

Vegetation, Climate and Habitability in the Marseille Basin (SE France) circa 1 Ma
 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
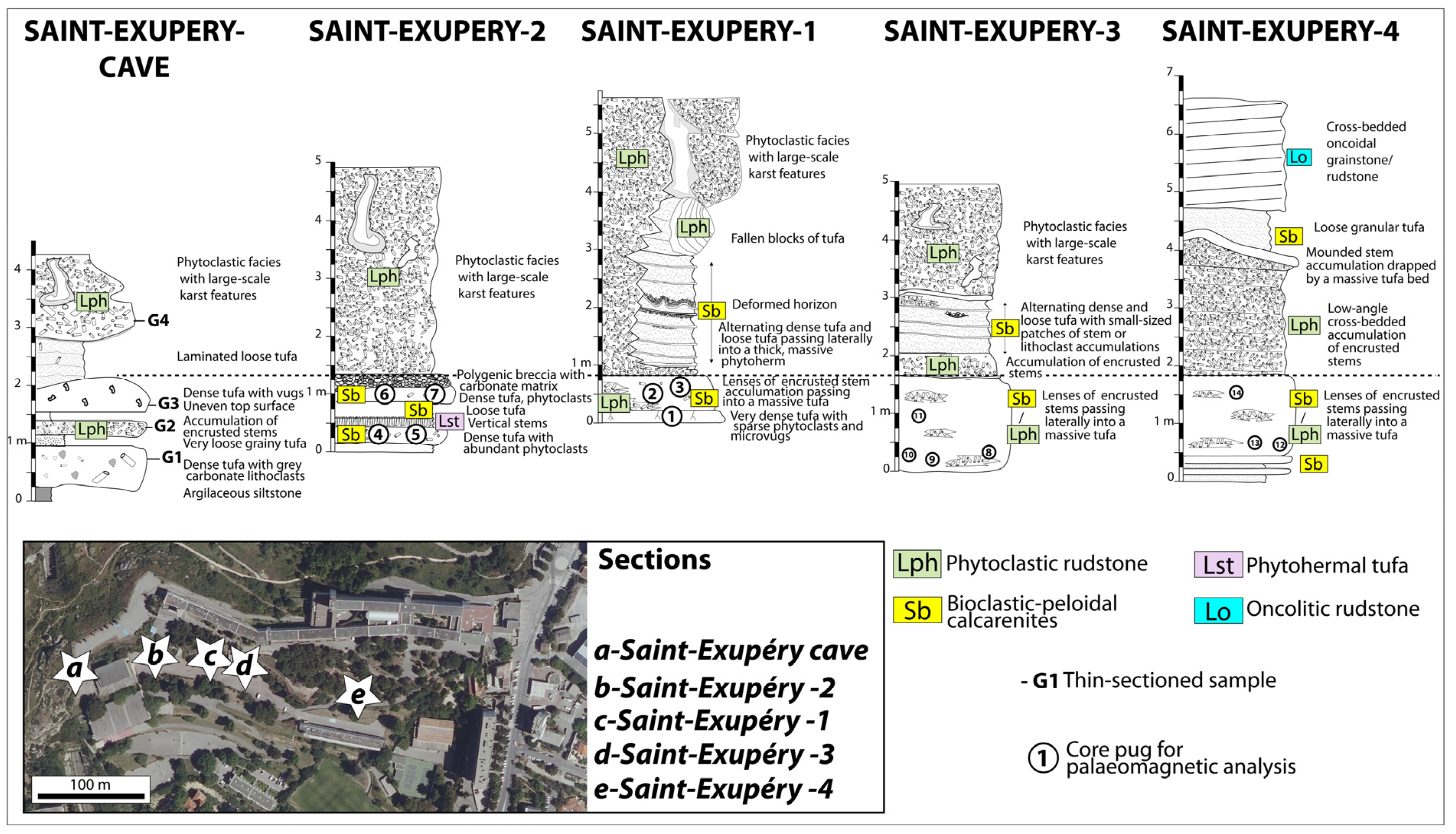
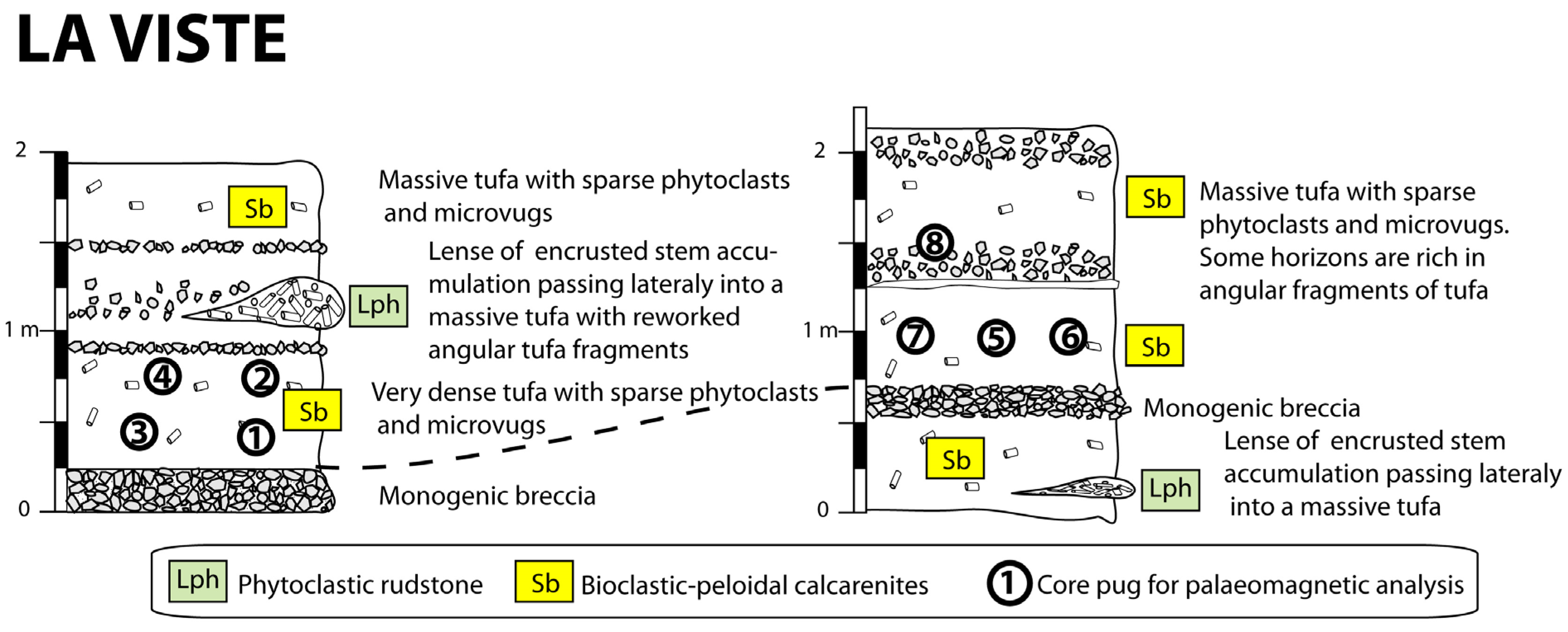
2. The Studied Sites and Sampling
3. Materials and Methods
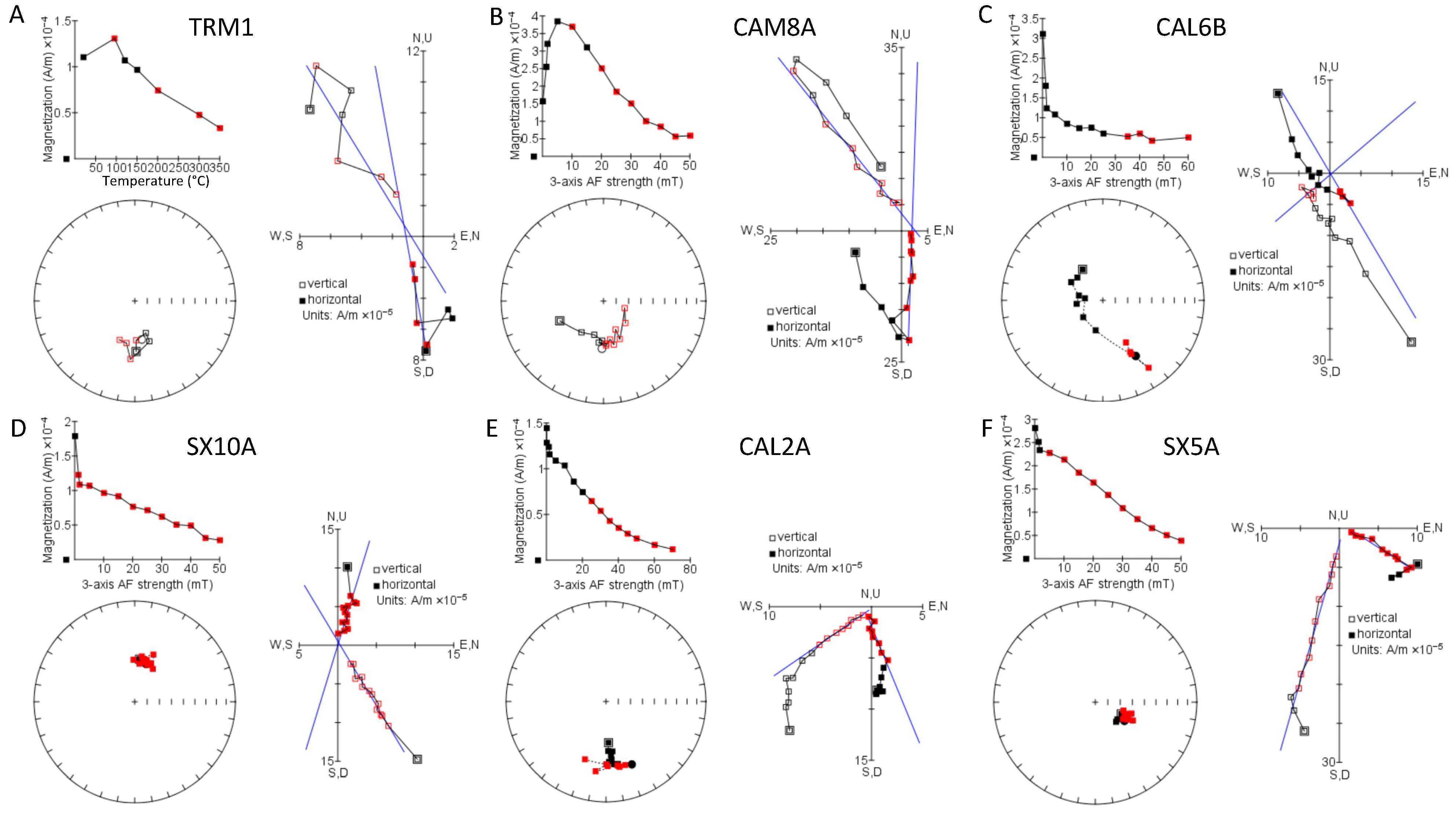
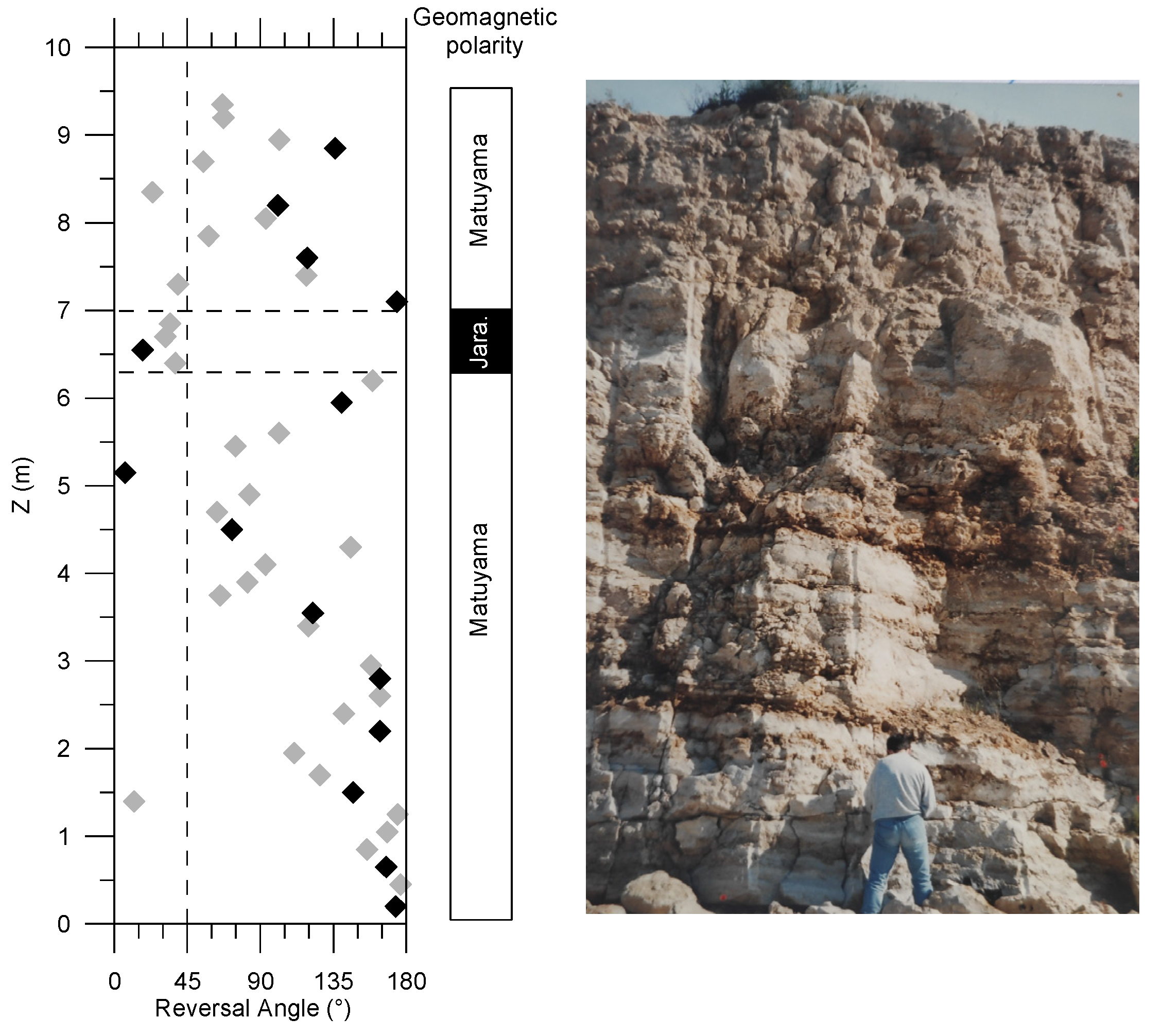
3.1. Palaeomagnetism
3.2. Sedimentologic and Petrographic Analyses
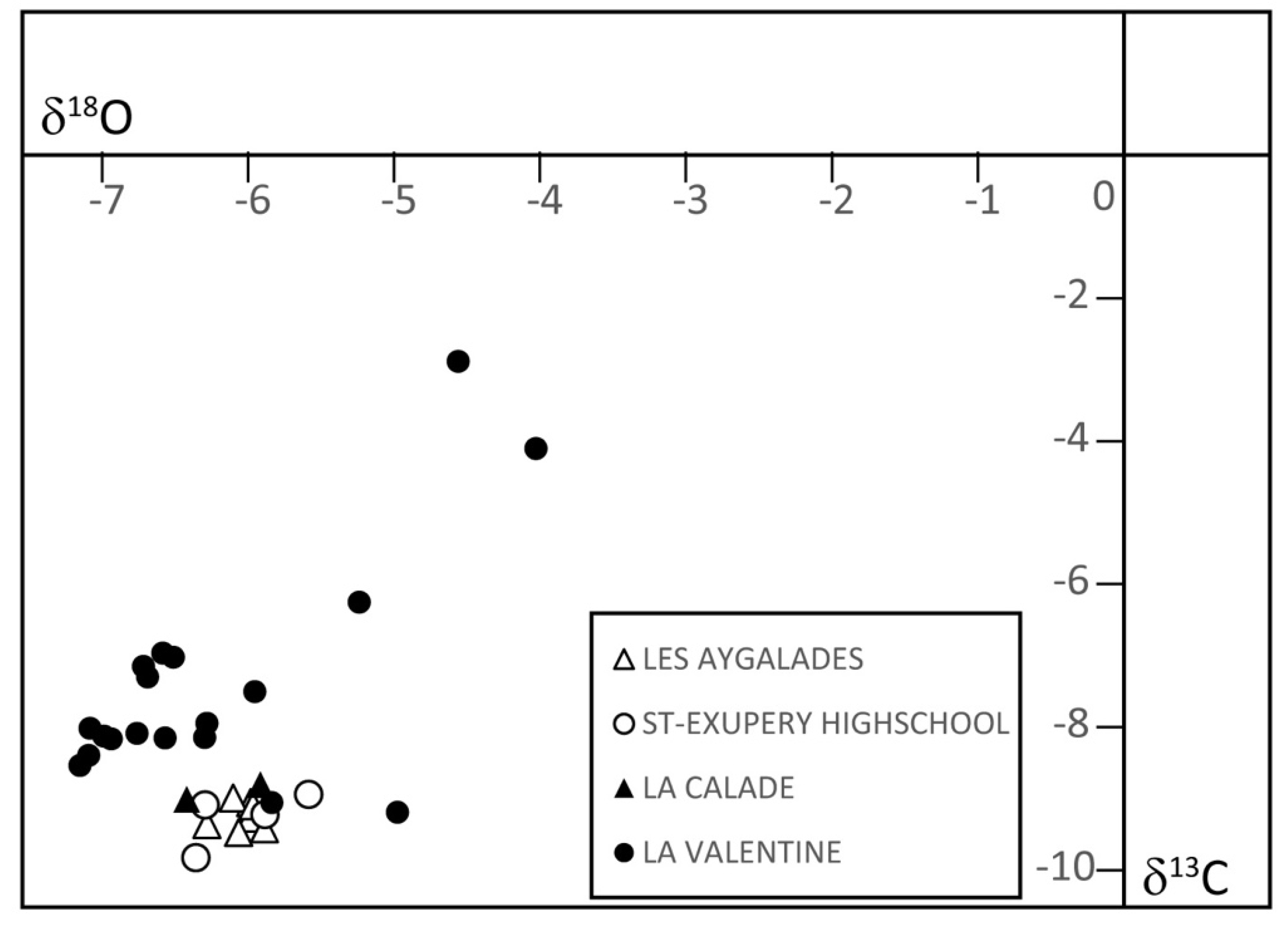
3.3. Carbon and Oxygen Stable Isotope Composition of Carbonates
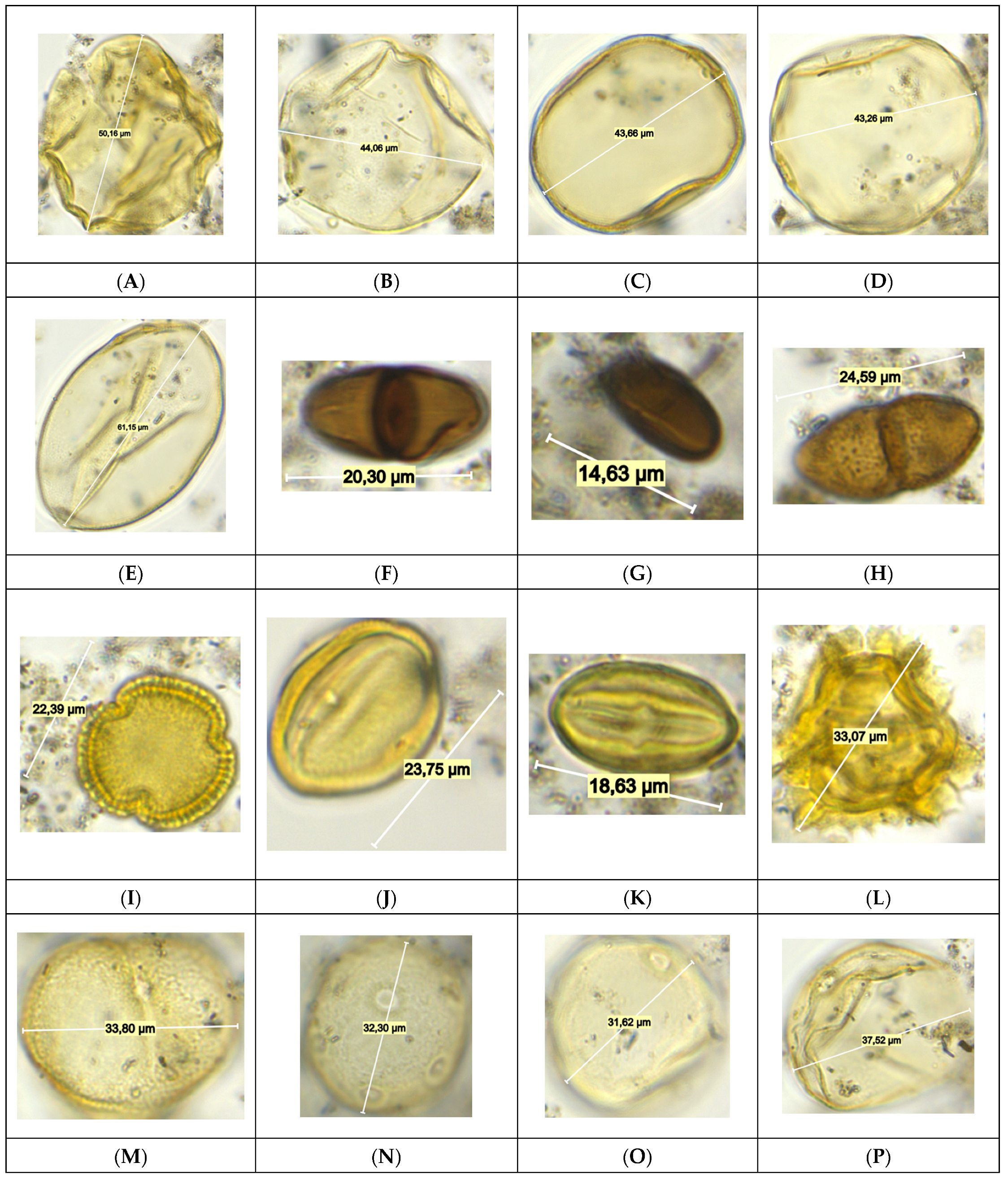
3.4. Pollen and Non-Pollen Palynomorphs (NPP) Analyses
3.5. Pollen-Inferred Climate Reconstructions
4. Results
4.1. Magnetostratigraphy
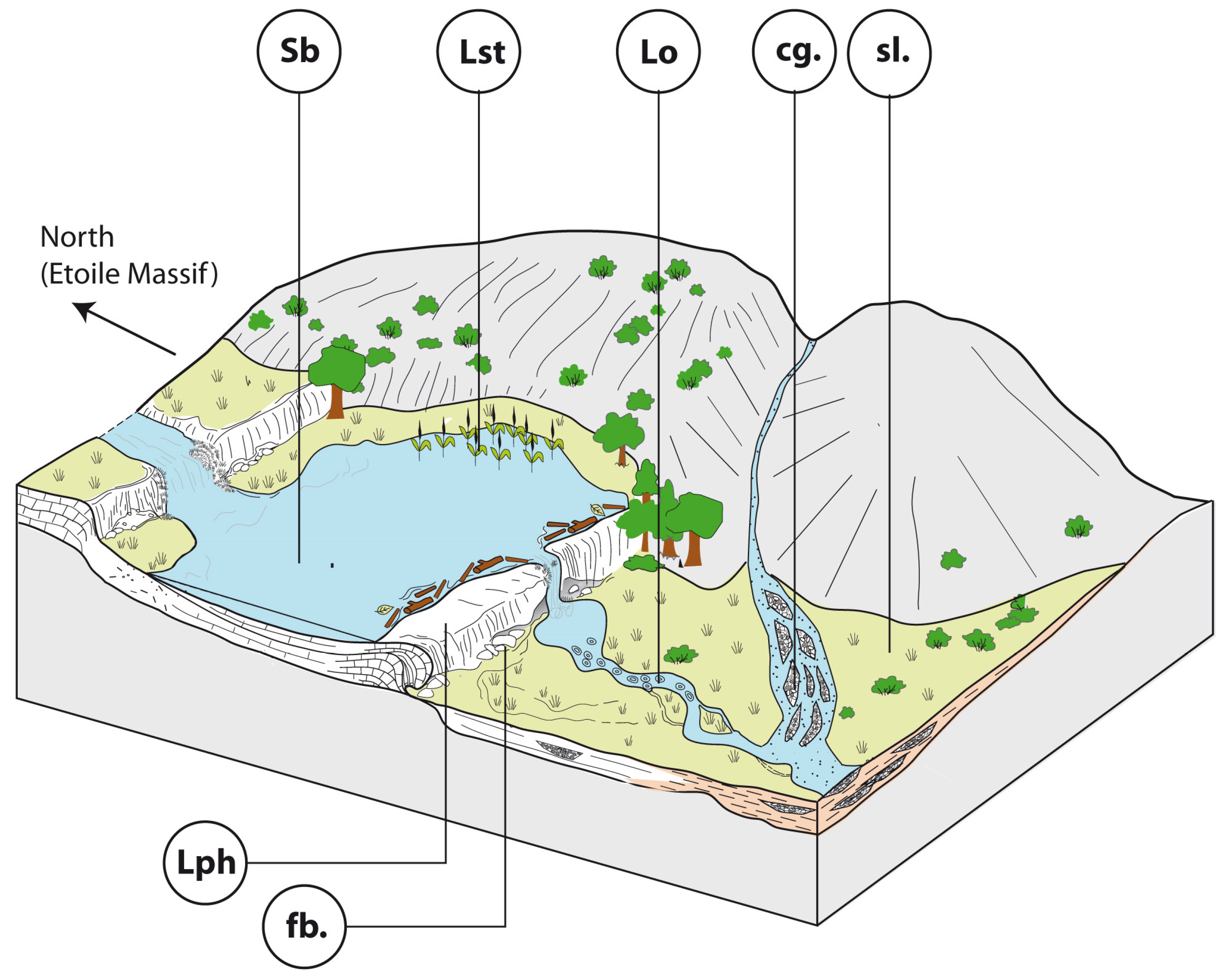
4.2. Depositional Facies of Calcareous Tufa and Palaeoenvironmental Reconstructions
- -
- Phytoclastic rudstones (Lph) consist of an accumulation of coated stems (centimetre- to decimetre-long) with a scarce peloidal micrite matrix. Phytoclastic rudstone form up to 4 m high domes, sometimes with subvertical walls (Figure 8B). They develop through calcite precipitation around plants. These plants may be transported before encrustation, thus forming a dam. Plant accumulations are believed to initiate dams around obstructions or slope breaks. Phytoclastic rudstones (Lph1) commonly pass laterally to calcarenites (Sb) (Figure 8B).
- -
- Bioclastic-peloidal calcarenites (Sb) are fine-to-medium grained carbonate sands. Grains predominantly consist of peloids, intraclasts (broken calcitic crusts), micro-oncoids and bioclasts (dominantly ostracods, more occasionally molluscs). They are organised into centimetric to decimetric thick beds with horizontal laminations (Figure 8B,C). They are interpreted as forming in slow-flowing dammed areas.
- -
- Photothermal tufa (Lst) are boundstones of in situ stems (mainly reeds) growing upward. Stems are commonly decimetre-long and exhibit thick (up to few centimetres) calcite coatings (Figure 8A). They form in palustrine settings on fluvial banks or lake shores.
- -
- Oncolitic rudstone (Lo) are formed by an accumulation of oncoids (typically less than 1 cm in diameter). They are organised into decimetre-thick lenses displaying an erosive base (Figure 8C) and commonly exhibit cross-stratification. They are interpreted as representing fluvial channel fills.
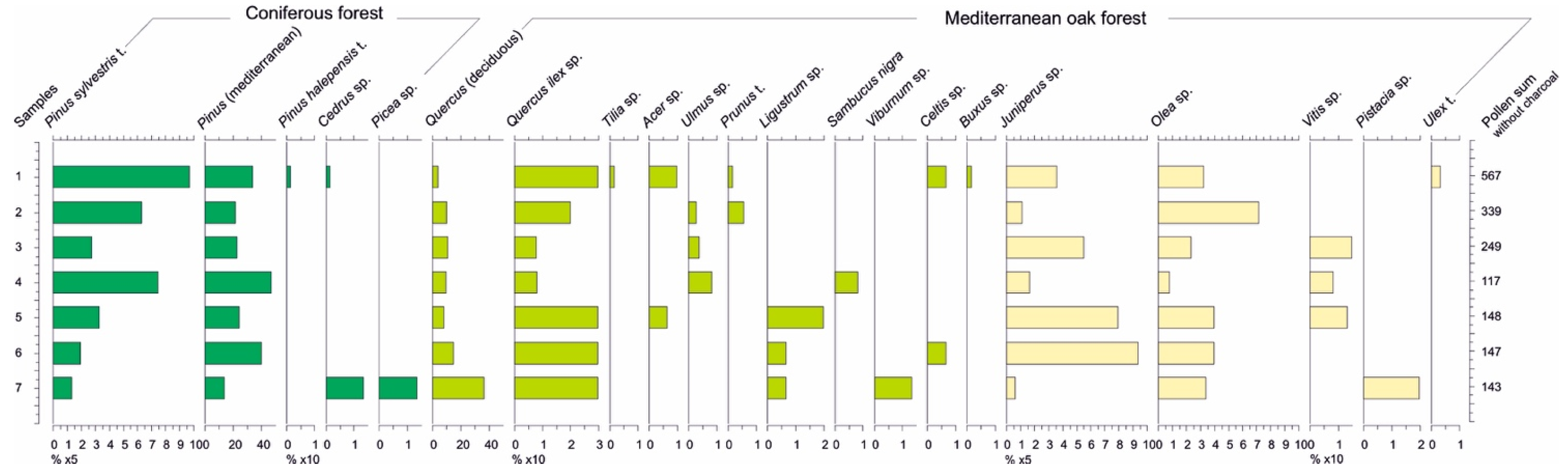
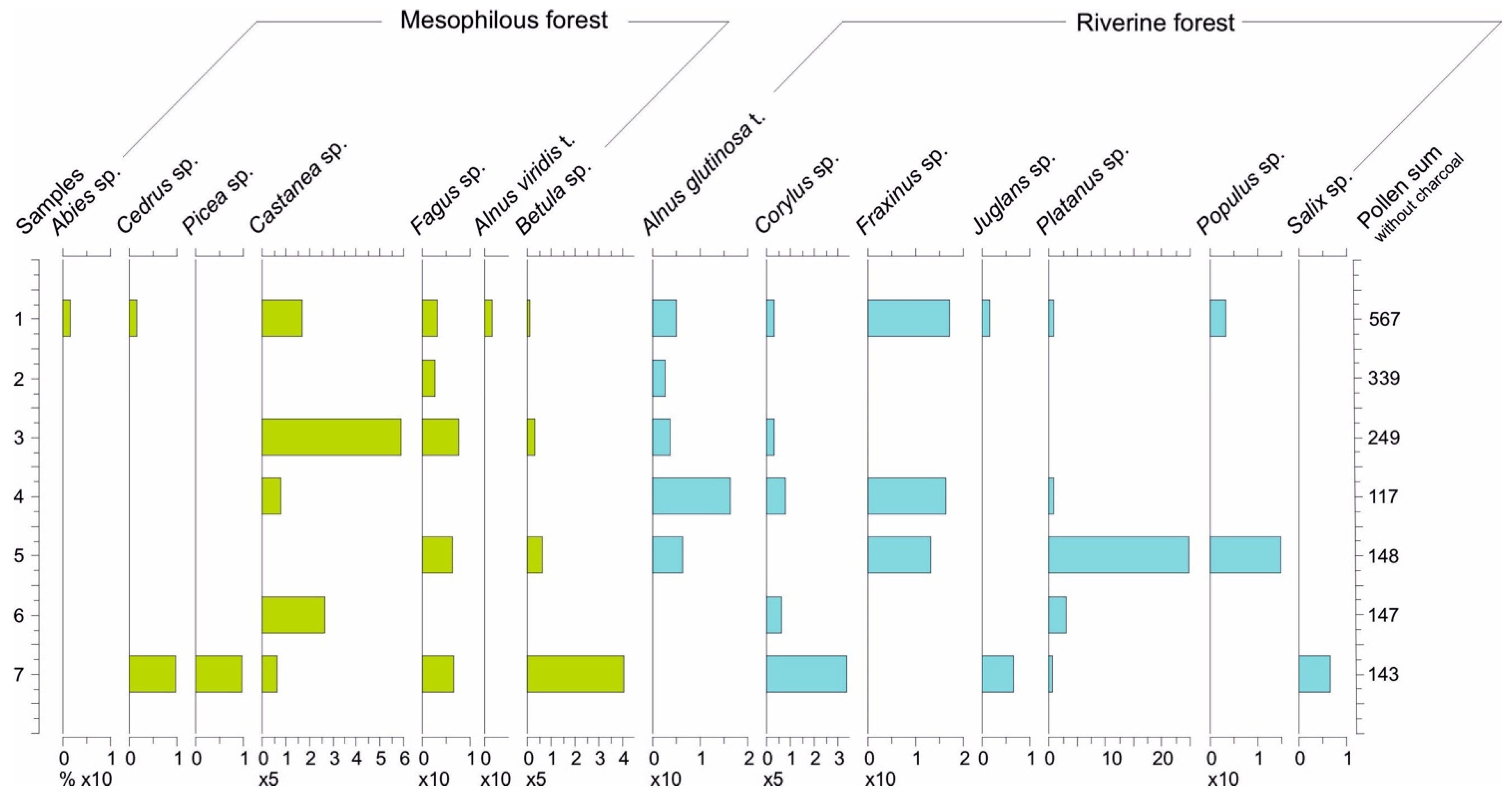
4.3. Pollen and NPP Data
4.4. Pollen-Inferred Climate Reconstructions
5. Discussion
5.1. A Diversified Geomorphological and Hydrological Landscape
5.2. A mosaic of Mediterranean Vegetation
5.3. The Early Appearance of Proto-Cereals
5.4. Potential Food Resources
5.5. A Cooler and Wetter Climate
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asmerom, Y.; Polyak, V.J.; Wagner, J.D.M.; Patchett, P.J. Hominin expansion into Central Asia during the last interglacial. Earth Planet. Sci. Lett. 2018, 494, 148–152. [Google Scholar] [CrossRef]
- Barboni, B.; Ashley, G.; Bourel, B.; Arraiz, H.; Mazur, J.C. Springs, Palm groves, and the record of early hominins in Africa. Rev. Palaeobot. Palynol. 2019, 266, 23–41. [Google Scholar] [CrossRef]
- Lebatard, A.E.; Alcicek, M.C.; Rochette, P.; Khatib, S.; Vialet, A.; Boulbes, N.; Bourlès, D.L.; Demory, F.; Guipert, G.; Mayda, S.; et al. Dating the Homo erectus bearing travertine from Kocabaş (Denizli, Turkey) at least 1.1 Ma. Earth Planet. Sci. Lett. 2014, 390, 8–18. [Google Scholar] [CrossRef]
- Rausch, L.; Alçiçek, H.; Vialet, A.; Boulbes, N.; Mayda, S.; Titov, V.V.; Stoica, M.; Charbonnier, S.; Abels, H.A.; Tesakov, A.S.; et al. An integrated reconstruction of the Early Pleistocene palaeoenvironment of Homo erectus in the Denizli Basin (SW Turkey). Geobios 2019, 57, 77–95. [Google Scholar] [CrossRef]
- Andrieu-Ponel, V.; Rochette, P.; Demory, F.; Alcicek, H.; Boulbes, N.; Bourlès, N.; Helvaci, C.; Lebatard, A.; Mayda, S.; Moigne, A.M.; et al. Continuous presence of proto-cereals in Anatolia since 2.3 Ma, and their possible co-evolution with large herbivores and hominins. Sci. Rep. (Nat.) 2021, 11, 8914. [Google Scholar] [CrossRef] [PubMed]
- Dupire, S. Etude Cartographique à 1/ 25.000 de la Zone Sud du Basin de Marseille. Les Travertins de Marseille: Aperçu Géomorphologique et Néotectonique. Ph.D. Thesis, Université de Provence, Marseille, France, 1985; p. 105. [Google Scholar]
- Toro Moyano, I.; Martinez Navarro, B.; Agusti, J.; Souday, C.; Bermudez de Castro, J.M.; Martinon Torres, M.; Fajardo, B.; Duval, M.; Falgueres, C.; Oms, O.; et al. The oldest human fossil in Europe, from Orce (Spain). J. Hum. Evol. 2013, 65, 1–9. [Google Scholar] [CrossRef]
- Alvarez, C.; Pares, J.M.; Granger, D.; Duval, M.; Sala, R.; Toro, I. New magnetostratigraphic and numerical age of the Fuente Nueva-3 site (Guadix-Baza basin, Spain). Quat. Int. 2015, 389, 224–234. [Google Scholar] [CrossRef]
- Carbonell, E.; Bermudez de Castro, J.M.; Pares, J.M.; Perez Gonzalez, A.; Cuenca Bescos, G.; Olle, A.; Mosquera, M.; Huguet, R.; van der Made, J.; Rosas, A.; et al. The first hominin of Europe. Nature 2008, 452, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Huguet, R.; Vallverdú, J.; Rodríguez Álvarez, X.P.; Terradillos Bernal, M.; Bargalló, A.; Lombera Hermida, A.; Menéndez, L.; Modesto Mata, M.; Van der Made, J.; Soto, M.; et al. Level TE9c of Sima del Elefante (Sierra de Atapuerca, Spain): A comprehensive approach. Quat. Int. 2017, 433, 278–295. [Google Scholar] [CrossRef]
- Michel, V.; Shen, C.C.; Woodhead, J.; Hu, H.M.; Wu, C.C.; Moullé, P.E.; Khatib, S.; Cauche, D.; Moncel, M.H.; Valensi, P.; et al. New dating evidence of the early presence of hominins in Southern Europe. Sci. Rep. (Nat.) 2017, 7, 10074. [Google Scholar] [CrossRef]
- Bourguignon, L.; Crochet, J.Y.; Capdevila, R.; Ivorra, J.; Antoine, P.O.; Agustí, J.; Barsky, D.; Blain, H.A.; Boulbes, N.; Bruxelles, L.; et al. Bois-de-Riquet (Lézignan-la-Cèbe, Hérault): A late Early Pleistocene archaeological occurrence in southern France. Quat. Int. 2016, 393, 24–40. [Google Scholar] [CrossRef]
- de Saporta, G. Sur la flore des tufs quaternaires en Provence. In Extrait des Comptes-Rendus de la 33ème Session du Congrès Scientifique de France; Typographie Remontet-Aubin: Aix-en-Provence, France, 1867; pp. 3–32. [Google Scholar]
- Pons, A. Les macroflores quaternaires de France. In Etudes Françaises sur le Quaternaire; VIIIth INQUA Congress: Paris, France, 1969; pp. 85–93. [Google Scholar]
- Médail, F. Mise en place de la flore: Approche biogéographique. In La Flore Remarquable des Bouches-du-Rhône; Pirès, M., Pavon, D., Eds.; Plantes, milieux naturels, paysages; Biotope: Mèze, France, 2018; pp. 22–50. [Google Scholar]
- Bonifay, E. La tectonique récente du bassin de Marseille dans le cadre de l’évolution post-Miocène du littoral méditerranéen français. Bull. Soc. Gél. France 1967, IX, 549–560. [Google Scholar]
- Villeneuve, M. (Ed.) Mémoire Explicatif, 3rd ed.; Carte géol. France (1/50,000), feuille Aubagne-Marseille; BRGM: Orléans, France, 2018; 333p. [Google Scholar]
- Bertini, A.; Minissale, A.; Ricci, M. Palynological approach in upper Quaternary terrestrial carbonates of central Italy: Anything but a ‘mission impossible’. Sedimentology 2014, 61, 200–220. [Google Scholar] [CrossRef]
- Ali, A.A.; Terral, J.F.; Girard, V.; Roiron, P. Les travertins à empreintes, témoins de la paléobiodiversité végétale. Quaternaire 2014, 25, 157–161. [Google Scholar] [CrossRef]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Rousselière, F. Les Proboscidiens du basin de Marseille. Bull. Du Musée Anthropol. Et De Préhistoire De Monaco 2004, 44, 39–48. [Google Scholar]
- Muttoni, G.; Scardia, G.; Kent, D.V. Early hominins in Europe: The Galerian migration hypothesis. Quat. Sci. Rev. 2018, 180, 1–29. [Google Scholar] [CrossRef]
- Laurent, L. Appendice: Paléobotanique. In Les Bouches-du-Rhône; Encyclopédie départementale. Honoré Champion, Marseille; Archives Départementales des Bouches-du-Rhône: Paris, France; Marseille, France, 1932; pp. 339–391. [Google Scholar]
- Dubar, M.; Magnin, F. Présence d’hélicelles (Helicellinae) dans le Pliocène du Midi de la France, nouvelles données sur la dispersion du groupe au Pléistocène inférieur, implications paléoclimatiques. GÉObios 1992, 25, 357–366. [Google Scholar] [CrossRef]
- Triat-Laval, H. Contribution Pollenanalytique et L’histoire Tardi- et Post-Glaciaire de la Végétation de la Basse Vallée du Rhône; Thèse Université de Marseille: Marseille, France, 1978. [Google Scholar]
- Lurcock, P.C.; Wilson, G.S. PuffinPlot: A versatile, user-friendly program for paleomagnetic analysis. Geochem. Geophys. Geosystems 2012, 13, Q06Z45. [Google Scholar] [CrossRef]
- Channell, J.E.T.; Singer, B.S.; Jicha, B.R. Timing of Quaternary geomagnetic reversals and excursions in volcanic and sedimentary archives. Quat. Sci. Rev. 2020, 228, 1–29. [Google Scholar] [CrossRef]
- Arenas-Abad, C.; Vázquez-Urbez, M.; Pardo-Tirapu, G.; Sancho-Marcén, C. Fluvial and associated carbonate deposits. In Carbonates in Continental Settings: Facies, Environments and Processes, Developments in Sedimentology; Alonso-Zarza, A.M., Tanner, L.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 61, pp. 133–175. [Google Scholar]
- Fournier, F.; Ouass, A.; Rochette, P.; Bromblet, P.; Léonide, P.; Conesa, G.; Marié, L.; Boularand, S.; D’Ovidio, A.-M.; Vigie, B.; et al. Provenance of sculptural limestones in protohistoric Provence (SE France): Insights from carbonate rock petrography and stable isotope geochemistry. J. Archaeol. Sci. Rep. 2023, 48, 103922. [Google Scholar] [CrossRef]
- Nakagawa, T.; Brugiapaglia, E.; Digerfeldt, G.G.; Reille, M.; de Beaulieu, J.L.; Yasuda, Y. Dense-media separation as a more efficient pollen extraction method for use with organic sediment/deposit samples: Comparison with the conventional method. Boreas 1998, 27, 15–24. [Google Scholar] [CrossRef]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Laboratoire de Botanique Historique et de Palynologie: Marseille, France, 1992. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Supplement 1; Laboratoire de Botanique Historique et de Palynologie: Marseille, France, 1995. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Supplement 2; Laboratoire de Botanique Historique et de Palynologie: Marseille, France, 1998. [Google Scholar]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrenzende Gebiete; Dr. Friedrich Pfeil: Munich, Germany, 2004. [Google Scholar]
- Cugny, C.; Mazier, F.; Galop, D. Modern and fossil non-pollen palynomorphs from the Basque mountains (western Pyrenees, France): The use of coprophilous fungi to reconstruct pastoral activity. Veg. Hist. Archaeobot. 2010, 19, 391–408. [Google Scholar] [CrossRef]
- Haas, J.N. Fresh insights into the palaeoecological and palaeoclimatological value of Quaternary non-pollen palynomorphs. Veget. Hist. Archaeobot. 2010, 19, 389–560. [Google Scholar] [CrossRef]
- Van Geel, B.; Aptroot, A. Fossil ascomycetes in Quaternary deposits. Nova Hedwig. 2006, 82, 313–329. [Google Scholar] [CrossRef]
- Brewer, S.; Guiot, J.; Sánchez-Goñi, M.F.; Klotz, S. The Climate in Europe during the Eemian: A Multi-Method Approach Using Pollen Data. Quat. Sci. Rev. 2008, 27, 2303–2315. [Google Scholar] [CrossRef]
- Peyron, O.; Magny, M.; Goring, S.; Joannin, S.; de Beaulieu, J.-L.; Brugiapaglia, E.; Sadori, L.; Garfi, G.; Kouli, K.; Ioakim, C.; et al. Contrasting Patterns of Climatic Changes during the Holocene across the Italian Peninsula Reconstructed from Pollen Data. Clim. Past 2013, 9, 1233–1252. [Google Scholar] [CrossRef]
- Salonen, J.S.; Korpela, M.; Williams, J.W.; Luoto, M. Machine-Learning Based Reconstructions of Primary and Secondary Climate Variables from North American and European Fossil Pollen Data. Sci. Rep. 2019, 9, 15805. [Google Scholar] [CrossRef] [PubMed]
- Guiot, J. Methodology of the Last Climatic Cycle Reconstruction in France from Pollen Data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 80, 49–69. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Juggins, S. Weighted Averaging Partial Least Squares Regression (WA-PLS): An Improved Method for Reconstructing Environmental Variables from Species Assemblages. Hydrologia 1993, 269, 485–502. [Google Scholar] [CrossRef]
- De’ath, G. Boosted Trees for Ecological Modeling and Prediction. Ecology 2007, 88, 243–251. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A Working Guide to Boosted Regression Trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Prasad, A.M.; Iverson, L.R.; Liaw, A. Newer Classification and Regression Tree Techniques: Bagging and Random Forests for Ecological Prediction. Ecosystems 2006, 9, 181–199. [Google Scholar] [CrossRef]
- Fauquette, S.; Guiot, J.; Suc, J.-P. A Method for Climatic Reconstruction of the Mediterranean Pliocene Using Pollen Data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 144, 183–201. [Google Scholar] [CrossRef]
- Davis, B.A.S.; Brewer, S.; Stevenson, A.C.; Guiot, J. The Temperature of Europe during the Holocene Reconstructed from Pollen Data. Quat. Sci. Rev. 2003, 22, 1701–1716. [Google Scholar] [CrossRef]
- Joannin, S.; Bassinot, F.; Nebout, N.C.; Peyron, O.; Beaudouin, C. Vegetation Response to Obliquity and Precession Forcing during the Mid-Pleistocene Transition in Western Mediterranean Region (ODP Site 976). Quat. Sci. Rev. 2011, 30, 280–297. [Google Scholar] [CrossRef]
- Salonen, J.S.; Luoto, M.; Alenius, T.; Heikkilä, M.; Seppä, H.; Telford, R.J.; Birks, H.J.B. Reconstructing Palaeoclimatic Variables from Fossil Pollen Using Boosted Regression Trees: Comparison and Synthesis with Other Quantitative Reconstruction Methods. Quat. Sci. Rev. 2014, 88, 69–81. [Google Scholar] [CrossRef]
- Sinopoli, G. Vegetation and Climate Reconstruction during the Last Interglacial Complex: The Pollen Record of Lake Ohrid (Albania/Fyrom), the Oldest European Lake. Ph.D. Thesis, Università di Roma, University of Montpellier, Rome, Italy, 2017. [Google Scholar]
- Juggins, S. R Package, Version 0.9-26; Analysis of Quaternary Science Data. 2020.
- Robles, M.; Peyron, O.; Brugiapaglia, E.; Ménot, G.; Dugerdil, L.; Ollivier, V.; Ansanay-Alex, S.; Develle, A.-L.; Tozalakyan, P.; Meliksetian, K.; et al. Impact of Climate Changes on Vegetation and Human Societies during the Holocene in the South Caucasus (Vanevan, Armenia): A Multiproxy Approach Including Pollen, NPPs and brGDGTs. Quat. Sci. Rev. 2022, 277, 107297. [Google Scholar] [CrossRef]
- Chevalier, M.; Davis, B.A.S.; Heiri, O.; Seppä, H.; Chase, B.M.; Gajewski, K.; Lacourse, T.; Telford, R.J.; Finsinger, W.; Guiot, J.; et al. Pollen-Based Climate Reconstruction Techniques for Late Quaternary Studies. Earth-Sci. Rev. 2020, 210, 103384. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. R Package, version 1.3-5; Dismo: Species Distribution Modeling. 2021.
- Dugerdil, L.; Joannin, S.; Peyron, O.; Jouffroy-Bapicot, I.; Vannière, B.; Boldgiv, B.; Unkelbach, J.; Behling, H.; Ménot, G. Climate Reconstructions Based on GDGT and Pollen Surface Datasets from Mongolia and Baikal Area: Calibrations and Applicability to Extremely Cold–Dry Environments over the Late Holocene. Clim. Past 2021, 17, 1199–1226. [Google Scholar] [CrossRef]
- Vázquez Urbez, M.; Arenas, C.; Pardo, G. A sedimentary facies model for stepped, fluvial tufa systems in the Iberian Range (Spain): The Quaternary Piedra and Mesa valleys. Sedimentology 2012, 59, 502–526. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Pentecost, A. Travertine; Springer: Berlin/Heidelberg, Germany, 2005; 445p. [Google Scholar]
- Gandin, A.; Capezzuoli, E. Travertine versus calcareous tufa: Distinctive pterographic features and stable isotope signatures. Ital. J. Quat. Sci. 2008, 21, 125–136. [Google Scholar]
- Andrews, J.E.; Pedly, M.; Dennis, P.F. Palaeoenvironmental records in Holocene Spanish tufas: A stable isotope approach in search of reliable climatic archives. Sedimentology 2000, 47, 961–978. [Google Scholar] [CrossRef]
- Arenas, C.; Guttiérrez, F.; Osacar, C.; Sancho, C. Sedimentology and geochemistry of fluvio-lacustrine tufa deposits controlled by evaporite solution subsidence in the Central Ebro Depression, NE Spain. Sedimentology 2000, 47, 883–909. [Google Scholar] [CrossRef]
- Cronquist, A. An Integrated System of Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1981; p. 1262. [Google Scholar]
- Djamali, M.; Cilleros, K. Statistically significant minimum pollen count in Quaternary pollen analysis; the case of pollen-rich lake sediments. Rev. Palaeobot. Palynol. 2020, 275, 104156. [Google Scholar] [CrossRef]
- Juggins, S. 2014. Available online: https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm (accessed on 16 April 2014).
- Altolaguirre, Y.; Bruch, A.A.; Gibert, L. A long Early Pleistocene pollen record from Baza Basin (SE Spain): Major contributions to the palaeoclimate and palaeovegetation of Southern Europe. Quat. Sci. Rev. 2020, 231, 106199. [Google Scholar] [CrossRef]
- Ponel, P.; Andrieu-Ponel, V.; Djamali, M.; Lahijani, H.; Leydet, M.; Mashkour, M. Fossil beetles as possible evidence for transhumance during the middle and late Holocene in the high mountains of Talysch (Talesh) in NW Iran? Environ. Archaeol. 2013, 18, 201–210. [Google Scholar] [CrossRef]
- Vernet, J.L. History of the Pinus sylvestris and Pinus nigra ssp. salzmannii forest in the Sub-Mediterranean mountains (Grands Causses, Saint-Guilhem-le-Désert, southern Massif Central, France) based on charcoal from limestone and dolomitic deposits. Veg. Hist. Archaeobotany 2006, 16, 23–42. [Google Scholar] [CrossRef]
- INPN. Pin de salzmann (fiche I.N.P.N.: Inventaire National du Patrimoine Naturel). Available online: https://inpn.mnhn.fr/espece/cd_nom/138844 (accessed on 29 May 2024).
- Roiron, P.; Chabal, L.; Figueiral, I.; Terral, J.F.; Adam, A.A. Palaeobiogeography of Pinus nigra Arn. subsp. salzmannii (Dunal) Franco in the north-western Mediterranean Basin: A review based on macroremains. Rev. Palaeobot. Palynol. 2013, 194, 1–11. [Google Scholar] [CrossRef]
- Molinier, R. Catalogue des Plantes Vasculaires des Bouches-du-Rhône; Muséum d’Histoire Naturelle de Marseille: Marseille, France, 1981; 375p. [Google Scholar]
- Joannin, S.; Ciaranfi, N.; Stefanelli, S. Vegetation changes during the late Early Pleistocene at Montalbano Jonico (Province of Matera, southern Italy) based on pollen analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 270, 92–101. [Google Scholar] [CrossRef]
- Magri, D.; Di Rita, F.; Palombo, M.R. An Early Pleistocene interglacial record from an intermontane basin of central Italy (Scoppito, L’Aquila). Quat. Int. 2010, 225, 106–113. [Google Scholar] [CrossRef]
- Russo-Ermolli, E.; Aucelli, P.P.C.; Di Rollo, A.; Mattei, A.; Petrosino, P.; Porreca, M.; Rosskopf, C. An intergrated stratigraphical approach to the late Middle Pleistocene succession of the Sessano lacustrine basin (Molise, Italy). Quat. Int. 2010, 225, 114–127. [Google Scholar] [CrossRef]
- Orain, R.; Lebreton, V.; Russo-Ermolli, E.; Semah, A.M.; Nomade, S.; Shao, Q.; Bahain, J.J.; Thun Hohenstein, U.; Peretto, C. Hominin responses to environmental changes during the Middle Pleistocene in central and southern Italy. Clim. Past 2013, 9, 687–697. [Google Scholar] [CrossRef]
- Suc, J.-P.; Popescu, S.-M.; Fauquette, S.; Bessedik, M.; Jiménez-Moreno, G.; Bachiri-Taoufiq, N.; Zheng, Z.; Médail, F.; Klotz, S. Reconstruction of Mediterranean flora, vegetation and climate for the last 23 million years based on an extensive pollen dataset. Ecol. Mediterr. 2018, 44, 53–86. [Google Scholar] [CrossRef]
- Magri, D.; Palombo, M.R. Early to middle pleistocene dynamics of plant and mammal communities in south west Europe. Quat. Int. 2013, 288, 63–72. [Google Scholar] [CrossRef]
- Romey, C.; Vella, C.; Rochette, P.; Andrieu-Ponel, V.; Magnin, F.; Veron, A.; Landuré, C.; Anne-D’Ovidio, A.M.; Delanghe-Sabatier, D. Environmental imprints of landscape evolution and human activities during the Holocene in a small catchment of the Calanques Massif (Cassis, southern France). Holocene 2015, 25, 1454–1469. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W. Abies-Circum-Mediterranean firs in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; p. e015be7+. [Google Scholar]
- Garcia Anton, M.; Sainz Ollero, H. Pollen records from the middle Pleistocene Atapuerca site (Burgos, Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1991, 85, 199–206. [Google Scholar] [CrossRef]
- Postigo Mijarra, J.M.; Burjachs, F.; Gomez Manzaneque, F.; Morla, C. A palaeoecological interpretation of the lower-middle Pleistocene Cal Guardiola site (Terrassa, Barcelona, NE Spain) from the comparative study of wood and pollen samples. Rev. Palaeobot. Palynol. 2007, 146, 247–264. [Google Scholar] [CrossRef]
- Leroy, S.; Ambert, P.; Suc, J.P. Pollen record of the Saint-Macaire maar (Hérault, S. France): A lower Pleistocene glacial phase in the Languedoc coastal plain. Rev. Palaeobot. Palynol. 1994, 80, 149–157. [Google Scholar] [CrossRef]
- Girard, M. Résultats préliminaire de l’analyse pollinique. In L’épave romaine de La Madrague de Giens (Var). Gallia 1978, XXXIV, 112–116. [Google Scholar]
- Andrieu-Ponel, V.; Ponel, P.; Jull, A.J.T.; de Beaulieu, J.L.; Bruneton, H.; Leveau, P. Towards the reconstruction of the Holocene vegetation history of Lower Provence: Two new pollen profiles from Marais des Baux. Veg. Hist. Archaeobotany 2000, 9, 71–85. [Google Scholar] [CrossRef]
- Andrieu-Ponel, V.; Ponel, P.; Bruneton, H.; Leveau, P. Palaeoenvironments and cultural landscape of the last 2000 years reconstructed from pollen and coleopteran records in the lower Rhône valley, southern France. Holocene 2000, 10, 341–355. [Google Scholar] [CrossRef]
- Groman-Yaroslavski, I.; Weiss, E.; Nadel, D. Composite sickles and cereal harvesting methods at 23,000-years-Old Ohalo II, Israel. PLoS ONE 2016, 11, e0167151. [Google Scholar] [CrossRef] [PubMed]
- Bonnier, G.; Douin, R. Flore Complète Illustrée en Couleurs de France, Suisse et Belgique; Re-édition modern; Belin, Germany; Paris, France, 1990. [Google Scholar]
- Couplan, F. La cuisine sauvage. Comment accommoder mille plantes oubliées. In Encyclopédie des Plantes Comestibles de l’Europe; Equilibres: Condé-sur-Noireau, France, 1989; Volume 2, 512p. [Google Scholar]
- de Cortes Sanchez-Mata, M.; Tardio, J. Mediterranean Wild Edible Plants Ethnobotany and Food Composition Tables; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Hardy, K. Plant use in the Lower and Middle Palaeolithic: Food, medicine and raw materials. Quat. Sci. Rev. 2018, 191, 393–405. [Google Scholar] [CrossRef]
- Rodríguez, J.; Burjachs, F.; Cuenca Bescós, G.; García, N.; Van der Made, J.; Pérez González, A.; Blain, H.A.; Expósito, I.; López García, J.M.; García Antón, M.; et al. One million years of cultural evolution in a stable environment at Atapuerca (Burgos, Spain). Quat. Sci. Rev. 2011, 30, 1396–1412. [Google Scholar] [CrossRef]
- Pelletier, M.; Cochard, D.; Boudadi-Maligne, M.; Crochet, J.Y.; Bourguignon, L. Lower Pleistocene leporids (Lagomorpha, Mammalia) in Western Europe: New data from the Bois-de-Riquet (Lézignan-la-Cèbe, Hérault, France). Comptes Rendus Palevol 2015, 14, 371–385. [Google Scholar] [CrossRef]
- Roebroeks, W.; Villa, P. On the earliest evidence for habitual use of fire in Europe. Proc. Natl. Acad. Sci. USA 2011, 108, 5209–5214. [Google Scholar] [CrossRef]
- Gowlett, J.A.J. The discovery of fire by humans: A long and convoluted process. Philos. Trans. R. Soc. B 2016, 371, 20150164. [Google Scholar] [CrossRef]
- Bellomo, R.V.; Kean, W.F. Evidence of hominid controlled fire at the FxJj20 site complex, Karari escarpment. Appendix 4A. In Koobi Fora Research Project Volume 5: Plio-Pleistocene Archaeology; Isaac, G.L., Isaac, B., Eds.; Clarendon Press: Oxford, UK, 1997; pp. 224–233. [Google Scholar]
- Rowlett, R.M. Fire control by Homo erectus in East Africa and Asia. Acta Anthropol. Sin. 2000, 19, 198–208. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Herbert, T.D. The Mid-Pleistocene Climate Transition. Annu. Rev. Earth Planet. Sci. 2023, 51, 389–418. [Google Scholar] [CrossRef]
- Hays, J.D.; Imbrie, J.; Shackleton, N.J. Variations in the Earth’s Orbit: Pacemaker of the Ice Ages. Science 1976, 194, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Combourieu-Nebout, N.; Bertini, A.; Russo-Ermolli, E.; Peyron, O.; Klotz, S.; Montade, V.; Fauquette, S.; Allen, J.; Fusco, F.; Goring, S.; et al. Climate Changes in the Central Mediterranean and Italian Vegetation Dynamics since the Pliocene. Rev. Palaeobot. Palynol. 2015, 218, 127–147. [Google Scholar] [CrossRef]
- Kirschvink, J.L. The least-squares line and plane and the analysis of palaeomagnetic data. Geophys. J. Int. 1980, 62, 699–718. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| •Abies sp. | #Chamaerops humilis | #Laurus nobilis | •Potentilla sp. |
| *Acer monspessulanum | •Charcoal | •Ligustrum sp. | •Prunus t. |
| •Acer sp. | •Chenopodiaceae | •Lycopodium selago | •Quercus (deciduous) |
| •Alnus glutinosa t. | •Chrysophyceae | *#Magniolia vasseuri | #•Quercus ilex |
| •Alnus viridis sp. | •Cichorioideae | •Malva sp. | *Quercus pubescens |
| •Ammi sp. | •Cirsium t. | *Malus acerba | •Reseda sp. |
| •Anthemis sp. | *Cornus sanguinea | •Mentha sp. | #Ribes sp. |
| •Apiaceae | *#Corylus avellana | •Micro-foraminifera | *#Rubus idaeus |
| •Artemisia sp. | •Corylus sp. | •Myriophyllum spicatum | •Rumex sp. |
| #Arundo sp. | *Crataegus oxycantha | #Nerium oleander | •Salix sp. |
| •Aster t. | •Cyperaceae | •Odontites sp. | *Salix viminalis |
| •Atriplex t. | •Delitschia sp. | •Olea sp. | •Sambucus nigra |
| •Betula sp. | •Dipsacus sp. | •Ostrya/Carpinus orientalis | #Scolopendrium officinale |
| •Blackstonia t. | •Draba t. | •Pedicularis t. | •Secale cereale |
| •Botryococcus sp. | •Erica arborea/multiflora t. | *Phoebe barbusana | •Sinapis t. |
| *•Buxus sp. | •Euphorbia sp. | *Phragmites sp. | #Sorbus domestica |
| •Carduus t. | •Fagus sp. | •Picea sp. | •Sordaria sp. |
| •Castanea sp. | *#Ficus carica | #•Pinus halepensis t. | •Tilia sp. |
| •Cedrus sp. | •Fraxinus sp. | •Pinus (Mediterranean) | *#Tilia europaea |
| *Celtis australis | •Galium sp. | *Pinus nigra subsp. salzmannii | #Trachycarpus t. |
| •Celtis sp. | •Geranium sp. | •Pinus sylvestris | •Trifolium t. |
| •Centaurea cyanus t. | •Glomus sp. | #Pirus acerba | •Type 200 |
| •Centaurea nigra t. | •Gymnosperm micro-remains | •Pistacia sp. | •Ulex t. |
| *#Cercis siliquastrum | •HdV20d | •Plantago coronopus | •Ulmus sp. |
| •Cerealia 40 µm | *Hedera helix | •Plantago lanceolata t. | •Urtica sp. |
| •Cerealia 42.5 µm | •Hypericum sp. | •Plantago major-media t. | •Valsaria sp. |
| •Cerealia 45 µm | •Iris sp. | •Platanus sp. | •Viburnum sp. |
| •Cerealia 47.5 µm | •Juglans sp. | •Poaceae | *Viburnum tinus |
| •Cerealia 52.5 | •Juniperus sp. | *Populus alba | #Vitis vinifera subsp. sylvestris |
| •Cerealia 55 µm | •Lamiaceae | #Populus nigra | •Vitis sp. |
| •Cerealia 60 µm | *Laurus canariensis | •Populus sp. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrieu, V.; Rochette, P.; Fournier, F.; Demory, F.; Robles, M.; Peyron, O.; Fauquette, S.; Charrat, E.; Magniez, P.; Gambin, B.; et al. Vegetation, Climate and Habitability in the Marseille Basin (SE France) circa 1 Ma. Geosciences 2024, 14, 211. https://doi.org/10.3390/geosciences14080211
Andrieu V, Rochette P, Fournier F, Demory F, Robles M, Peyron O, Fauquette S, Charrat E, Magniez P, Gambin B, et al. Vegetation, Climate and Habitability in the Marseille Basin (SE France) circa 1 Ma. Geosciences. 2024; 14(8):211. https://doi.org/10.3390/geosciences14080211
Chicago/Turabian StyleAndrieu, Valérie, Pierre Rochette, François Fournier, François Demory, Mary Robles, Odile Peyron, Séverine Fauquette, Eliane Charrat, Pierre Magniez, Belinda Gambin, and et al. 2024. "Vegetation, Climate and Habitability in the Marseille Basin (SE France) circa 1 Ma" Geosciences 14, no. 8: 211. https://doi.org/10.3390/geosciences14080211
APA StyleAndrieu, V., Rochette, P., Fournier, F., Demory, F., Robles, M., Peyron, O., Fauquette, S., Charrat, E., Magniez, P., Gambin, B., & De Coignac, S. B. (2024). Vegetation, Climate and Habitability in the Marseille Basin (SE France) circa 1 Ma. Geosciences, 14(8), 211. https://doi.org/10.3390/geosciences14080211