ratios. Am. J. Sci. 1954, 252, 149–158. [Google Scholar] [CrossRef]
- Bé, A.W.H. Ecology of recent planktonic foraminifera. Bathymetric and seasonal distribution in the Sargasso Sea off Bermuda. Micropaleontology 1960, 6, 373–392. [Google Scholar] [CrossRef]
- Bé, A.W.H.; Ericson, D.B. Aspect of calcification in planktonic Foraminifera (Sarcodinia). Ann. N. Y. Acad. Sci. 1963, 109, 81–97. [Google Scholar]
- Bé, A.W.H.; Tolderlund, D.S. Distribution and ecology of living foraminifera in surface waters of the Atlantic and Indian oceans. In The Micropaleontology of the Oceans; Funnel, B.M., Riedel, W.R., Eds.; Cambridge University Press: London, UK, 1971; pp. 104–105. [Google Scholar]
- Deuser, W.; Ross, E. Seasonally abundant planktonic foraminifera of the Sargasso Sea; succession, deep-water fluxes, isotopic compositions, and paleoceanographic implications. J. Foraminifera Res. 1989, 19, 268–293. [Google Scholar] [CrossRef]
- Lohmann, G.P.; Schweitzer, P.N. Globorotalia truncatulinoides Growth and chemistry as probes of the past thermocline: 1. Shell size. Paleoceanography 1990, 5, 55–75. [Google Scholar] [CrossRef]
- Le Grande, A.N.; Lynch-Stieglitz, J.; Farmer, E.C. Oxygen isotopic composition of Globorotalia truncatulinoides as a proxy for intermediate depth density. Paleoceanography 2004, 19, PA4025. [Google Scholar]
- Schiebel, R.; Waniek, J.; Zeltner Alves, M. Impact of the Azores Front on the distribution of planktic foraminifers, shelled gastropods, and coccolithophorids. Deep-Sea Res. II 2002, 49, 4035–4050. [Google Scholar] [CrossRef]
- Spear, J.W.; Poore, R.Z.; Quinn, T.M. Globorotalia truncatulinoides (dextral) Mg/Ca as a proxy for Gulf of Mexico winter mixed-layer temperature: Evidence from a sediment trap in the northern Gulf of Mexico. Mar. Micropaleontol. 2011, 80, 53–56. [Google Scholar] [CrossRef]
- Bè, A.W.H.; Lott, L. Shell growth and structure of planktonic foraminifera. Science 1964, 145, 823–824. [Google Scholar] [CrossRef]
- Orr, W.N. Secondary Calcification in the Foraminiferal Genus Globorotalia. Sci. New Ser. 1967, 157, 1554–1555. [Google Scholar] [CrossRef]
- Mulitza, S.; Dürkoop, A.; Hale, W.; Wefer, G.; Niebler, H.S. Planktonic foraminifera as recorders of past surface-water stratification. Geology 1997, 25, 335–338. [Google Scholar] [CrossRef]
- Wilke, I.; Meggers, H.; Bickert, T. Depth habitats and seasonal distributions of recent planktic foraminifers in the Canary Islands region (29°N) based on oxygen isotopes. Deep-Sea Res. 2009, 1, 89–106. [Google Scholar] [CrossRef]
- De Vargas, C.; Renaud, S.; Hilbrecht, H.; Pawlowski, J. Pleistocene adaptive radiation in Globorotalia truncatulinoides: Genetic, morphologic, and environmental evidence. Paleobiology 2001, 27, 104–125. [Google Scholar] [CrossRef]
- Ujiié, Y.; Lipps, J.H. Cryptic diversity in planktic foraminifera in the northwest Pacific Ocean. J. Foraminifera Res. 2009, 39, 145–154. [Google Scholar] [CrossRef]
- Renaud, S.; Schmidt, D.N. Habitat tracking as a response of the planktic foraminifer Globorotalia truncatulinoides to environmental fluctuations during the last 140 kyr. Mar. Micropaleontol. 2003, 49, 97–122. [Google Scholar] [CrossRef]
- Darling, K.F.; Wade, C.M. The genetic diversity of planktic foraminifera and the and the global distribution of ribosomal RNA genotypes. Mar. Micropalaeontol. 2008, 67, 216–238. [Google Scholar] [CrossRef]
- Lohmann, G.P. Increasing seasonal upwelling in the subtropical South Atlantic over the past 700,000 years: Evidence from deep-living planktonic foraminifera. Mar. Micropaleontol. 1992, 19, 1–12. [Google Scholar] [CrossRef]
- Sprovieri, R.; Di Stefano, E.; Incarbona, A.; Gargano, M.E. A high-resolution record of the last deglaciation in the Sicily Channel based on foraminifera and calcareous nannofossil quantitative distribution. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 202, 119–142. [Google Scholar] [CrossRef]
- Schiebel, R.; Hemleben, C. Planktic Foraminifers in the Modern Ocean; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- De Castro Coppa, M.G.; Moncharmont Zei, M.; Placella, B.; Sgarrella, F.; Taddei Ruggiero, E. Distribuzione stagionale e verticale dei Foraminiferi planctonici del Golfo di Napoli. Boll. Soc. Nat. Napoli 1980, 89, 1–25. [Google Scholar]
- Rigual-Hernández, A.; Sierro, F.J.; Bárcena, M.A.; Flores, J.A.; Heussner, S. Seasonal and interannual changes of planktic foraminiferal fluxes in the Gulf of Lions (NW Mediterranean) and their implications for paleoceanographic studies: Two 12- year sediment trap records. Deep-Sea Res. I 2012, 66, 26–40. [Google Scholar] [CrossRef]
- Vergnaud Grazzini, C. Nonequilibrium isotopic compositions of shells of planktonic foraminifera in the Mediterranean Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1976, 20, 263–276. [Google Scholar] [CrossRef]
- Avnaim-Katav, S.; Herut, B.; Rahav, E.; Katz, T.; Weinstein, Y.; Alkalay, R.; Berman-Frank, I.; Zlatkin, O.; Almogi-Labin, A. Sediment trap and deep sea coretop sediments as tracers of recent changes in planktonic foraminifera assemblages in the southeastern ultra-oligotrophic Levantine Basin. Deep-Sea Res. II. 2020, 171, 104669. [Google Scholar] [CrossRef]
- Bé, A.W.H. Ecology of Recent planktonic foraminifera: Part 1- Areal distribution in the western North Atlantic. Micropaleontology 1959, 5, 77–100. [Google Scholar] [CrossRef]
- Tolderlund, D.S.; Bé, A.W.H. Seasonal distribution of planktonic foraminifera in the western North Atlantic. Micropaleontology 1971, 17, 297–329. [Google Scholar] [CrossRef]
- Schmuker, B.; Schiebel, R. Planktic foraminifers and hydrography of the eastern and northern Caribbean Sea. Mar. Micropaleontol. 2002, 46, 387–403. [Google Scholar] [CrossRef]
- Geraga, M.; Mylona, G.; Tsaila-Monopoli, S.; Papatheodorou, G.; Ferentinos, G. Northeastern Ionian Sea: Palaeoceanographic variability over the last 22 ka. J. Mar. Syst. 2008, 74, 623–638. [Google Scholar] [CrossRef]
- Siani, G.; Paterne, M.; Colin, C. Late glacial to Holocene planktic foraminifera bio events and climatic record in the South Adriatic Sea. J. Quat. Sci. 2010, 25, 808–821. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Antonarakou, A.; Kouli, K.; Dimiza, M.; Kontakiotis, G.; Papanikolaou, M.D.; Ziveri, P.; Mortyn, P.G.; Lianou, V.; Lykousis, V.; et al. Late Glacial-Holocene ecostratigraphy of the south-eastern Aegean Sea, based on plankton and pollen assemblages. Geo-Mar. Lett. 2009, 29, 249–267. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Antonarakou, A.; Zachariasse, W.J. Late Quaternary palaeoenvironmental changes in the Aegean Sea: Interrelations and interactions between North and South Aegean Sea. Bull. Geol. Soc. Greece 2013, 47, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Kontakiotis, G. Late Quaternary paleoenvironmental reconstruction and paleoclimatic implications of the Aegean Sea (eastern Mediterranean) based on paleoceanographic indexes and stable isotopes. Quat. Int. 2016, 401, 28–42. [Google Scholar] [CrossRef] [Green Version]
- Giamali, C.; Koskeridou, E.; Antonarakou, A.; Ioakim, C.; Kontakiotis, G.; Karageorgis, A.P.; Roussakis, G.; Karakitsios, V. Multiproxy ecosystem response of abrupt Holocene climatic changes in the north-eastern Mediterranean sedimentary archive and hydrologic regime. Quat. Res. 2019, 92, 665–685. [Google Scholar] [CrossRef]
- Rohling, E.J.; Jorissen, F.J.; Vergnaud-Grazzini, C.; Zachariasse, W.J. Northern Levantine and Adriatic Quaternary planktic foraminifera: Reconstruction of paleoenvironmental gradients. Mar. Micropaleontol. 1993, 21, 191–218. [Google Scholar] [CrossRef]
- Lazarus, D.; Hilbrecht, H.; Spencer-Cervato, C.; Thierstein, H. Sympatric Speciation and Phyletic Change in Globorotalia truncatulinoides. Paleobiology 1995, 21, 28–51. [Google Scholar] [CrossRef]
- Spencer-Cervato, C.; Thierstein, H.R. First appearance of Globorotalia truncatulinoides: Cladogenesis and immigration. Mar. Micropaleontol. 1997, 30, 267–291. [Google Scholar] [CrossRef]
- Lourens, L.; Hilgen, F.; Shackleton, N.J.; Laskar, J.; Wilson, D. The Neogene Period. In A Geologic Time Scale; Gradstein, F.M., Ogg, J.G., Smith, A., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 409–440. [Google Scholar]
- Boscolo Galazzo, F.; Jones, A.; Jones, T.D.; Crichton, K.S.; Wade, B.S.; Pearson, P.N. Late Neogene evolution of modern deep-dwelling plankton. Biogeosciences 2022, 19, 743–762. [Google Scholar] [CrossRef]
- Sexton, P.F.; Norris, R.D. Dispersal and biogeography of marine plankton: Long distance dispersal of the foraminifer Truncorotalia truncatulinoides. Geology 2008, 36, 899–902. [Google Scholar] [CrossRef]
- Pharr, R.B.; Williams, D.F. Shape changes in Globorotalia truncatulinoides as a function of ontogeny and paleobiogeography in the Southern Ocean. Mar. Micropaleontol. 1987, 12, 343–355. [Google Scholar] [CrossRef]
- Kennett, J.P. Pleistocene paleoclimates and foraminiferal biostratigraphy in subantarctic deep-sea cores. Deep-Sea Res. 1970, 17, 125–140. [Google Scholar] [CrossRef]
- Cita, M.B.; Gartner, S. Studi sul Pliocene e gli strati di passaggio dal Miocene al Pliocene. IV. The stratotype Zanclean foraminiferal and nannofossil biostratigraphy. Riv. Ital. Paleontol. E Stratigr. 1973, 79, 503–558. [Google Scholar]
- Ruggieri, G.; Sprovieri, R. A revision of Italian Pleistocene Stratigraphy. Geol. Romana 1977, 16, 131–139. [Google Scholar]
- Di Stefano, E.; Sprovieri, R.; Caruso, A. High resolution biochronology in the Monte Narbone Formation of the Capo Rossello section and the Mediterranean first occurrence of Globorotalia truncatulinoides. Riv. Ital. Paleontol. E Stratigr. 1993, 99, 357–370. [Google Scholar]
- Caruso, A. Climatic Changes during Upper Pliocene/Lower Pleistocene at Capo Rossello (Sicily, Italy): A Planktic Foraminifera Approach. In Proceedings of the First Italian Meeting on Environmental Micropaleontology, Urbino, Italy, 4–6 June 2002; Coccioni, R., Galeotti, S., Lirer, F., Eds.; Special Volume Grzybowski Foundation. The Grzybowski Foundation: Kraków, Poland, 2004; Volume 9, pp. 17–36. [Google Scholar]
- Capotondi, L.; Borsetti, A.M.; Vergnaud-Grazzini, C. Biostratigraphie et Composition Isotopique de Foraminifères Planctiques des Derniers 140000 ans dans la Région de Minorque (Méditerranée Occidentale). Comptes Rendus Académie Sci.—Ser. IIA—Earth Planet. Sci. 1987, 305, 493–498. [Google Scholar]
- Ducassou, E.; Capotondi, L.; Murat, A.; Bernasconi, S.M.; Mulder, T.; Gonthier, E.; Migeon, S.; Duprat, J.; Giraudeau, J.; Mascle, J. Multiproxy Late Quaternary stratigraphy of the Nile deep-sea turbidite system—Towards a chronology of deep-sea terrigenous systems Sediment. Geology 2007, 200, 1–13. [Google Scholar]
- Crundwell, M.P.; Woodhouse, A. A detailed biostratigraphic framework for 0–1.2 Ma Quaternary sediments of north-eastern Zealandia. N. Z. J. Geol. Geophys. 2022; 1–14. [Google Scholar] [CrossRef]
- Capotondi, L.; Borsetti, A.M.; Morigi, C. Foraminiferal ecozones, a high-resolution proxy for the Late Quaternary biochronology in the Central Mediterranean Sea. Mar. Geol. 1999, 153, 253–274. [Google Scholar] [CrossRef]
- Di Donato, V.; Insinga, D.; Iorio, M.; Molisso, F.; Rumolo, P.; Cardines, C.; Passaro, S. The palaeoclimatic and palaeoceanographic history of the Gulf of Taranto (Mediterranean Sea) in the last 15 ky. Glob. Planet. Change 2019, 172, 278–297. [Google Scholar] [CrossRef]
- Mojtahid, M.; Manceau, R.; Schiebel, R.; Hennekam, R.; de Lange, G.J. Thirteen thousand years of southeastern Mediterranean climate variability inferred from an integrative planktic foraminiferal-based approach. Paleoceanography 2015, 30, 402–422. [Google Scholar] [CrossRef]
- Rouis-Zargouni, I.; Turon, J.L.; Londeix, L.; Essallami, L.; Kallel, N.; Sicre, M.A. Environmental and climatic changes in the central Mediterranean Sea (Siculo–Tunisian Strait) during the last 30 ka based on dinoflagellate cyst and planktonic foraminifera assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 285, 17–29. [Google Scholar] [CrossRef]
- Incarbona, A.; Jonkers, L.; Ferraro, S.; Sprovieri, R.; Tranchida, G. Sea surface temperatures and paleoenvironmental variability in the Central Mediterranean during historical times reconstructed using planktonic foraminifera. Paleoceanogr. Paleoclimatol. 2019, 34, 394–408. [Google Scholar] [CrossRef]
- Buccheri, G.; Capretto, G.; Di Donato, V.; Esposito, P.; Ferruzza, G.; Pescatore, T.; Russo Ermolli, E.; Senatore, M.R.; Sprovieri, M.; Bertoldo, M.; et al. A high-resolution record of the last deglaciation in the southern Tyrrhenian Sea: Environmental and climatic evolution. Mar. Geol. 2002, 186, 447–470. [Google Scholar] [CrossRef]
- Amore, O.F.; Caffau, M.; Massa, B.; Morabito, S. Late Pleistocene- Holocene paleoclimate and related paleoenvironmental changes as recorded by calcareous nannofossils and planktonic foraminifera assemblages in the southern Tyrrhenian Sea (Cape Palinuro, Italy). Mar. Micropaleontol. 2004, 52, 255–276. [Google Scholar] [CrossRef]
- Sbaffi, L.; Wezel, F.C.; Curzi, G.; Zoppi, U. Millennial- to centennial-scale palaeoclimate variations during Termination I and the Holocene in the central Mediterranean Sea. Glob. Planet. Change 2004, 40, 201–217. [Google Scholar] [CrossRef]
- Di Bella, L.; Frezza, V.; Bergamin, L.; Carboni, M.G.; Falese, F.; Martorelli, E.; Tarragoni, C.; Chiocci, F.L. Foraminiferal record and high-resolution seismic stratigraphy of the Late Holocene succession of the submerged Ombrone River delta (Northern Tyrrhenian Sea, Italy). Quat. Int. 2014, 328–329, 287–300. [Google Scholar] [CrossRef]
- Checa, H.; Margaritelli, G.; Pena, L.D.; Frigola, J.; Cacho, I.; Rettori, R.; Lirer, F. High resolution paleo-environmental changes during the Sapropel 1 in the North Ionian Sea, central Mediterranean. Holocene 2020, 30, 11. [Google Scholar] [CrossRef]
- Ehrmann, W.; Schmiedl, G.; Beuscher, S.; Krüger, S. Intensity of African Humid Periods Estimated from Saharan Dust Fluxes. PLoS ONE 2017, 12, e0170989. [Google Scholar] [CrossRef] [Green Version]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical Controls and Feedbacks on Ocean Primary Production. Science 1998, 281, 200–207. [Google Scholar] [CrossRef] [Green Version]
- D’Ortenzio, F.; Iudicone, D.; de Boyer Montegut, C.; Testor, P.; Antoine, D.; Marullo, S.; Santoleri, R.; Madec, G. Seasonal variability of the mixed layer depth in the Mediterranean Sea as derived from in situ profiles. Geophys. Res. Lett. 2005, 32, 12. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, K.; Garcìa-Lafuente, J.; Josey, S.A.; Artale, V.; Nardelli, B.B.; Carrillo, A.; Gačić, M.; Gasparini, G.P.; Herrmann, M.; Lionello, P.; et al. Circulation of the mediterranean sea and its variability. Clim. Medit. Reg. 2012, 3, 187–256. [Google Scholar]
- Smith, R.O.; Bryden, H.L.; Stansfield, K. Observations of new western Mediterranean deep-water formation using Argo floats 2004–2006. Ocean Sci. 2008, 4, 133–149. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.; Saito, K.; Entekhabi, D. The role of the Siberian high in northern hemisphere climate variability. Geophys. Res. Lett. 2001, 28, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Rîmbu, N.; Lohmann, G.; Ionita, M. Interannual to multidecadal Euro-Atlantic blocking variability during winter and its relationship with extreme low temperatures in Europe. J. Geophys. Res. 2014, 119, 13621–13636. [Google Scholar] [CrossRef] [Green Version]
- Ionita, M.; Badăluţă, C.A.; Scholz, P.; Chelcea, S. Vanishing river ice cover in the lower part of the Danube basin—Signs of a changing climate. Sci. Rep. 2018, 8, 7948. [Google Scholar] [CrossRef]
- Perşoiu, A.; Ionita, M.; Harvey Weiss, H. Atmospheric blocking induced by the strengthened Siberian High led to drying in west Asia during the 4.2 ka BP event– a hypothesis. Clim. Past 2019, 15, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Pallacks, S.; Ziveri, P.; Martrat, B.; Mortyn, P.G.; Grelaud, M.; Schiebel, R.; Incarbona, A.; Garcia-Orellana, J.; Anglada-Ortiz, G. Planktic foraminiferal changes in the western Mediterranean Anthropocene. Glob. Planet. Change 2021, 204, 103549. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Core | Area | Water Depth (m) | Coordinates | References |
|---|---|---|---|---|
| M40/4-82-2SL | Gulf of Lion (GoL) | 1079 | 42°18.51′ N, 03°46.40′ E | Broggy PhD thesis |
| MD99-2343 | North Balearic (NB) | 2391 | 40°29.84′ N, 04°01.69′ E | This work (Català et al., 2019 age model) |
| ODP975B | South Balearic (SB) | 2415 | 38°53.78′ N, 04°30.59′ E | Broggy PhD thesis |
| CET1 | South Tyrrhenian Sea (ST) | 2088 | 39°54.69′ N, 14°06.65′ E | Morabito et al., 2014 |
| C33 | South Tyrrhenian Sea (ST) | 2368 | 38°39.48′ N, 10°20.98′ E | Di Stefano et al., 2015 |
| C08 | South Tyrrhenian Sea (ST) | 2370 | 38°38.53′ N, 10°21.55′ E | Budillon et al., 2009 |
| ODP 974B | Central Tyrrhenian Sea (CT) | 3453 | 40° 21.36′ N, 12°08.51′ E | Broggy PhD thesis |
| M40/4 80 SL | Tyrrhenian Sea (TS) | 1881 | 40°57.31′ N, 11°00.22′ E | Broggy PhD thesis |
| C90 | South Tyrrhenian Sea (TC) | 103 | 40°35.76′ N, 14°42.48′ E | Lirer et al., 2013 |
| ND11 | Sicily Channel (SC) | 475 | 37°01′ N, 13°10′ E | Margaritelli et al., 2020 |
| NDT6 | West Sicily (WS) | 1066 | 38°0′ N, 11°47′ E | Trias-Navarro et al., 2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margaritelli, G.; Lirer, F.; Schroeder, K.; Cloke-Hayes, A.; Caruso, A.; Capotondi, L.; Broggy, T.; Cacho, I.; Sierro, F.J. Globorotalia truncatulinoides in the Mediterranean Basin during the Middle–Late Holocene: Bio-Chronological and Oceanographic Indicator. Geosciences 2022, 12, 244. https://doi.org/10.3390/geosciences12060244
Margaritelli G, Lirer F, Schroeder K, Cloke-Hayes A, Caruso A, Capotondi L, Broggy T, Cacho I, Sierro FJ. Globorotalia truncatulinoides in the Mediterranean Basin during the Middle–Late Holocene: Bio-Chronological and Oceanographic Indicator. Geosciences. 2022; 12(6):244. https://doi.org/10.3390/geosciences12060244
Chicago/Turabian StyleMargaritelli, Giulia, Fabrizio Lirer, Katrin Schroeder, Angela Cloke-Hayes, Antonio Caruso, Lucilla Capotondi, Teresa Broggy, Isabel Cacho, and Francisco J. Sierro. 2022. "Globorotalia truncatulinoides in the Mediterranean Basin during the Middle–Late Holocene: Bio-Chronological and Oceanographic Indicator" Geosciences 12, no. 6: 244. https://doi.org/10.3390/geosciences12060244