
New Dinosaur Ichnological, Sedimentological, and Geochemical Data from a Cretaceous High-Latitude Terrestrial Greenhouse Ecosystem, Nanushuk Formation, North Slope, Alaska

 ,
, 
 , and
, and
Abstract
1. Introduction
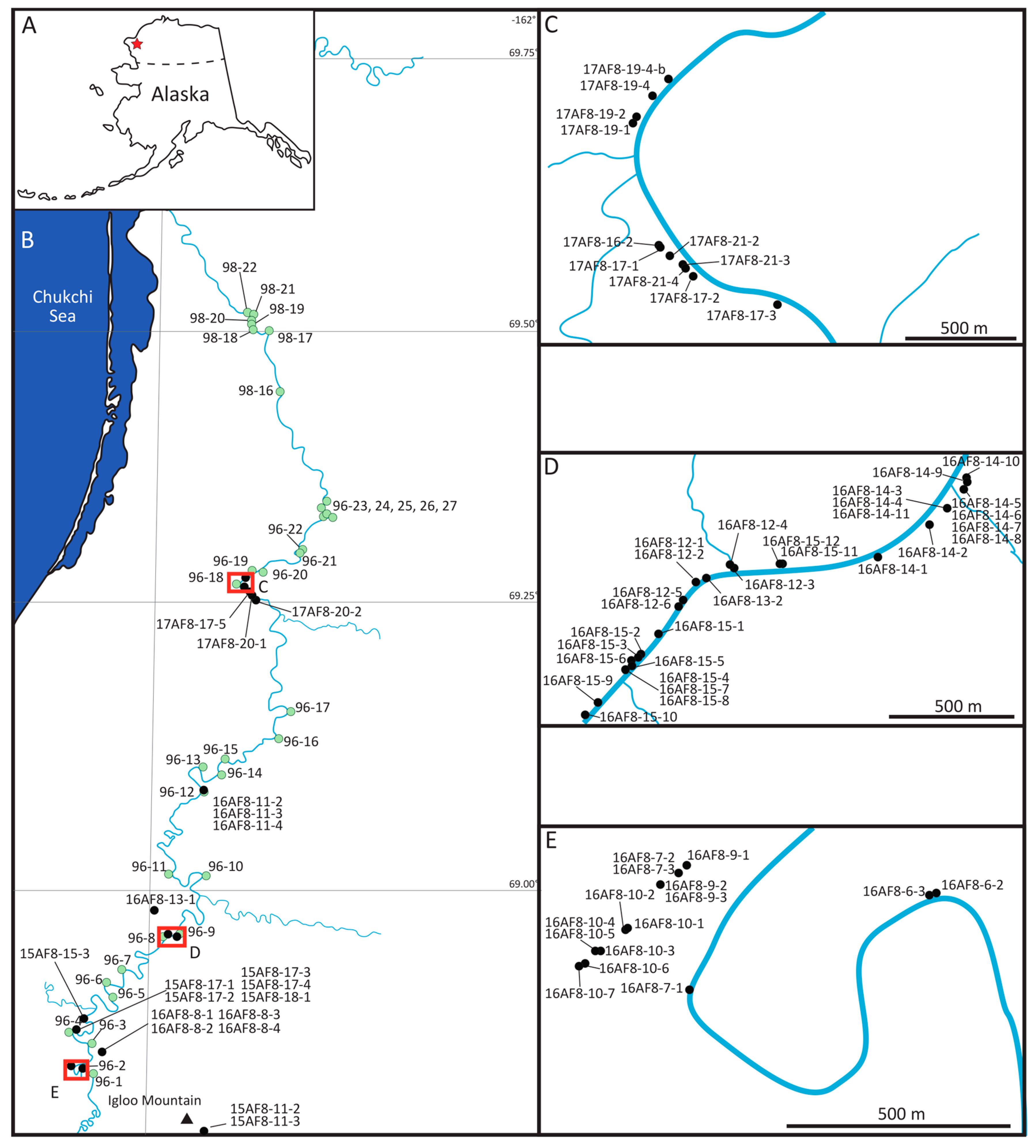
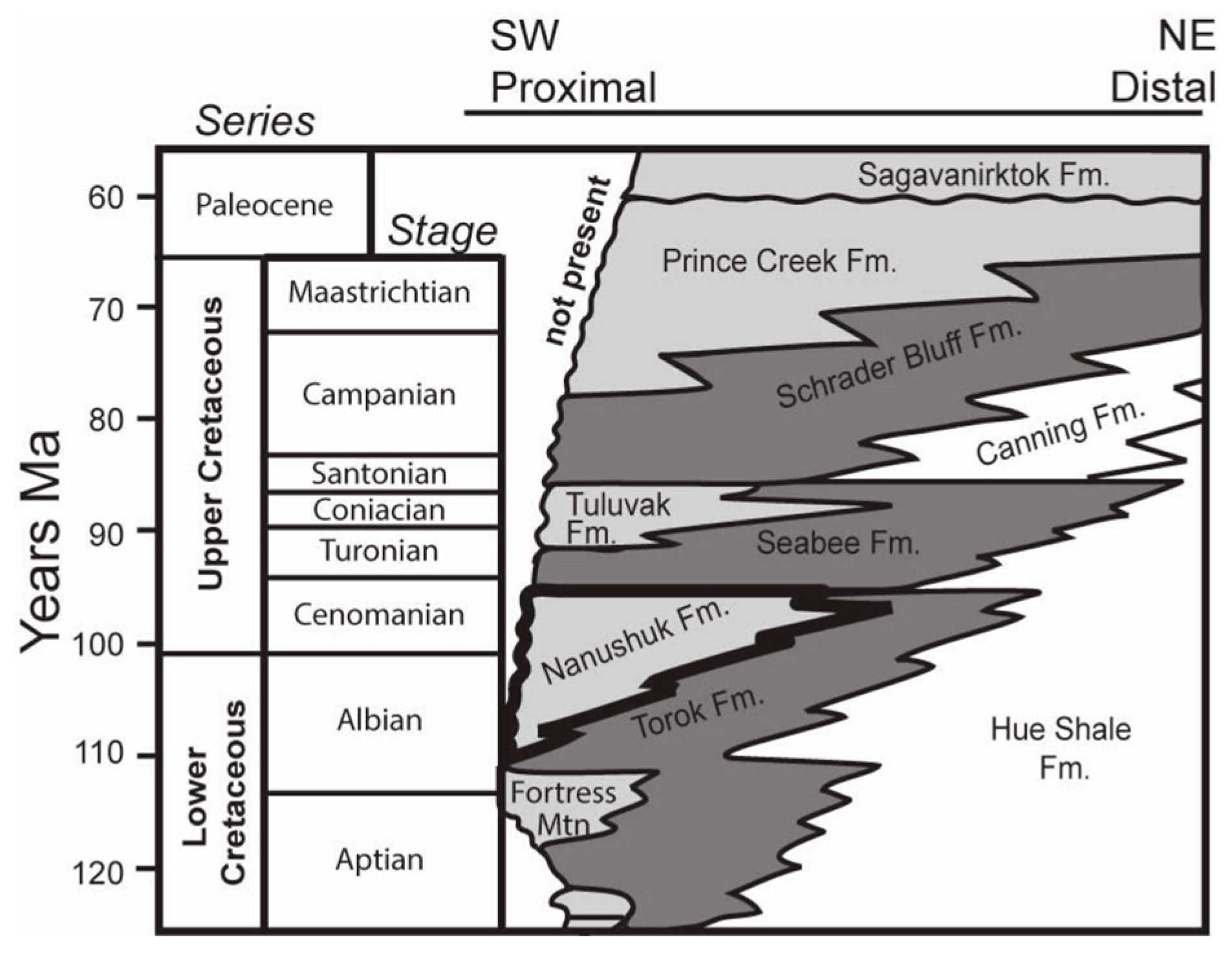
2. Geologic Framework
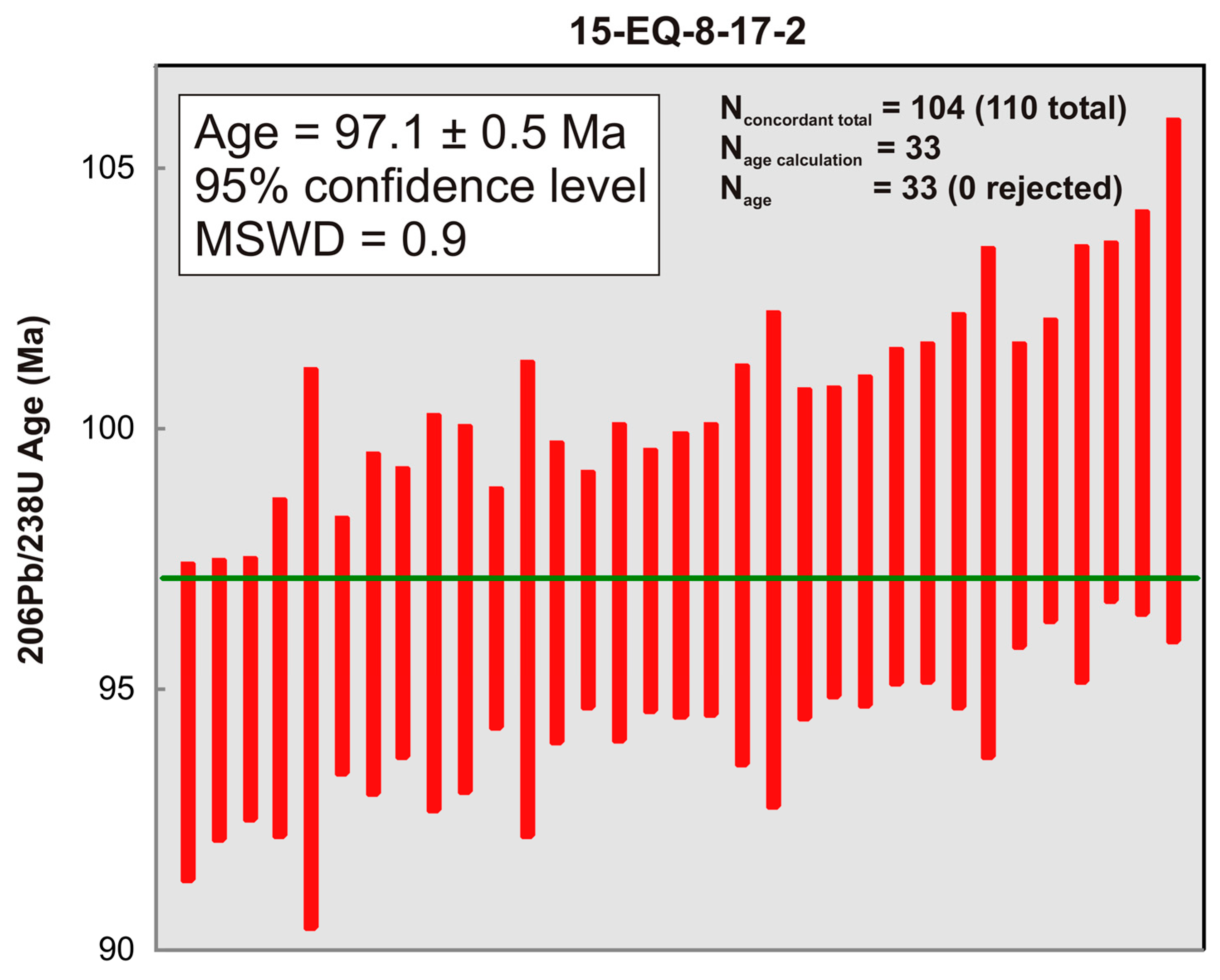
3. Radiometric Data from Study Area
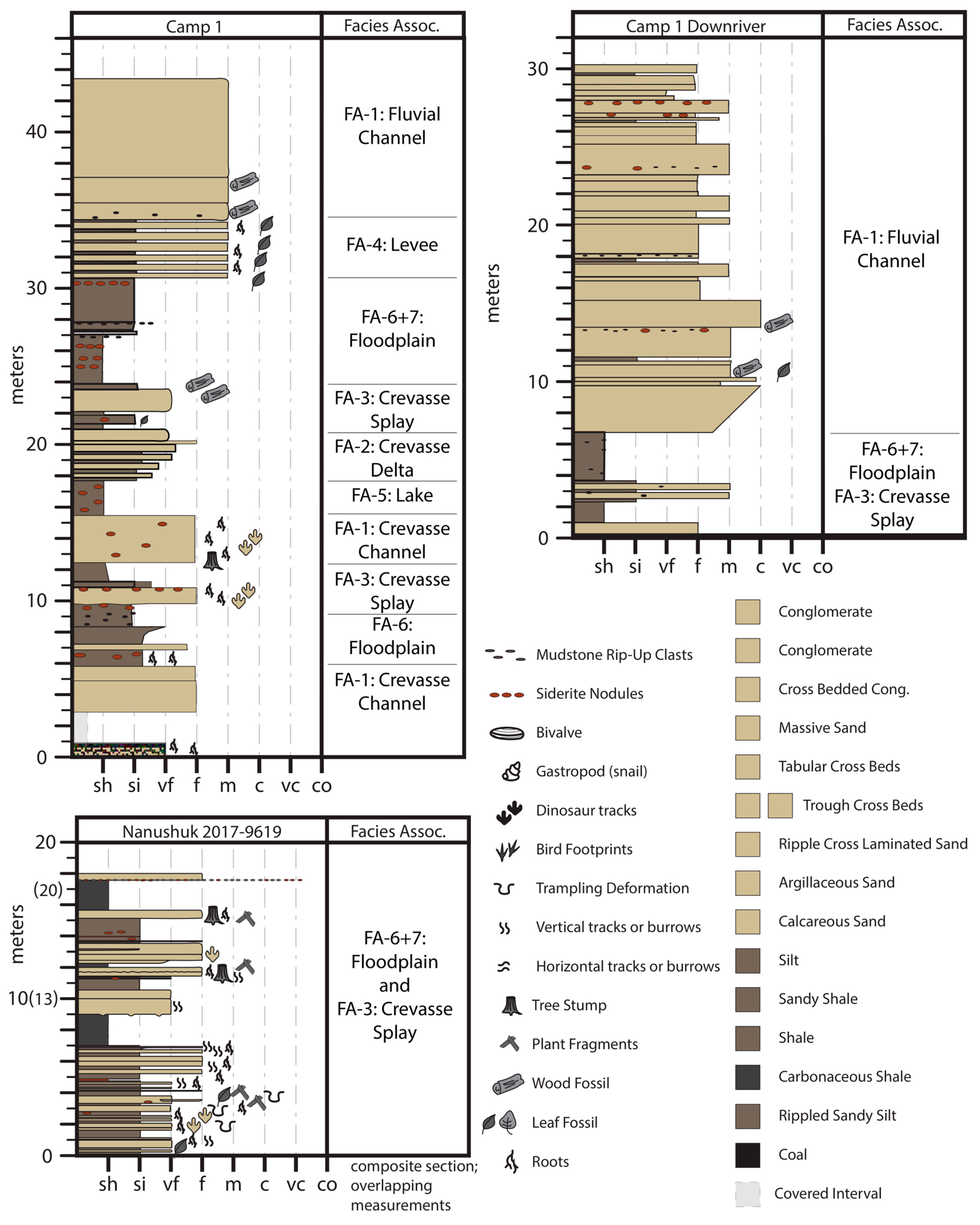
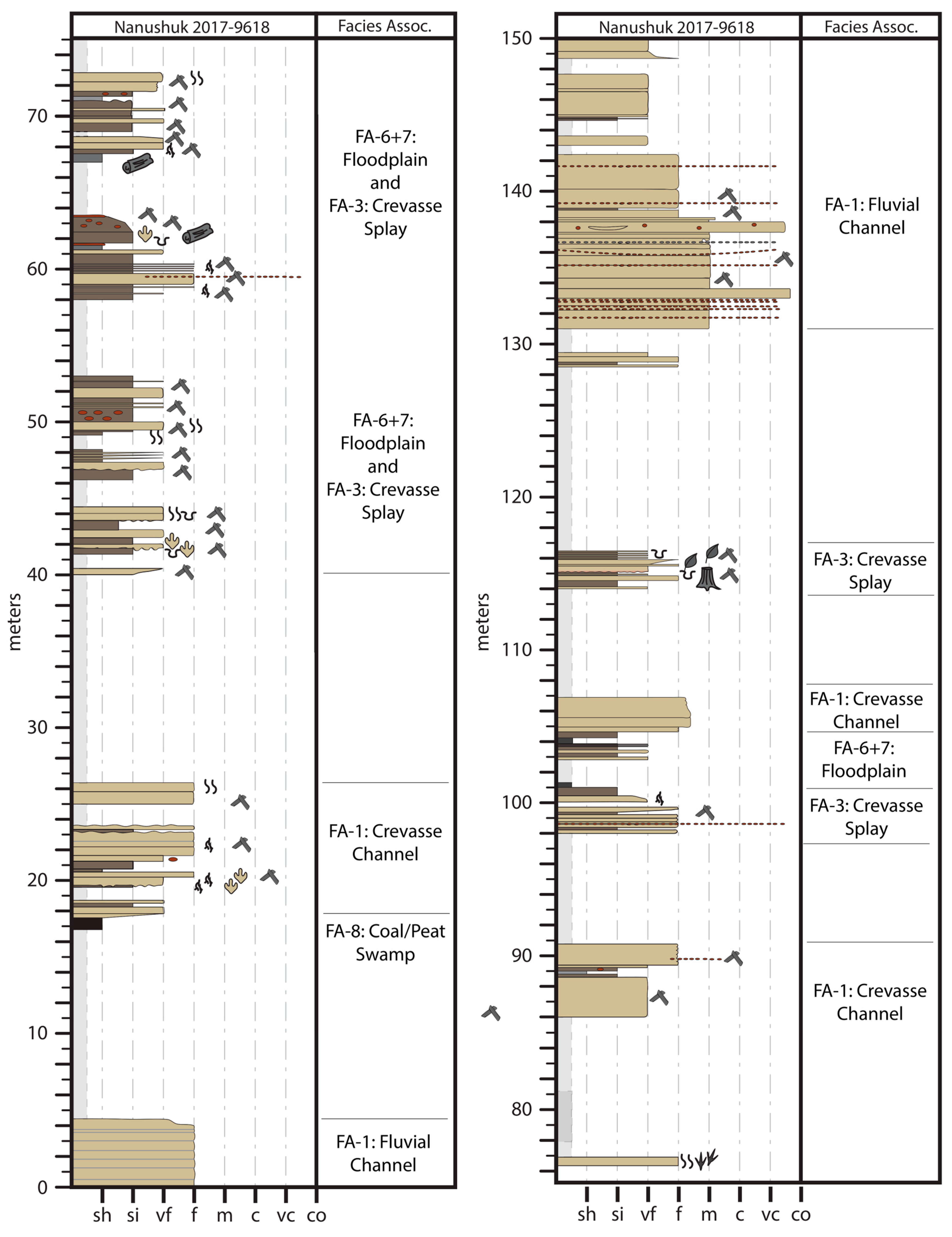
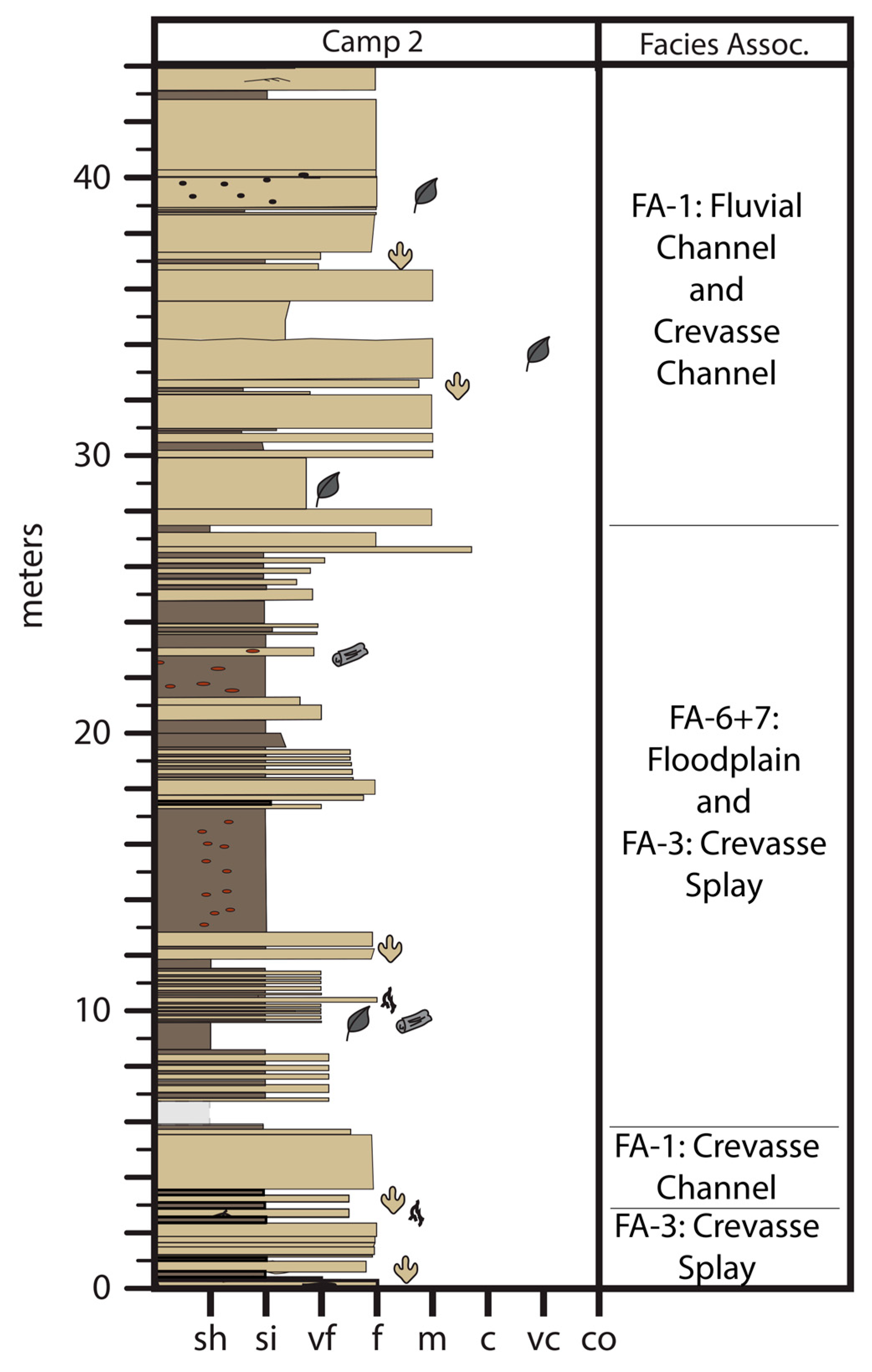
4. Sedimentology
4.1. Facies Association 1: Thick Sandstone
4.2. Interpretation
4.3. Facies Association 2: Interbedded Sandstone and Mudstone
4.4. Interpretation
4.5. Facies Association 3: Tabular or Lenticular Sharp-Based Sandstone
4.6. Interpretation
4.7. Facies Association 4: Rooted Siltstone and Sandstone
4.8. Interpretation
4.9. Facies Association 5: Platy Mudstone and Siltstone
4.10. Interpretation
4.11. Facies Association 6: Dark Brown to Brownish-Grey, Blocky Mudstone
4.12. Interpretation
4.13. Facies Association 7: Pale Grey, Sideritic Mudstone
4.14. Interpretation
4.15. Facies Association 8: Coal
4.16. Interpretation
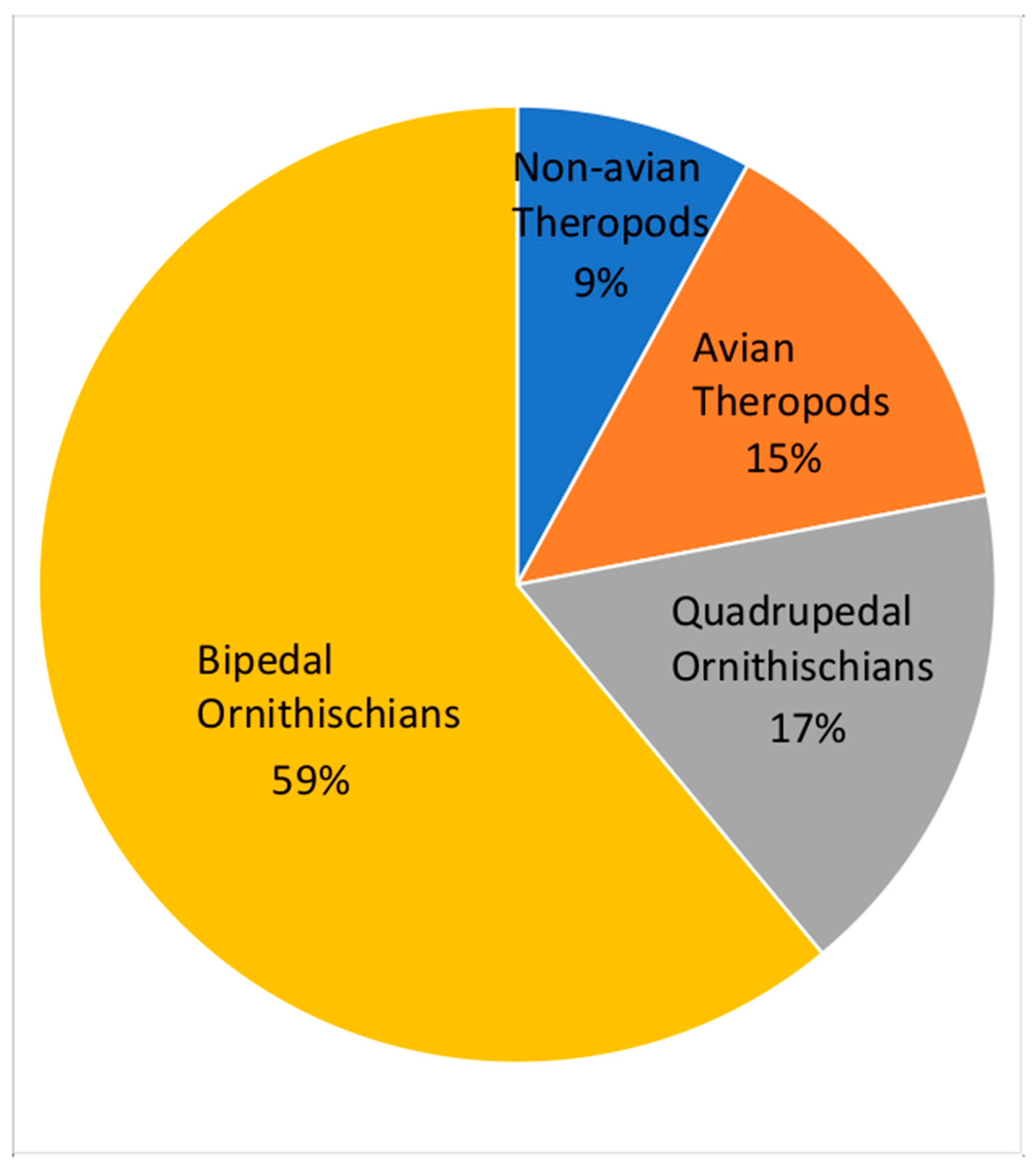
5. Fossil Vertebrates
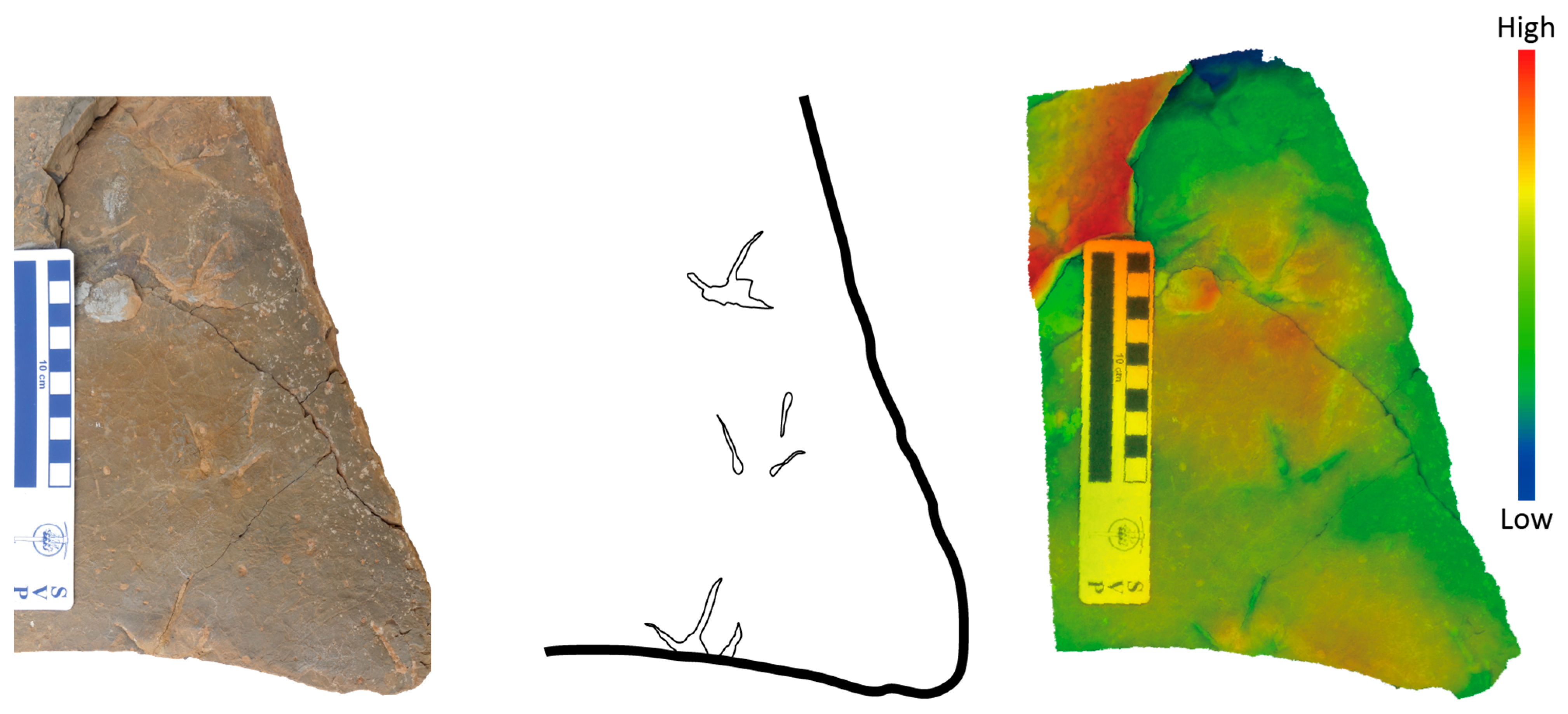
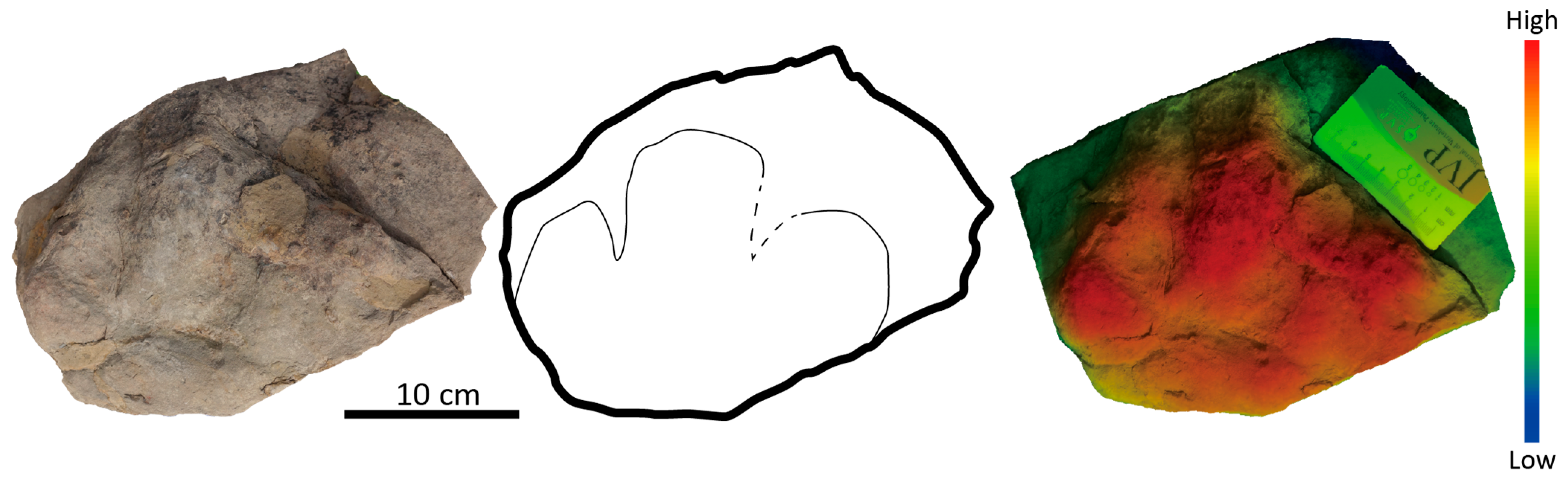
5.1. Non-Avian Theropods Tracks
5.2. Avian Theropod Tracks
5.3. Ornithischian Tracks
6. Paleoclimate
7. Discussion
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnosky, A.D.; Hadly, E.A.; Gonzalez, P.; Head, J.; Polly, P.D.; Lawing, A.M.; Eronen, J.T.; Ackerly, D.D.; Alex, K.; Biber, E.; et al. Merging paleobiology with conservation biology to guide the future of terrestrial ecosystems. Science 2017, 355, eaah4787. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Rodríguez-Sánchez, F.; Orsini, L.; de Boer, E.; Jansson, R.; Morlon, H.; Fordham, D.A.; Jackson, S.T. Cracking the code of biodiversity responses to past climate change. Trends Ecol. Evol. 2018, 33, 765–776. [Google Scholar] [CrossRef]
- Williams, J.W.; Ordonez, A.; Svenning, J.C. A unifying framework for studying and managing climate-driven rates of ecological change. Nat. Ecol. Evol. 2021, 5, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, W.; Smith, J.A.; Raja, N.B. Improving the relevance of paleontology to climate change policy. Proc. Natl. Acad. Sci. USA 2023, 120, e2201926119. [Google Scholar] [CrossRef]
- Parrish, J.T.; Spicer, R.A. Late Cretaceous terrestrial vegetation: A near-polar temperature curve. Geology 1988, 16, 22–25. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; McCarthy, P.J.; Kobayashi, Y.; Suarez, M.B. Cretaceous dinosaurs across Alaska show the role of paleoclimate in structuring ancient large-herbivore populations. Geosciences 2022, 12, 161. [Google Scholar] [CrossRef]
- Hedges, S.B.; Parker, P.H.; Sibley, C.G.; Kumar, S. Continental breakup and the ordinal diversification of birds and mammals. Nature 1996, 381, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Dilcher, D. Toward a new synthesis: Major evolutionary trends in the angiosperm fossil record. Proc. Natl. Acad. Sci. USA 2000, 97, 7030–7036. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.T.; Davis, K.E.; Pisani, D.; Tarver, J.E.; Ruta, M.; Sakamoto, M.; Hone, D.W.; Jennings, R.; Benton, M.J. Dinosaurs and the Cretaceous terrestrial revolution. Proc. R. Soc. B: Biol. Sci. 2008, 275, 2483–2490. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.W.; Oboh-Ikuenobe, F.E.; Benson, D.G., Jr.; Holbrook, J.M.; Alnahwi, A. Cenomanian-Turonian flooding cycles: US Gulf Coast and western interior. Cretac. Res. 2018, 89, 191–210. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; Decker, P.L.; LePain, D.L.; Wartes, M.; McCarthy, P.J. A probable neoceratopsian manus track from the Nanushuk Formation (Albian, northern Alaska). J. Iber. Geol. 2010, 36, 165–174. [Google Scholar] [CrossRef]
- Spicer, R.A.; Herman, A.B. The Albian–Cenomanian flora of the Kukpowruk River, western North Slope, Alaska: Stratigraphy, palaeofloristics, and plant communities. Cretac. Res. 2001, 22, 1–40. [Google Scholar] [CrossRef]
- Moore, T.E.; Wallace, W.K.; Bird, K.J.; Karl, S.M.; Mull, C.G.; Dillon, T.T. The Geology of North America, v. G-1, the Geology of Alaska; Plafker, G., Berg, H.C., Eds.; Geological Society of America: Boulder, CO, USA, 1994; pp. 49–140. [Google Scholar]
- Bird, K.J.; Molenaar, C.M. The North Slope Foreland Basin, Alaska: Chapter 13. Am. Assoc. Pet. Geol. Mem. 1992, 55, 363–393. [Google Scholar]
- Molenaar, C.M. Subsurface correlations and depositional history of the Nanushuk Group and related strata, North Slope, Alaska. In Geology of the Nanushuk Group and Related Rocks; U.S. Geological Survey Bulletin: North Slope, AK, USA, 1985; Volume 1614, pp. 37–59. [Google Scholar]
- Molenaar, C.M. Depositional history and seismic stratigraphy of Lower Cretaceous rocks in the National Petroleum Reserve in Alaska and adjacent areas. In U.S. Geological Survey Professional Paper; U.S. Geological Survey Bulletin: North Slope, AK, USA, 1988; Volume 1399, pp. 593–621. [Google Scholar]
- Finzel, E.S.; McCarthy, P.J. Architectural Analysis of Fluvial Conglomerate in the Nanushuk Formation, Brooks Range Foothills, Alaska; Alaska Division of Geological & Geophysical Surveys: Fairbanks, AK, USA, 2005; pp. 1–20.
- Fisher, W.I.; Brown, L.F., Jr.; Scott, A.J.; McGowen, J.H. DeltaSystems in the Exploration for Oil and Gas; Texas Bureau of Economic Geology: Austin, TX, USA, 1969; pp. 1–78. [Google Scholar]
- Huffman, A.C., Jr. Geology of the Nanushuk Group and Related Rocks; U.S. Geological Survey Bulletin: North Slope, AK, USA, 1985; Volume 1614, pp. 1–6.
- Huffman, A.C., Jr.; Ahlbrandt, T.S.; Bartsch-Winkler, S. Geology and exploration of the National Petroleum Reserve in Alaska, 1974 to 1982. In U.S. Geological Survey Professional Paper; U.S. Geological Survey Bulletin: North Slope, AK, USA, 1988; Volume 1399, pp. 281–298. [Google Scholar]
- Ahlbrandt, T.S.; Huffman, A.C.; Fox, J.E.; Pasternack, I. Depositional Framework and Reservoir-Quality Studies of Selected Nanushuk Group outcrops, North Slope, Alaska; U.S. Geological Survey Circular: North Slope, AK, USA, 1979; Volume 794, pp. 14–31.
- LePain, D.L.; McCarthy, P.J.; Kirkham, R. Sedimentology and sequence stratigraphy of the middle Albian–Cenomanian Nanushuk Formation in outcrop, central North Slope, Alaska. Alaska Division of Geological & Geophysical Surveys. Rep. Investig. 2009, 2009, 1–86. [Google Scholar]
- McCarthy, P.J. Alluvial Facies and Paleosols in the Cretaceous Nanushuk Formation, Kanayut River, North Slope, Alaska: Preliminary Results from the 2001 Field Season; Alaska Division of Geological & Geophysical Surveys: North Slope, AK, USA, 2003; Volume 1–19.
- Mull, C.G.; Houseknecht, D.W.; Bird, K.J. Revised Cretaceous and Tertiary stratigraphic nomenclature in the Colville Basin, northern Alaska. In U.S. Geological Survey Professional Paper; U.S. Geological Survey Bulletin: North Slope, AK, USA, 2003; Volume 1673, pp. 1–51. Available online: https://pubs.usgs.gov/pp/p1673/text.html (accessed on 5 May 2023).
- Detterman, R.L.; Bickel, R.S.; Gryc, G. Geology of the Chandler River region, Alaska. In U.S. Geological Survey Professional Paper; U.S. Geological Survey Bulletin: North Slope, AK, USA, 1963; Volume 303-E, pp. 233–324. [Google Scholar]
- Shimer, G.T.; McCarthy, P.J.; Hanks, C.L. Sedimentology, stratigraphy, and reservoir properties of an unconventional, shallow, frozen petroleum reservoir in the Cretaceous Nanushuk Formation at Umiat field, North Slope, Alaska. Am. Assoc. Pet. Geol. Bull. 2014, 98, 631–661. [Google Scholar] [CrossRef]
- Mull, C.G.; Harris, E.E.; Reifenstuhl, R.R.; Moore, T.E. Alaska Division of Geological & Geophysical Surveys Report of Investigations. 2000, 2000–2002, 1 sheet, scale 1:63,360. Available online: https://pubs.usgs.gov/pp/pp1662/biblio.pdf (accessed on 12 April 2023).
- LePain, D.L.; Decker, P.L. Lithofacies analysis of the Wainwright #1 continuous core, western Arctic Slope, Alaska: Transition from lower to upper delta plain environments in the Albian-Cenomanian Nanushuk Formation. In Alaska Division of Geological & Geophysical Surveys Report of Investigation; Alaska Division of Geological & Geophysical Surveys: North Slope, AK, USA, 2016; Volume 3, pp. 1–56. [Google Scholar] [CrossRef]
- Spicer, R.A. The Cretaceous World; Skelton, P., Ed.; Cambridge University Press: Cambridge, UK, 2003; pp. 85–162. [Google Scholar]
- Gehrels, G. Detrital Zircon U-Pb Geochronology Applied to Tectonics. Annu. Rev. Earth Planet. Sci. 2014, 42, 127–149. [Google Scholar] [CrossRef]
- Sundell, K.E.; Gehrels, G.E.; Pecha, M.E. Rapid U-Pb geochronology by laser ablation multi-collector ICP-MS. Geostand. Geoanal. Res. 2021, 45, 37–57. [Google Scholar] [CrossRef]
- Garza, H.K.; Catlos, E.J.; Chamberlain, K.R.; Suarez, S.E.; Brookfield, M.E.; Stockli, D.F.; Batchelor, R.A. How old is the Ordovician–Silurian boundary at Dob’s Linn, Scotland? Integrating LA-ICP-MS and CA-ID-TIMS U-Pb zircon dates. Geol. Mag. 2023, 160, 1775–1789. [Google Scholar] [CrossRef]
- Bowring, S.A.; Schmitz, M.D. High-precision U-Pb zircon geochronology and the stratigraphic record. Rev. Mineral. Geochem. 2003, 53, 305–326. [Google Scholar] [CrossRef]
- Tian, H.; Fan, M.; Valencia, V.A.; Chamberlain, K.; Waite, L.; Stern, R.J.; Loocke, M. Rapid early Permian tectonic reorganization of Laurentia’s plate margins: Evidence from volcanic tuffs in the Permian Basin, USA. Gondwana Res. 2022, 111, 76–94. [Google Scholar] [CrossRef]
- Coutts, D.S.; Matthews, W.A.; Hubbard, S.M. Assessment of widely used methods to derive depositional ages from detrital zircon populations. Geosci. Front. 2019, 10, 1421–1435. [Google Scholar] [CrossRef]
- Herriott, T.M.; Crowley, J.L.; Schmitz, M.D.; Wartes, M.A.; Gillis, R.J. Exploring the law of detrital zircon: LA-ICP-MS and CA-TIMS geochronology of Jurassic forearc strata, Cook Inlet, Alaska, USA. Geology 2019, 47, 1044–1048. [Google Scholar] [CrossRef]
- Trop, J.M.; Benowitz, J.; Cole, R.B.; O’Sullivan, P. Cretaceous to Miocene magmatism, sedimentation, and exhumation within the Alaska Range suture zone: A polyphase reactivated terrane boundary. Geosphere 2019, 15, 1066–1101. [Google Scholar] [CrossRef]
- Ludwig, K.R. Isoplot v. 3.75—a geochronological toolkit for Microsoft Excel. In Berkeley Geochronology Center Special Publication; Berkeley Geochronology Center: Berkeley, CA, USA, 2012; Volume 5, pp. 1–75. [Google Scholar]
- Smith, D.G. Meandering River Point Bar Lithofacies Models: Modern and Ancient Examples Compared; Ethridge, F.G., Flores, R.M., Harvey, M.D., Eds.; SEPM Special Publication: Tulsa, OK, USA, 1997; Volume 39, pp. 83–91. [Google Scholar]
- McCarthy, P.J.; Martini, I.P.; Leckie, D.A. Anatomy and evolution of a Lower Cretaceous alluvial plain: Sedimentology and palaeosols in the upper Blairmore Group, south-western Alberta, Canada. Sedimentology 1997, 44, 197–220. [Google Scholar] [CrossRef]
- Coleman, J.M. Ecological changes in a massive fresh-water clay sequence. Trans. Gulf Coast Assoc. Geol. Soc. 1966, 16, 159–174. [Google Scholar] [CrossRef]
- Perez-Arlucea, M.; Smith, N.D. Abstract: Proceedings of the 18th IAS Regional European Meeting of Sedimentology, Heidelberg, Germany, 2–4 September 1997; IAS: Heidelberg, Germany, 1997; Volume 204, pp. 269–270. [Google Scholar]
- Bridge, J.S. Large-scale facies sequences in alluvial overbank environments. J. Sediment. Petrol. 1984, 54, 583–588. [Google Scholar]
- Ten Brinke, W.B.M.; Schoor, M.R.; Sorber, A.M.; Berendsen, H.J.A. Overbank sand deposition in relation to transport volumes during large-magnitude floods in the Dutch sand-bed Rhine river system. Earth Surf. Process. Landf. 1998, 23, 809–824. [Google Scholar] [CrossRef]
- Smith, N.D.; Perez-Arlucea, M. Fine-grained splay deposition in the avulsion belt of the lower Saskatchewan River, Canada. J. Sediment. Res. 1994, B64, 159–168. [Google Scholar]
- Smith, D.G.; Smith, N.D. Sedimentation in anastomosed river systems; examples from alluvial valleys near Banff, Alberta. J. Sediment. Petrol. 1980, 50, 157–164. [Google Scholar] [CrossRef]
- Retallack, G.J. Soils of the Past: An Introduction to Paleopedology, 2nd ed.; Blackwell Scientific Publishers: Oxford, UK, 2001. [Google Scholar]
- McCarthy, P.J.; Martini, I.P.; Leckie, D.A. Use of micromorphology for palaeoenvironmental interpretation of complex alluvial palaeosols: An example from the Mill Creek Formation (Albian), southwestern Alberta, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 143, 87–110. [Google Scholar] [CrossRef]
- Kraus, M.J. Paleosols in clastic sedimentary rocks: Their geologic applications. Earth Sci. Rev. 1999, 47, 41–70. [Google Scholar] [CrossRef]
- McCarthy, P.J. Micromorphology and development of interfluve paleosols: A case study from the Cenomanian Dunvegan Formation, NE British Columbia, Canada. Bull. Can. Pet. Geol. 2002, 50, 158–177. [Google Scholar] [CrossRef]
- Spicer, R.A.; Parrish, J.T.; Grant, P.R. Controls on the Distribution and Quality of Cretaceous Coals. In Geological Society of America Special Paper; McCabe, P.J., Parrish, J.T., Eds.; Geological Society of America: Boulder, CO, USA, 1992; Volume 267, pp. 177–192. [Google Scholar]
- Fiorillo, A.R. Alaska Dinosaurs: An Ancient Arctic World; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Alexander, R. McN. Estimates of speeds of dinosaurs. Nature 1976, 261, 129–130. [Google Scholar] [CrossRef]
- Henderson, D. Footprints, trackways, and hip heights of bipedal dinosaurs—testing hip height predictions with computer models. Ichnos 2003, 10, 99–114. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; Tykoski, R.S. Small hadrosaur manus and pes tracks from the Lower Cantwell Formation (Upper Cretaceous) Denali National Park, Alaska: Implications for locomotion in juvenile hadrosaurs. Palaios 2016, 31, 479–482. [Google Scholar] [CrossRef]
- Seebacher, F. A new method to calculate allometric length-mass relationships of dinosaurs. J. Vertebr. Paleontol. 2001, 21, 51–60. [Google Scholar] [CrossRef]
- Nesbitt, S.J.; Denton, R.K., Jr.; Loewen, M.A.; Brusatte, S.L.; Smith, N.D.; Turner, A.H.; Kirkland, J.I.; McDonald, A.T.; Wolfe, D.G. A mid-Cretaceous tyrannosauroid and the origin of North American end-Cretaceous dinosaur assemblages. Nat. Ecol. Evol. 2019, 3, 892–899. [Google Scholar] [CrossRef]
- Brusatte, S.L.; Averianov, A.; Sues, H.D.; Muir, A.; Butler, I.B. New tyrannosaur from the mid-Cretaceous of Uzbekistan clarifies evolution of giant body sizes and advanced senses in tyrant dinosaurs. Proc. Natl. Acad. Sci. USA 2016, 113, 3447–3452. [Google Scholar] [CrossRef]
- Rezendes, P. Tracking and the Art of Seeing: How to Read Animal Tracks and Sign, 2nd ed.; Harper Collins Publishers: New York, NY, USA, 1999. [Google Scholar]
- Elbroch, M.; Marks, E. Bird Tracks and Sign; Stackpole Books: Mechanicsburg, PA, USA, 2001. [Google Scholar]
- Currie, P.J. Bird footprints from the Gething Formation (Aptian, Lower Cretaceous) of northeastern British Columbia, Canada. J. Vertebr. Paleontol. 1981, 1, 257–264. [Google Scholar] [CrossRef]
- Mehl, M.G. Additions to the vertebrate record of the Dakota Sandstone. Am. J. Sci. 1931, 21, 441–452. [Google Scholar] [CrossRef]
- Lockley, M.; Chin, K.; Houck, K.; Matsukawa, M.; Kukihara, R. New interpretations of Ignotornis, the first-reported Mesozoic avian footprints: Implications for the paleoecology and behavior of an enigmatic Cretaceous bird. Cretac. Res. 2009, 30, 1041–1061. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; Hasiotis, S.T.; Kobayashi, Y.; Breithaupt, B.H.; McCarthy, P.J. Bird tracks from the Upper Cretaceous Cantwell Formation of Denali National Park, Alaska, USA: A new perspective on ancient northern polar vertebrate biodiversity. J. Syst. Palaeontol. 2011, 9, 33–49. [Google Scholar] [CrossRef]
- Kang, S.H.; Buckley, L.G.; McCrea, R.T.; Kim, K.S.; Lockley, M.G.; Lim, J.D.; Lim, H.S.; Kim, C.B. First report of bird tracks (Ignotornis seoungjoseoi ichnosp. nov.) from the Jinju Formation (Lower Cretaceous), Sacheon City, Korea. Cretac. Res. 2021, 127, 104899. [Google Scholar] [CrossRef]
- Currie, P.; Badamgarav, D.; Koppelhus, E. The first Late Cretaceous footprints from the Nemegt locality in the Gobi of Mongolia. Ichnos 2003, 10, 1–13. [Google Scholar] [CrossRef]
- Lockley, M.G.; Nadon, G.; Currie, P.J. A diverse dinosaur-bird footprint assemblage from the Lance Formation, Upper Cretaceous, eastern Wyoming: Implications for ichnotaxonomy. Ichnos 2004, 11, 229–249. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; Hasiotis, S.T.; Kobayashi, Y. Herd structure in Late Cretaceous polar dinosaurs: A remarkable new dinosaur tracksite, Denali National Park, Alaska, USA. Geology 2014, 42, 719–722. [Google Scholar] [CrossRef]
- Diaz-Martinez, I.; Pereda-Suberbiola, X.; Perez-Lorente, F.; Canudo, J.I. Ichnotaxonomic review of large ornithopod dinosaur tracks: Temporal and geographic implications. PLoS ONE 2015, 10, e0115477. [Google Scholar] [CrossRef] [PubMed]
- Fiorillo, A.R.; Parrish, J.T. The first record of a Cretaceous dinosaur from southwestern Alaska. Cretac. Res. 2004, 25, 453–458. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; Adams, T.L.; Kobayashi, Y. New sedimentological, palaeobotanical, and dinosaur ichnological data on the palaeoecology of an unnamed Late Cretaceous rock unit in Wrangell-St. Elias National Park and Preserve, Alaska, USA. Cretac. Res. 2012, 37, 291–299. [Google Scholar] [CrossRef]
- Fanti, F.; Bell, P.R.; Sissons, R.L. A diverse, high-latitude ichnofauna from the Late Cretaceous Wapiti Formation, Alberta, Canada. Cretac. Res. 2013, 41, 256–269. [Google Scholar] [CrossRef]
- Lee, Y.N. Bird and dinosaur footprints in the Woodbine Formation (Cenomanian), Texas. Cretac. Res. 1997, 18, 849–864. [Google Scholar] [CrossRef]
- Xing, L.; Lockley, M.G.; Kim, K.S.; Klein, H.; Matsukawa, M.; Lim, J.D.; Persons, W.S.; Xu, X. Mid-Cretaceous dinosaur track assemblage from the Tongfosi Formation of China: Comparison with the track assemblage of South Korea. Cretac. Res. 2017, 74, 155–164. [Google Scholar] [CrossRef]
- McCrea, R.T.; Lockley, M.G.; Meyer, C.A. The Armored Dinosaurs; Carpenter, K., Ed.; Indiana University Press: Bloomington, IN, USA, 2001; pp. 413–454. [Google Scholar]
- Riguetti, F.; Citton, P.; Apesteguía, S.; Zacarías, G.G.; Pereda-Suberbiola, X. New ankylosaurian trackways (cf. Tetrapodosaurus) from an uppermost Cretaceous level of the El Molino Formation of Bolivia. Cretac. Res. 2021, 124, 104810. [Google Scholar] [CrossRef]
- Gangloff, R.A. Edmontonia sp., the first record of an ankylosaur from Alaska. J. Vertebr. Paleontol. 1995, 15, 195–200. [Google Scholar] [CrossRef]
- Godefroit, P.; Golovneva, L.; Shchepetov, S.; Garcia, G.; Alekseev, P. The last polar dinosaurs: High diversity of latest Cretaceous arctic dinosaurs in Russia. Naturwissenschaften 2009, 96, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Kohn, M.J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo) ecology and (paleo) climate. Proc. Natl. Acad. Sci. USA 2010, 107, 19691–19695. [Google Scholar] [CrossRef] [PubMed]
- Diefendorf, A.F.; Mueller, K.E.; Wing, S.L.; Koch, P.L.; Freeman, K.H. Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proc. Natl. Acad. Sci. USA 2010, 107, 5738–5743. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Long, A. Stable-carbon isotope variability in tree foliage and wood. Ecology 1986, 67, 1002–1010. [Google Scholar] [CrossRef]
- Ciesielczuk, J.; Górka, M.; Fabiańska, M.J.; Misz-Kennan, M.; Jura, D. The influence of heating on the carbon isotope composition, organic geochemistry and petrology of coal from the Upper Silesian Coal Basin (Poland): An experimental and field study. Int. J. Coal Geol. 2021, 241, 103749. [Google Scholar] [CrossRef]
- Barral, A.; Gomez, B.; Legendre, S.; Lécuyer, C. Evolution of the carbon isotope composition of atmospheric CO2 throughout the Cretaceous. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 471, 40–47. [Google Scholar] [CrossRef]
- Poulsen, C.J.; Pollard, D.; White, T.S. General circulation model simulation of the δ18O content of continental precipitation in the middle Cretaceous: A model-proxy comparison. Geology 2007, 35, 199–202. [Google Scholar] [CrossRef]
- Suarez, M.B.; González, L.A.; Ludvigson, G.A. Quantification of a greenhouse hydrologic cycle from equatorial to polar latitudes: The mid-Cretaceous water bearer revisited. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 307, 301–312. [Google Scholar] [CrossRef]
- Gangloff, R.A. Arctic dinosaurs with emphasis on the Cretaceous record of Alaska and the Eurasian-North American connection. New Mex. Mus. Nat. Hist. Sci. Bull. 1998, 14, 211–220. [Google Scholar]
- Roehler, H.W.; Stricker, G.D. Dinosaur and wood fossils from the Cretaceous Corwin Formation in the National Petroleum Reserve, north slope of Alaska. J. Alsk. Geol. Soc. 1984, 4, 35–41. [Google Scholar]
- Parrish, J.M.; Parrish, J.T.; Hutchison, J.H.; Spicer, R.A. Late Cretaceous vertebrate fossils from the North Slope of Alaska and implications for dinosaur ecology. Palaios 1987, 2, 377–389. [Google Scholar] [CrossRef]
- Lease, R.O.; Houseknecht, D.W.; Kylander-Clark, A.R. Quantifying large-scale continental shelf margin growth and dynamics across middle-Cretaceous Arctic Alaska with detrital zircon U-Pb dating. Geology 2022, 50, 620–625. [Google Scholar] [CrossRef]
- Fiorillo, A.R.; Kobayashi, Y.; McCarthy, P.J.; Tanaka, T.; Tykoski, R.S.; Lee, Y.N.; Takasaki, R.; Yoshida, J. Dinosaur ichnology and sedimentology of the Chignik Formation (Upper Cretaceous), Aniakchak National Monument, southwestern Alaska; Further insights on habitat preferences of high-latitude hadrosaurs. PLoS ONE 2019, 14, e0223471. [Google Scholar] [CrossRef] [PubMed]
- McCrea, R.T.; Buckley, L.G.; Currie, P.J.; Plint, A.G.; Helm, C.W.; Haggart, J.W. A review of vertebrate track-bearing formations from the Mesozoic and earliest Cenozoic of western Canada with a description of a new theropod ichnospecies and reassignment of an avian ichnogenus. New Mex. Mus. Nat. Hist. Sci. Bull. 2014, 62, 5–94. [Google Scholar]
- Bamforth, E. Exploring the Role of Evolving Forest Composition in Shaping Dinosaur Diversity Patterns in the Cretaceous of Northern Alberta, Canada. In Proceedings of the Society of Vertebrate Paleontology Annual Meeting, Cincinnati, OH, USA, 18–21 October 2023; pp. 81–82. [Google Scholar]
- Bart, J.R.; Johnston, V.H. Arctic shorebirds in North America: A Decade of Monitoring; University of California Press: Berkeley, CA, USA, 2012. [Google Scholar]
- Lawver, L.A.; Grantz, A.; Gahagan, L.M. Tectonic Evolution of the Bering Shelf–Chukchi Sea–Arctic Margin and Adjacent Landmasses. In Geological Society of America Special Paper; Miller, E.L., Grantz, A., Klemperer, S.L., Eds.; Geological Society of America: Boulder, CO, USA, 2002; Volume 360, pp. 333–358. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ankylosaur Skeletal | Ceratopsian Skeletal |
|---|---|
|
|
| Ankylosaur Footprint | Ceratopsian Footprint |
(Tetrapodosaurus ichnosp.)
| (Ceratopsipes ichnosp.)
|
| Sample Name | C ‰ vs. VPDB | Cleaf‰ vs. VPDB | Δleaf | Mean Annual Precipitation (mm/year) |
|---|---|---|---|---|
| KUKP-1 | −26.6 | −27.7 | 22.2 | 1901 |
| KUKP-3 | −27.0 | −28.0 | 22.7 | 2284 |
| KUKP-4 | −25.8 | −26.8 | 21.4 | 1334 |
| KUKP-5 | −26.1 | −27.1 | 21.8 | 1562 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiorillo, A.R.; McCarthy, P.J.; Shimer, G.; Suarez, M.B.; Takasaki, R.; Chinzorig, T.; Kobayashi, Y.; O’Sullivan, P.; Orphys, E. New Dinosaur Ichnological, Sedimentological, and Geochemical Data from a Cretaceous High-Latitude Terrestrial Greenhouse Ecosystem, Nanushuk Formation, North Slope, Alaska. Geosciences 2024, 14, 36. https://doi.org/10.3390/geosciences14020036
Fiorillo AR, McCarthy PJ, Shimer G, Suarez MB, Takasaki R, Chinzorig T, Kobayashi Y, O’Sullivan P, Orphys E. New Dinosaur Ichnological, Sedimentological, and Geochemical Data from a Cretaceous High-Latitude Terrestrial Greenhouse Ecosystem, Nanushuk Formation, North Slope, Alaska. Geosciences. 2024; 14(2):36. https://doi.org/10.3390/geosciences14020036
Chicago/Turabian StyleFiorillo, Anthony R., Paul J. McCarthy, Grant Shimer, Marina B. Suarez, Ryuji Takasaki, Tsogtbaatar Chinzorig, Yoshitsugu Kobayashi, Paul O’Sullivan, and Eric Orphys. 2024. "New Dinosaur Ichnological, Sedimentological, and Geochemical Data from a Cretaceous High-Latitude Terrestrial Greenhouse Ecosystem, Nanushuk Formation, North Slope, Alaska" Geosciences 14, no. 2: 36. https://doi.org/10.3390/geosciences14020036
APA StyleFiorillo, A. R., McCarthy, P. J., Shimer, G., Suarez, M. B., Takasaki, R., Chinzorig, T., Kobayashi, Y., O’Sullivan, P., & Orphys, E. (2024). New Dinosaur Ichnological, Sedimentological, and Geochemical Data from a Cretaceous High-Latitude Terrestrial Greenhouse Ecosystem, Nanushuk Formation, North Slope, Alaska. Geosciences, 14(2), 36. https://doi.org/10.3390/geosciences14020036