The Bruneau Woodpile: A Miocene Phosphatized Fossil Wood Locality in Southwestern Idaho, USA



Abstract
:1. Introduction
1.1. Previous Work
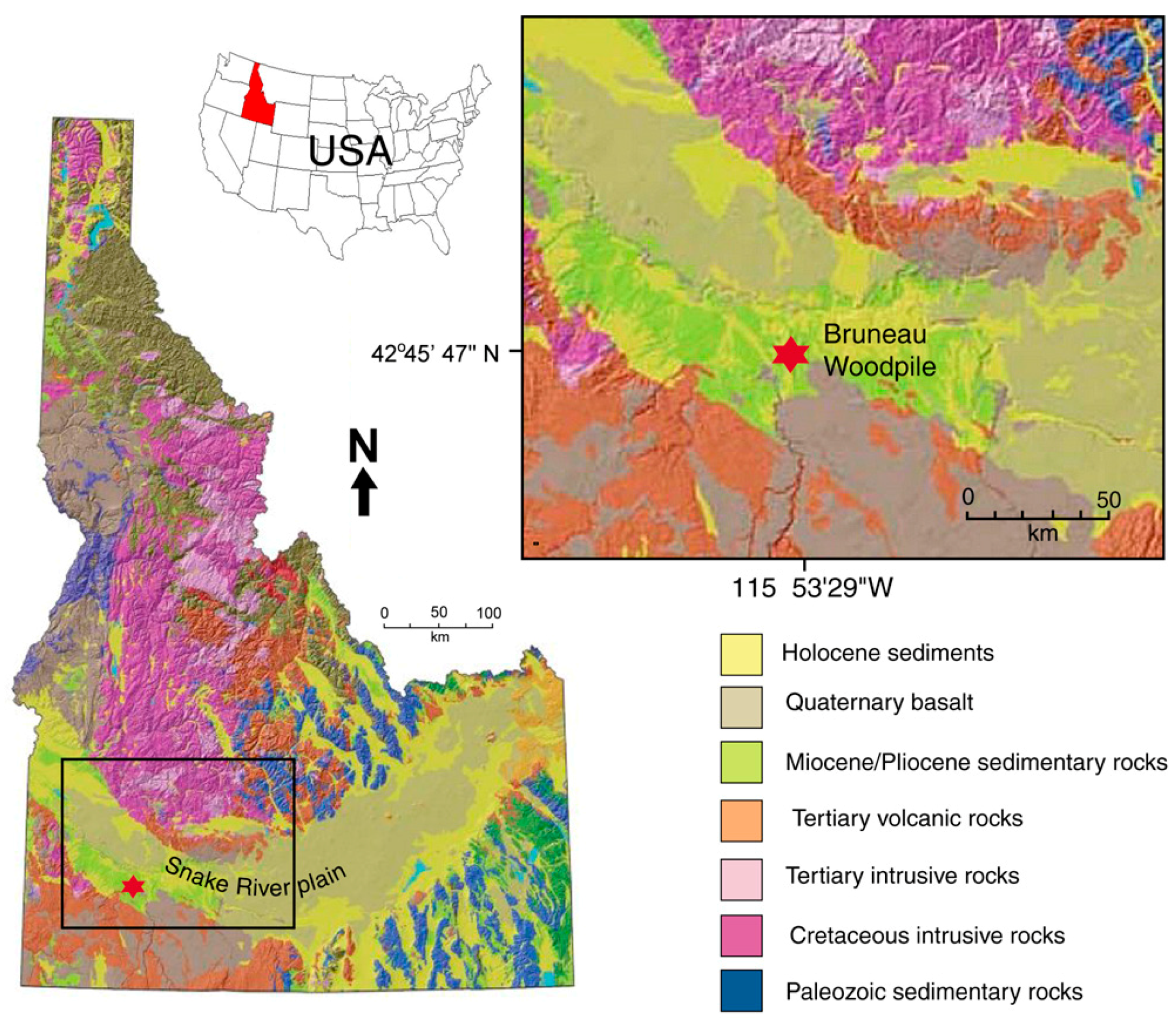
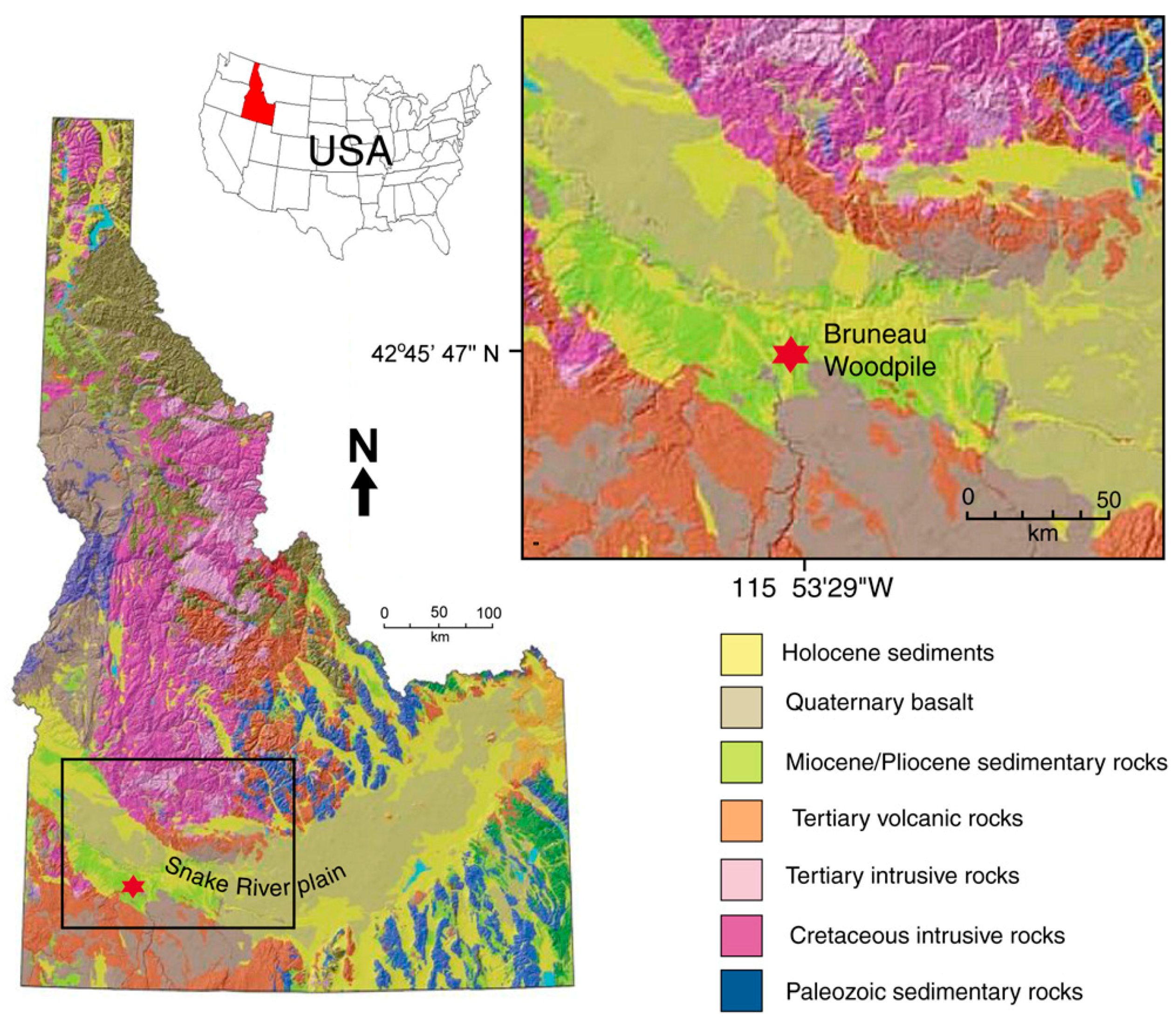
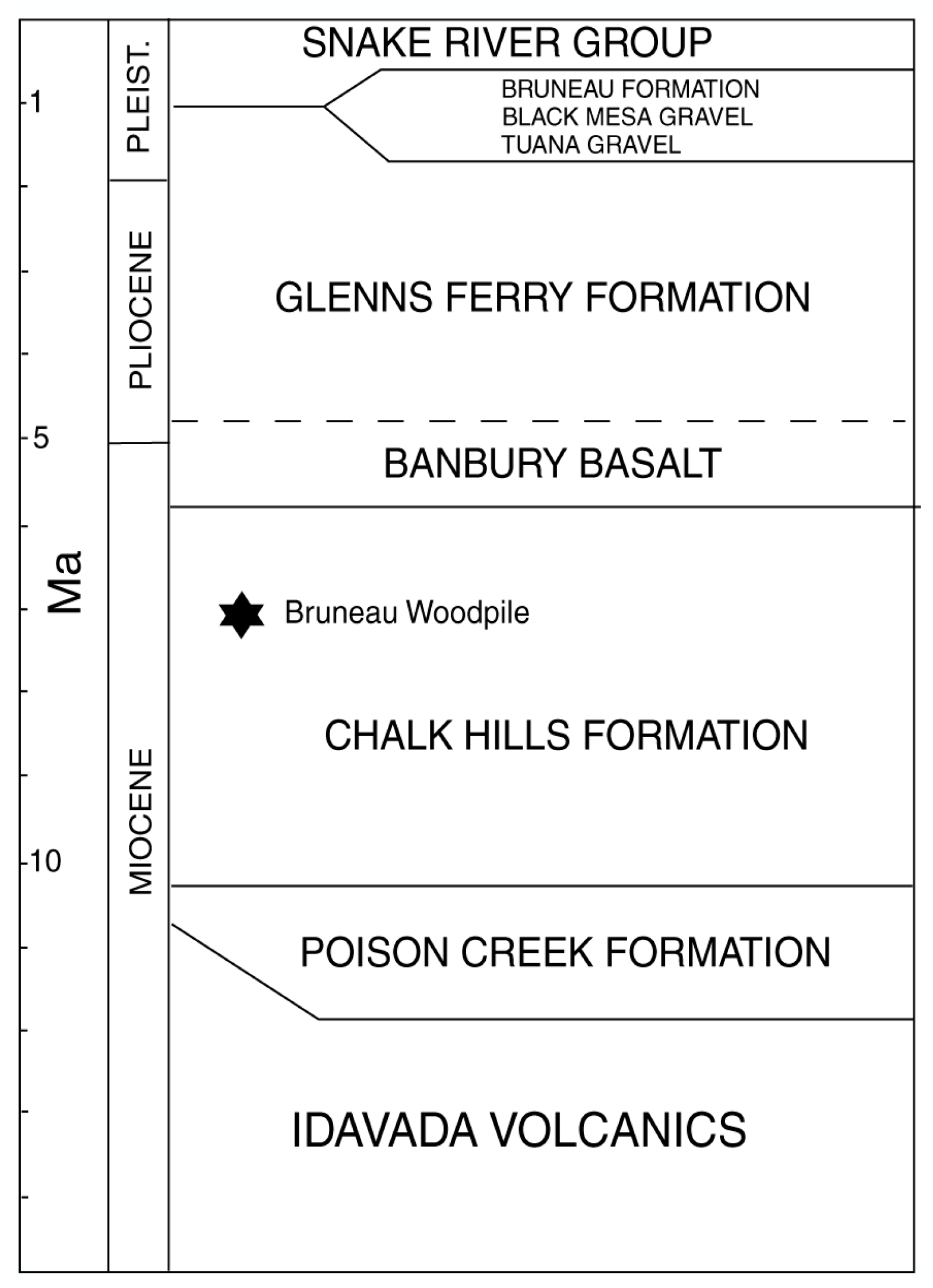
1.2. Geologic Setting
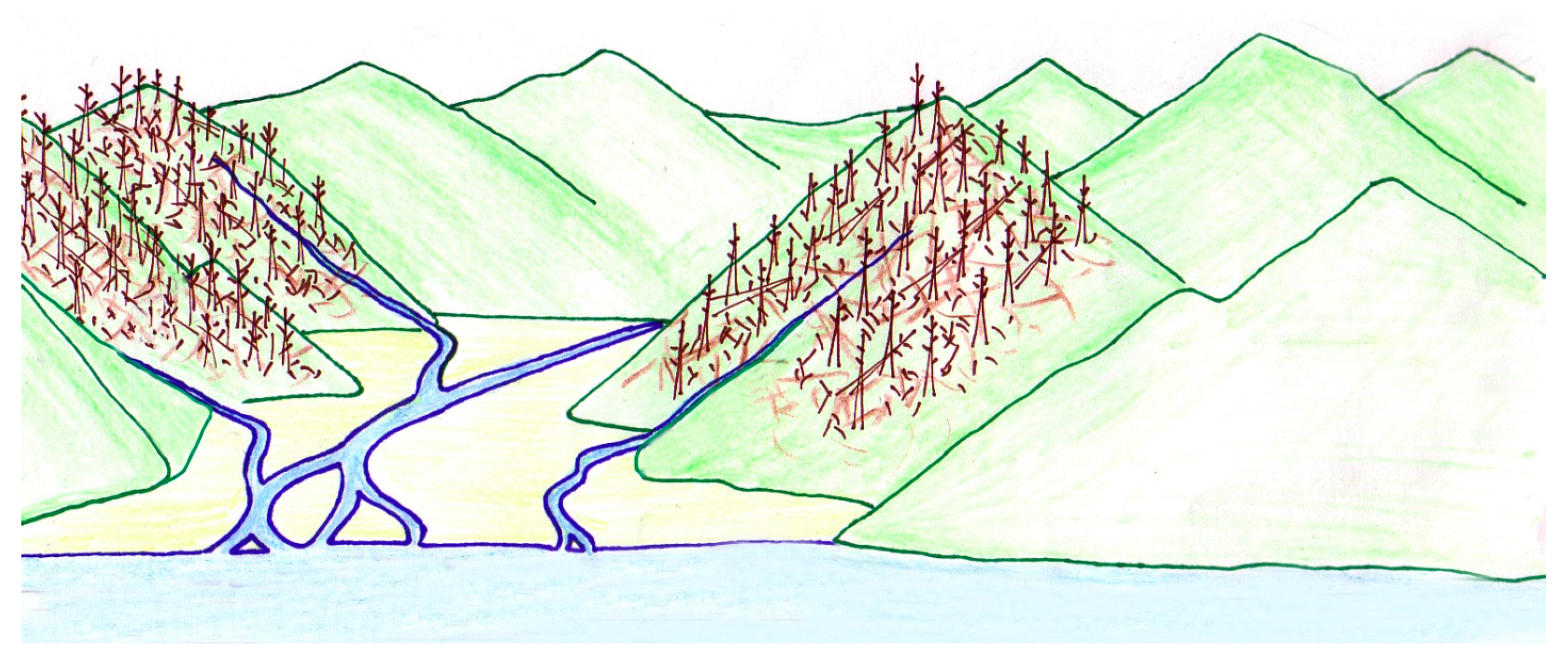
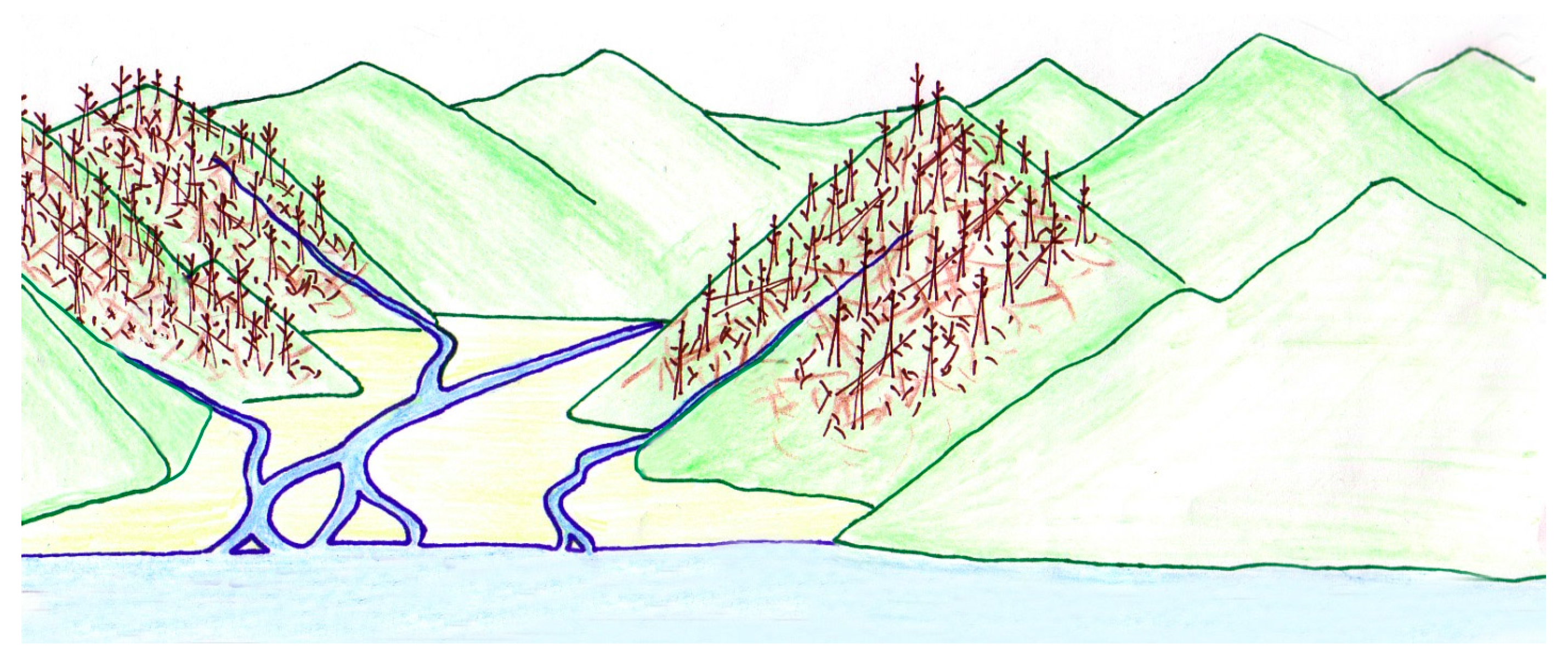
1.3. Paleoenvironment
2. Methods and Materials
3. Results
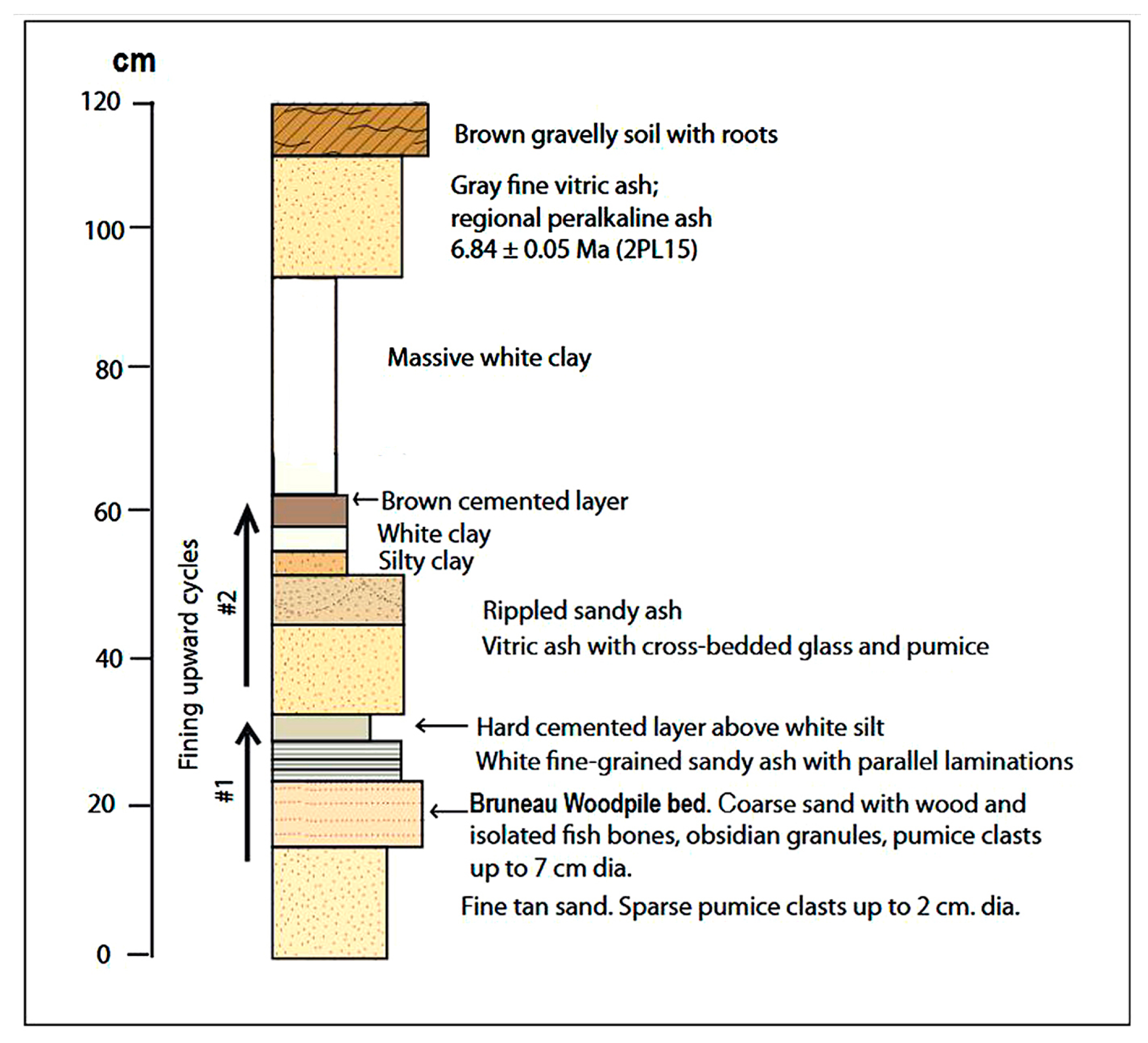
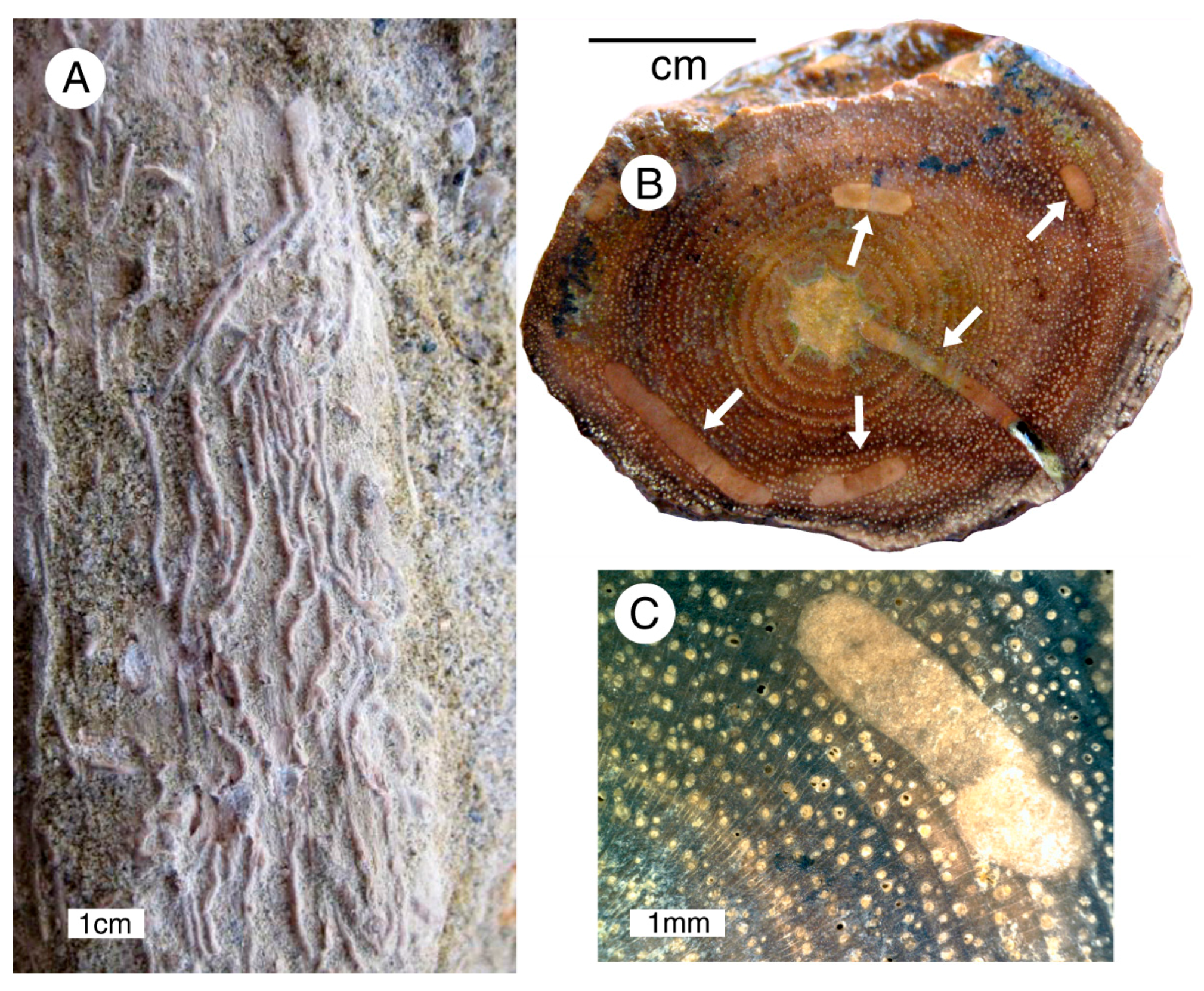
3.1. Site Description
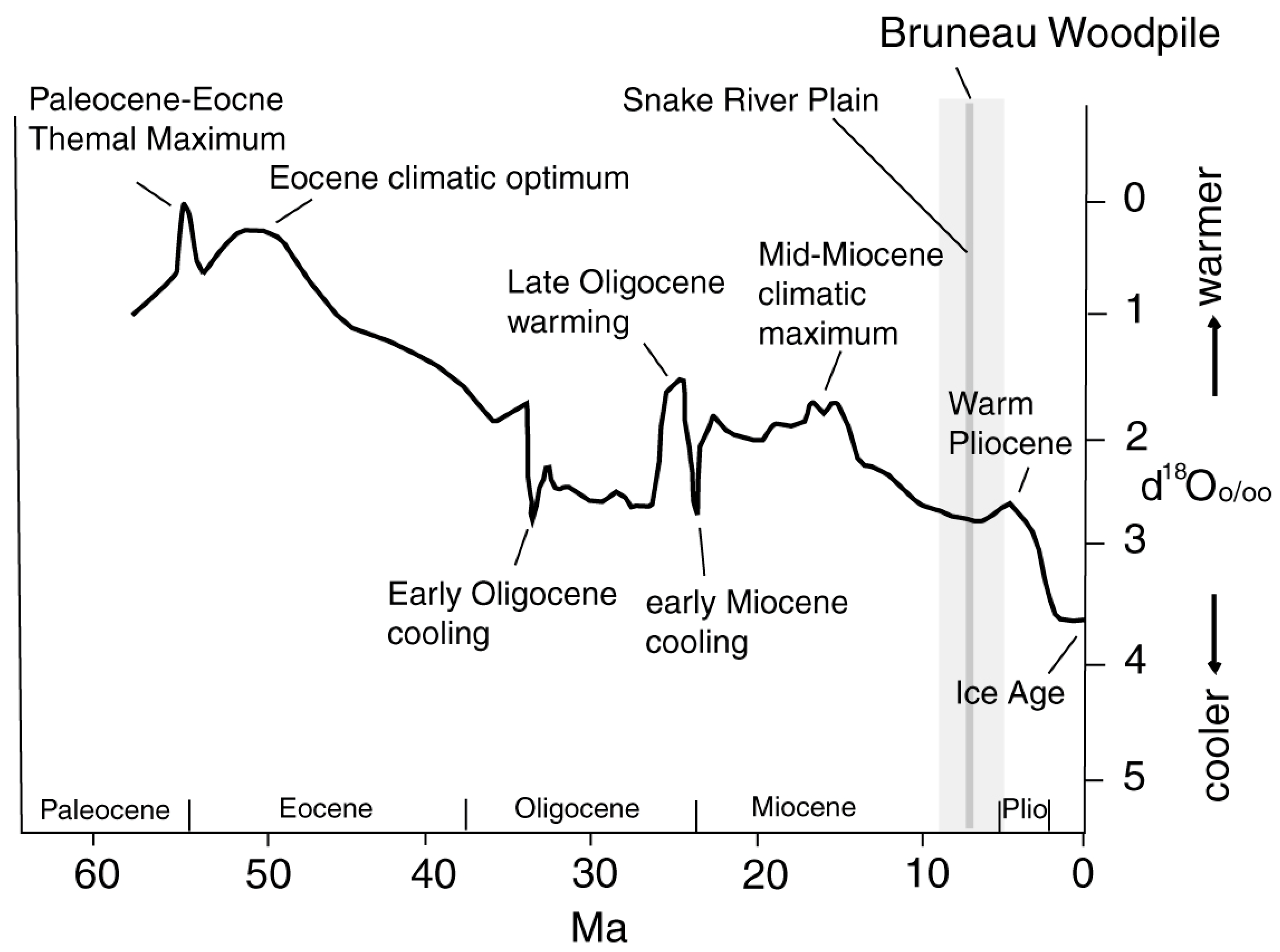
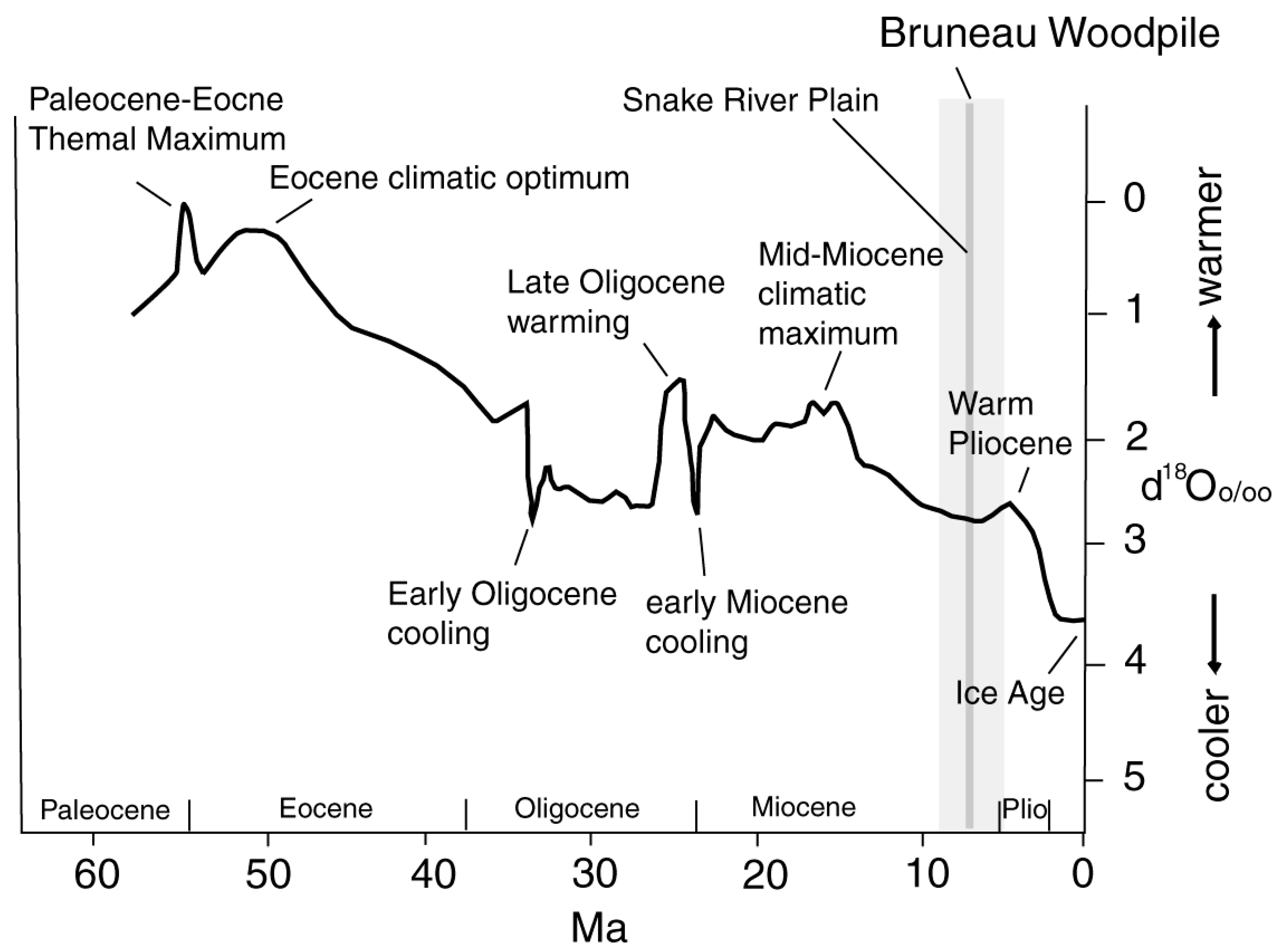
3.2. Age
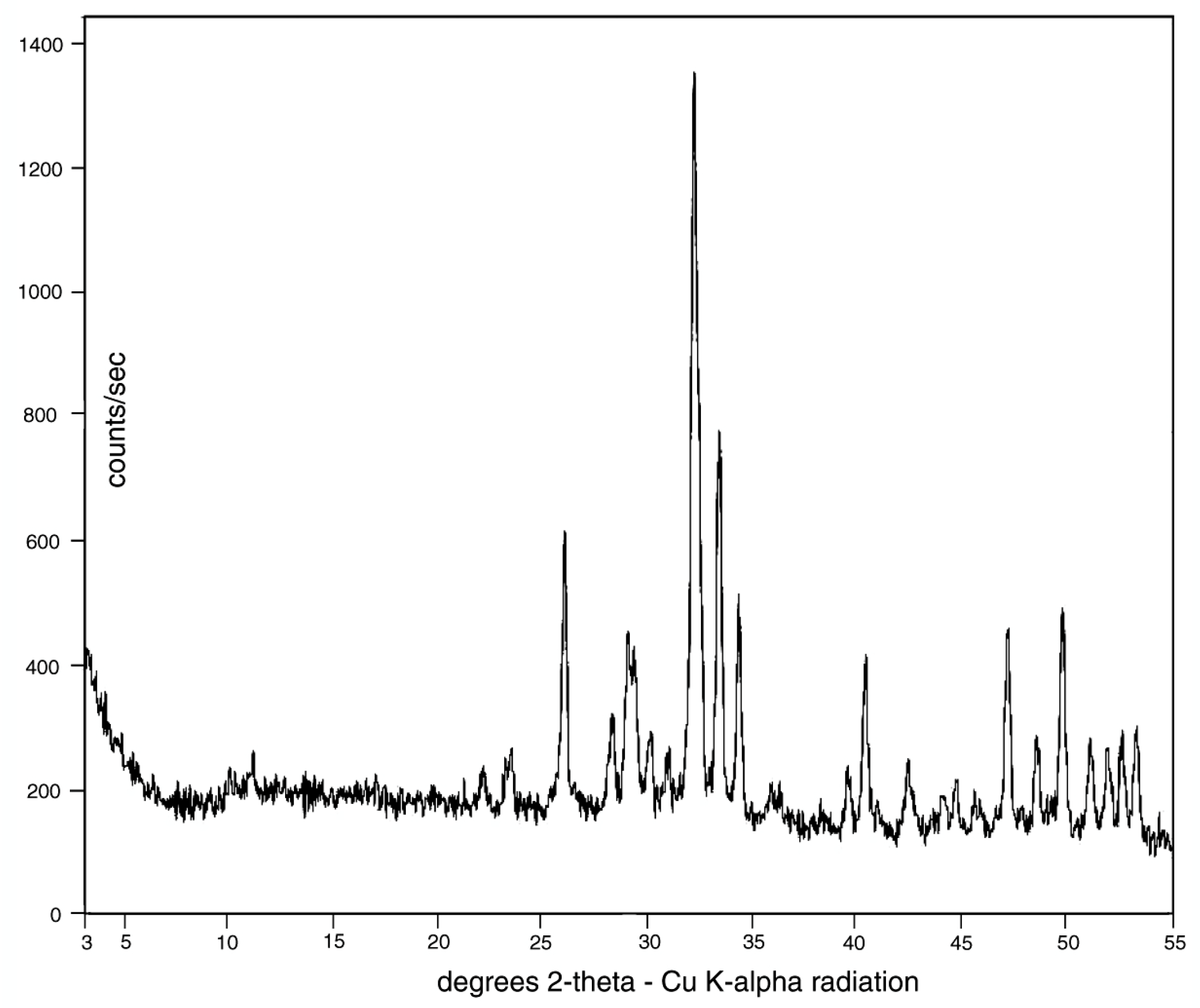
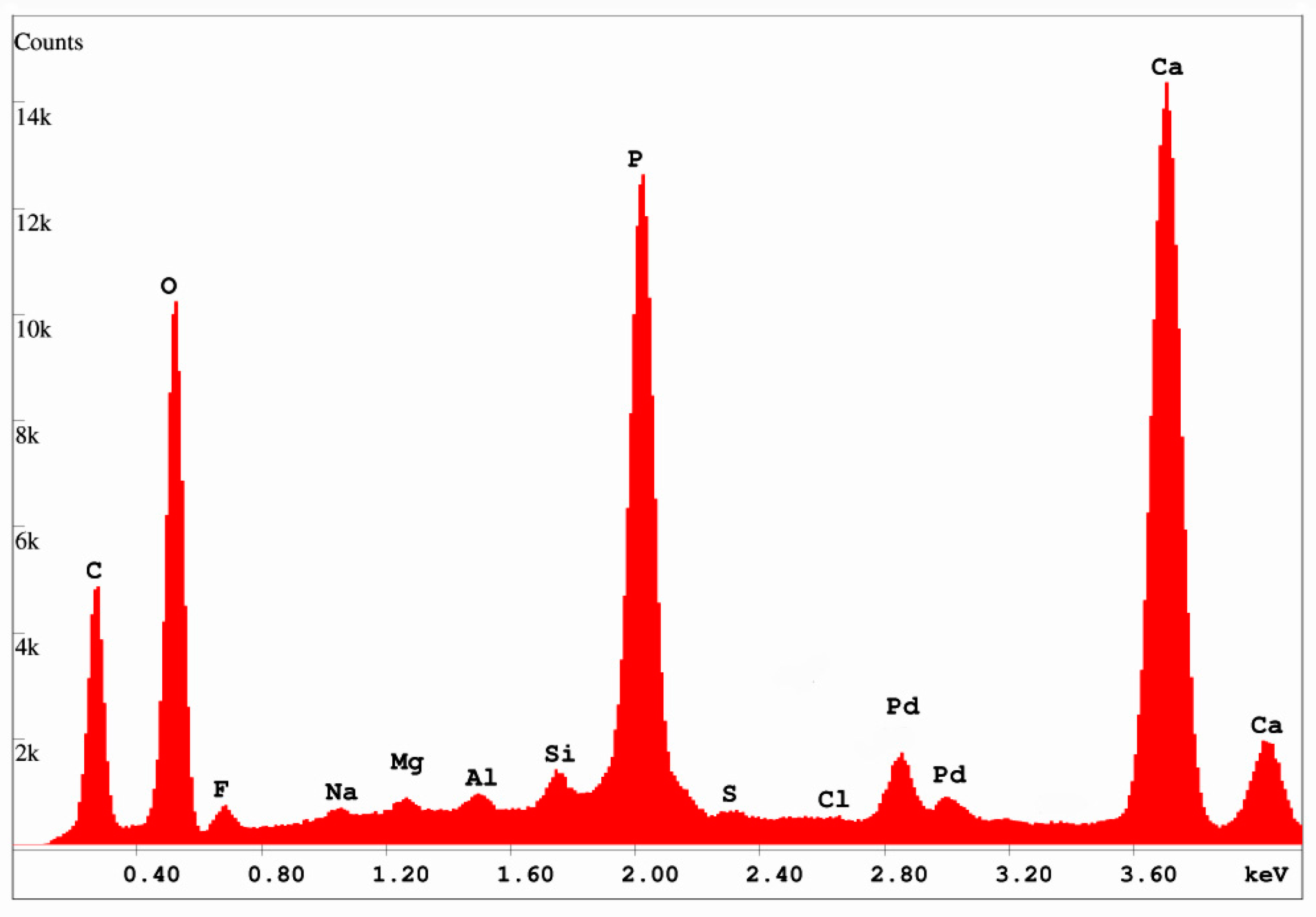
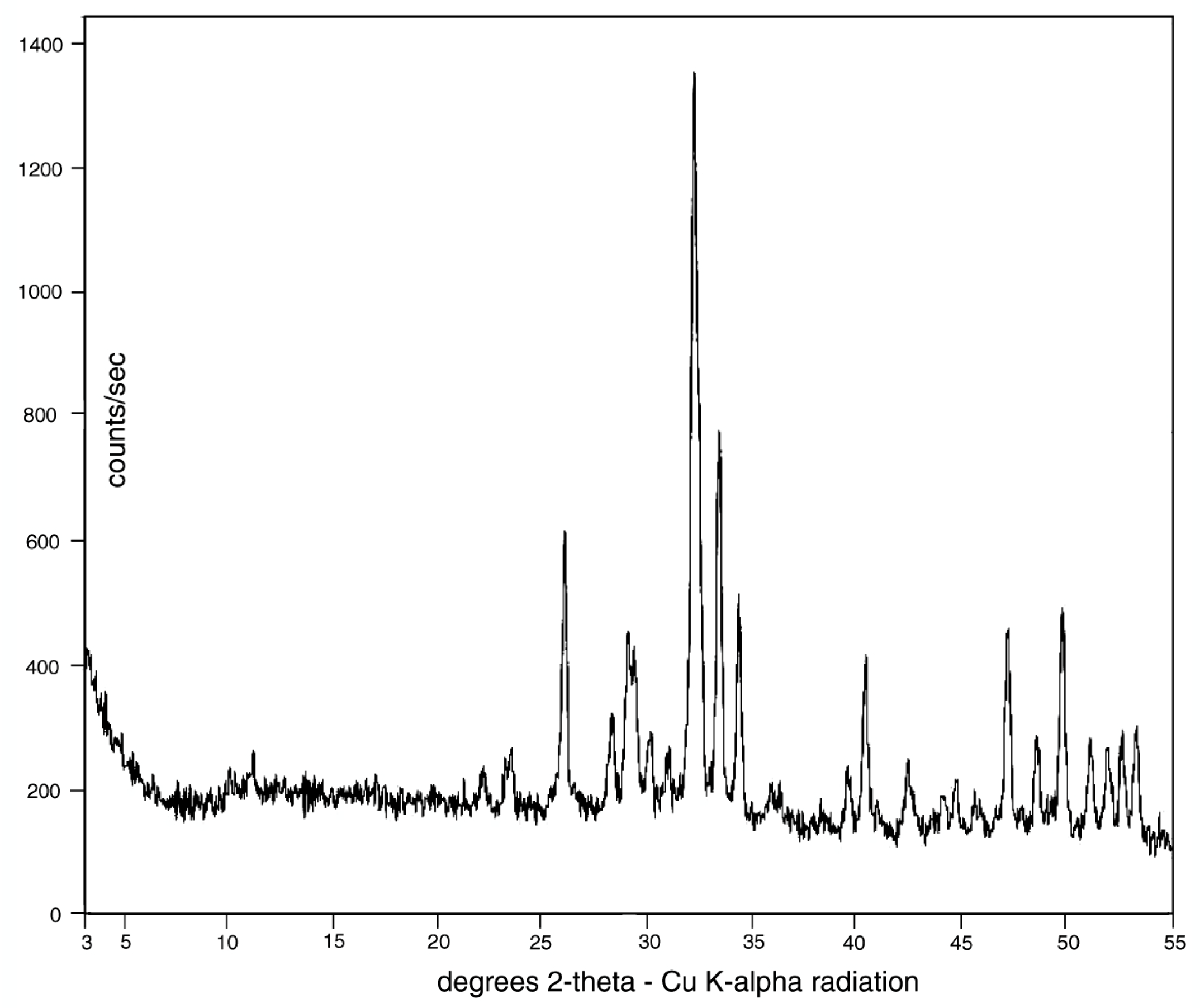
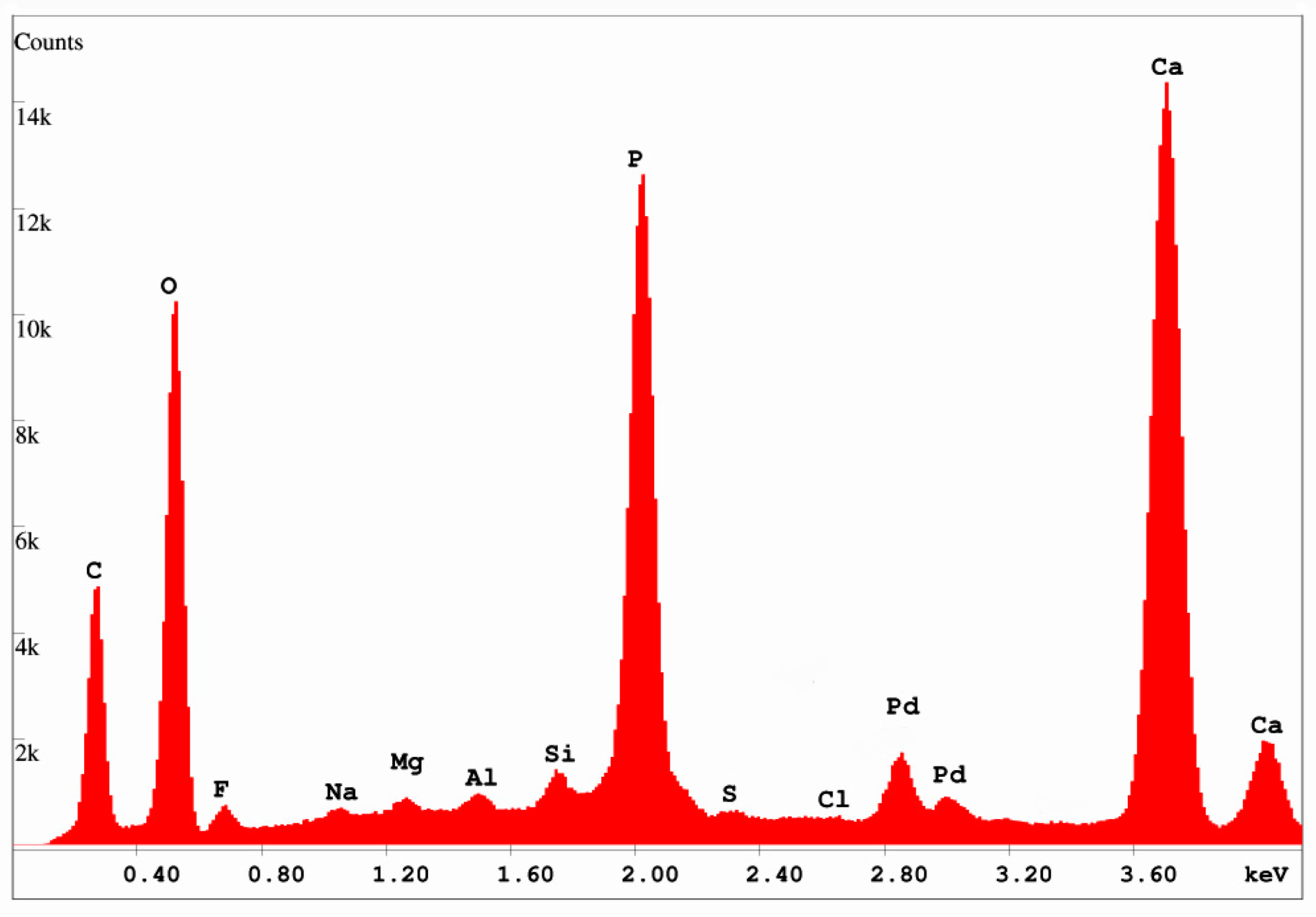
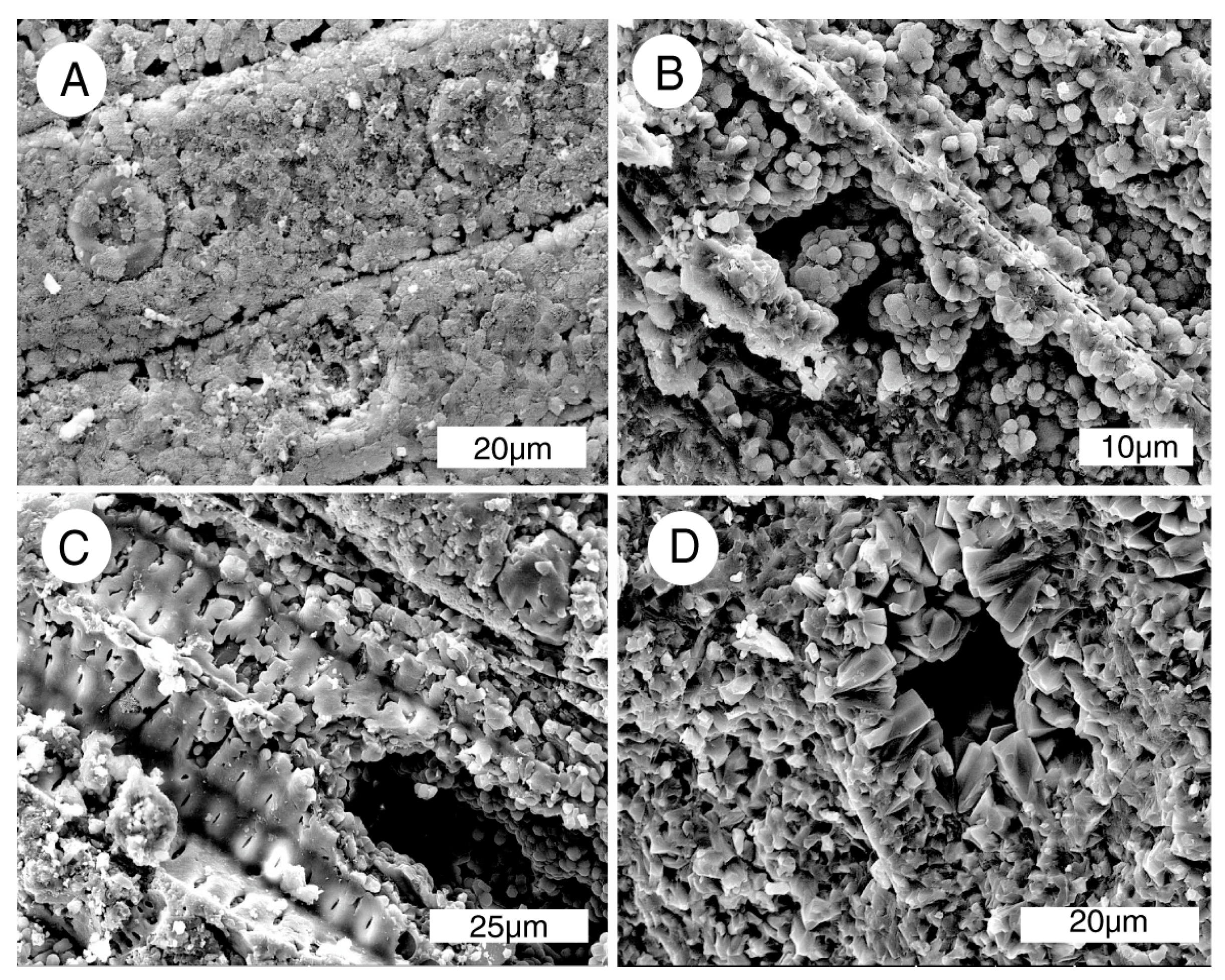
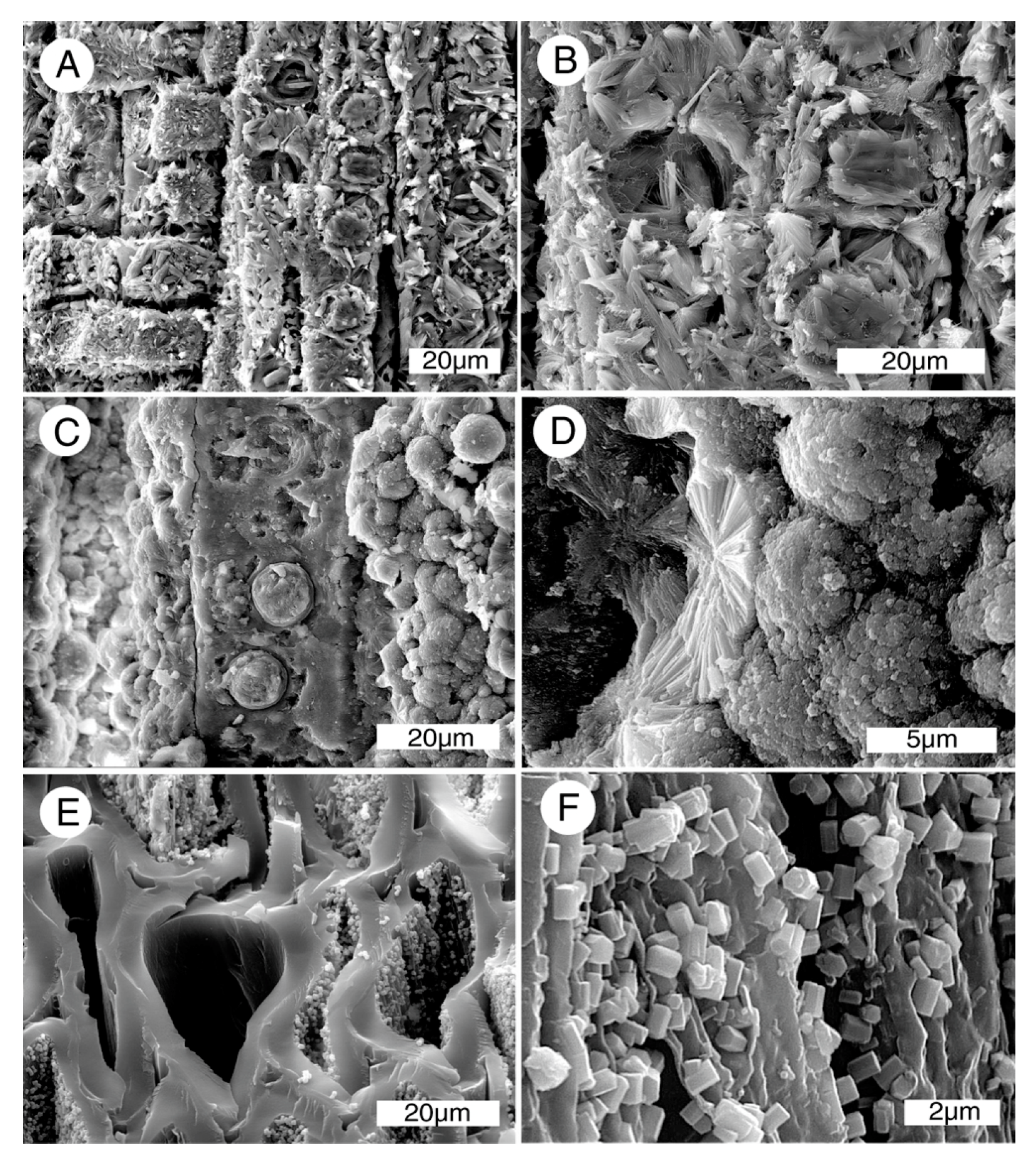
3.3. Mineralogy and Geochemistry
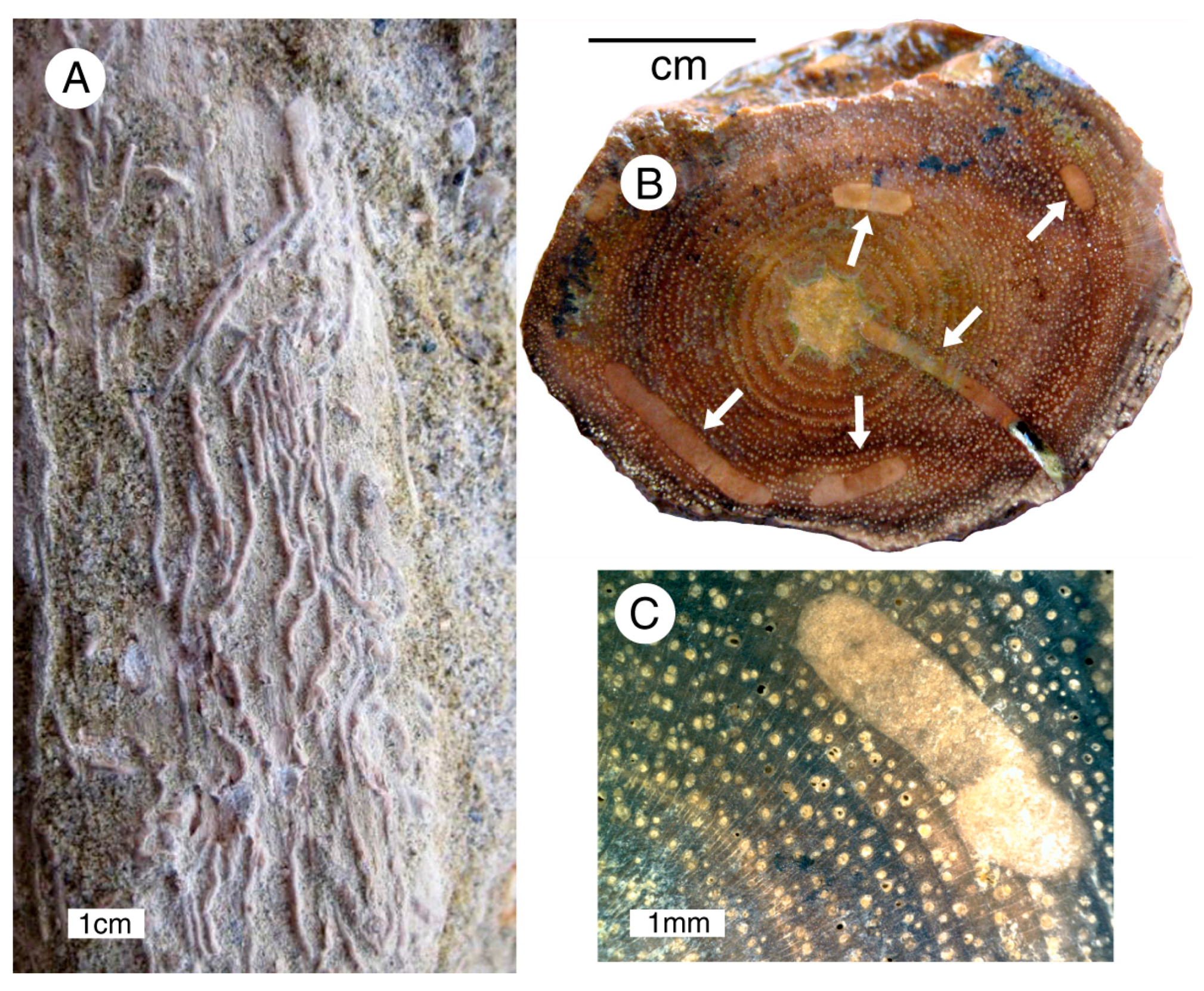
3.4. Paleobotany
Conifers
- Cupressaceae
- Pinaceae
- Piceoxylon (Spruce/Douglas fir type)
- Pinus sp. (Pine)
Angiosperms
- Berberidaceae
- cf. Berberis (Barberry/Oregon grape type)
- Fabaceae
- cf. Robinia (Black locust type)
- Fagaceae
- Quercus sp. (White oak group)
- Quercus/Lithocarpus (Live oak group)
- Juglandaceae
- Carya sp. (True hickory type)
- cf. Salicaceae
- Populus/Salix (Poplar/willow type)
- Sapindaceae
- Acer sp. (Soft maple group)
- Ulmaceae
- Ulmus sp. (Elm)
- Undetermined angiosperms
- Undetermined hardwood 1
- Undetermined hardwood 2
- Undetermined hardwood 3
- Undetermined hardwood 4
4. Discussion
4.1. Taphonomy and Sedimentology
4.2. Mineralogy and Geochemistry
4.3. Source of Phosphorus
4.4. Mineralization Process
4.5. Paleoclimate
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lewis, R.S.; Link, P.K.; Stanford, L.R.; Long, S.P. Geological Map of Idaho; Idaho Geological Survey Map 9; scale 1:750,000; Idaho Geological Survey: Moscow, ID, USA, 2012. [Google Scholar]
- Kimmel, P.G. Stratigraphy, age, and tectonic setting of the Miocene-Pliocene lacustrine sediments of the western Snake River Plain, Oregon and Idaho. In Cenozoic Geology of Idaho; Bonnichsen, B., Breckenridge, R.M., Eds.; Idaho Bureau of Mines and Geology Bulletin: Moscow, ID, USA, 1982; Volume 26, pp. 559–578. [Google Scholar]
- Leopold, E.B.; Wright, V.C. Pollen profiles of the Plio-Pleistocene transition in the Snake River Plain, Idaho. In Late Cenozoic History of the Pacific Northwest; Smiley, C.J., Ed.; Pacific Division, American Association for the Advancement of Science: Washington, DC, USA, 1985; pp. 323–348. [Google Scholar]
- Leopold, E.B.; Denton, M.F. Comparative age of grassland and steppe east and west of the northern Rocky Mountains. Ann. Mo. Bot. Gard. 1987, 74, 841–867. [Google Scholar] [CrossRef]
- Mustoe, G.E.; Leopold, E.B. Paleobotanical evidence for the post-Miocene uplift of the Cascade Range. Can. J. Earth Sci. 2014, 51, 809–824. [Google Scholar] [CrossRef]
- Buechler, W.K.; Dunn, M.T.; Rember, W.C. Late Miocene Pickett Creek flora of Owyhee County, Idaho. Univ. Mich. Contrib. Mus. Paleontol. 2007, 31, 305–362. [Google Scholar]
- Root, J.V. Bruneau wood. Gems Miner. 1971, 409, 36–37. [Google Scholar]
- Brown, R.W. A bracket fungus from the late Tertiary of southwestern Idaho. J. Wash. Acad. Sci. 1940, 30, 422–424. [Google Scholar]
- Axelrod, D.I. Cenozoic History of Some Western American Pines. Ann. Mo. Bot. Gard. 1986, 73, 565–641. [Google Scholar] [CrossRef]
- Pierce, K.L.; Morgan, L.A. The track of the Yellowstone hot spot: Volcanism, faulting, and uplift. In Regional Geology of Eastern Idaho and Western Wyoming; Link, P.K., Kuntz, M.A., Platt, L.B., Eds.; Geological Society of America Memoir: Boulder, CO, USA, 1992; Volume 179, pp. 1–53. [Google Scholar]
- Rodgers, D.W.; Ore, H.T.; Bobo, R.T.; McQuarrie, N.; Zentner, N. Extension and subsidence of the eastern Snake River Plain, Idaho. In Tectonic and Magmatic Evolution of the Snake River Plain Volcanic Province; Idaho Geological Survey Bulletin 30; Bonnichsen, B., White, C.M., McCurry, M., Eds.; Idaho Geological Survey: Moscow, ID, USA, 2002; pp. 121–155. [Google Scholar]
- Shervais, J.W.; Kauffman, J.D.; Gillerman, V.S.; Othberg, K.L.; Vetter, S.K.; Hobson, V.R.; Zarnetske, M.; Cooke, M.F.; Matthews, S.H.; Hanan, B.B. Basaltic volcanism of the central and western Snake River Plain: A guide to field relations between Twin Falls and Mountain Home, Idaho. In Interior Western United States: Geological Society of America Field Guide 6; Pederson, J., Dehler, C.M., Eds.; Geological Society of America: Boulder, CO, USA, 2005; Volume 6, pp. 27–52. [Google Scholar]
- Bonnichsen, B.; Godchaux, M.M. Late Miocene, Pliocene, and Pleistocene geology of southwestern Idaho with emphasis on basalts in the Bruneau-Jarbidge, Twin Falls and Western Snake River Plain regions. In Tectonic and Magmatic Evolution of the Snake River Plain Volcanic Province; Bonnichsen, B., White, C.M., McCurry, M., Eds.; Idaho Geological Survey Bulletin 30; Idaho Geological Survey: Moscow, UD, USA, 2002; pp. 233–312. [Google Scholar]
- Malde, H.E. Stratigraphy of the Glenns Ferry from Hammett to Hagerman, Idaho; U.S. Geological Survey Bulletin 1331-D; United States Government Printing Office: Washington, DC, USA, 1972; 19p.
- Hagerman Fossil Beds National Monument, Idaho. Available online: http://www.nps.gov/hafo/ (accessed on 30 April 2017).
- Wood, S.H.; Clemens, D.M. Geologic and tectonic history of the western Snake River Plain, Idaho and Oregon. In Tectonic and Magmatic Evolution of the Snake River Plain Volcanic Province; Bonnichsen, B., White, C.M., McCurry, M., Eds.; Idaho Geological Survey Bulletin 30; Idaho Geological Survey: Moscow, ID, USA, 2002; pp. 69–103. [Google Scholar]
- Jenks, M.D.; Bonnichsen, B.; Godchaux, M.M. Geologic Map of the Grand-View Bruneau Area, Owyhee County, Idaho; Digitally Revised Version, Idaho Geological Survey Technical Report 98-1; Idaho Geological Survey: Moscow, ID, USA, 1998.
- Malde, H.E.; Powers, H.A. Geologic Map of the Glenns Ferry-Hagerman Area, West-Central Snake River Plane Idaho; scale 1:48,000; Map I-696; U.S. Geological Survey Miscellaneous Investigations; U.S. Geological Survey: Reston, VA, USA, 1972.
- Smith, G.R.; Swirydczuk, K.; Kimmel, P.G.; Wilkinson, B.H. Fish biostratigraphy of the late Miocene to Pleistocene sediments of the western Snake River Plain, Idaho. In Cenozoic Geology of Idaho; Bonnichsen, B., Breckenridge, R.M., Eds.; Idaho Bureau of Mines Geology Bulletin: Moscow, ID, USA, 1982; Volume 26, pp. 519–542. [Google Scholar]
- Smith, G.R.; Patterson, W.P. Mio-Pliocene seasonality on the Snake River plain: Comparison of faunal and oxygen isotopic evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 107, 291–302. [Google Scholar] [CrossRef]
- Ruez, D.R., Jr. Middle Pliocene paleoclimate in the Glenns Ferry Formation of Hagerman Fossil Beds National Monument, Idaho: A baseline for evaluating faunal change. J. Ida. Acad. Sci. 2006, 42, 1–16. [Google Scholar]
- Perkins, M.E.; Brown, F.H.; Nash, W.P.; McIntosh, W.; Williams, S.K. Sequence, Age, and Source of Silicic Fallout Tuffs in Middle to Late Miocene Basins of the Northern Basin and Range Province; Geological Society of America Bulletin: Boulder, CO, USA, 1998; Volume 110, pp. 344–360. [Google Scholar]
- Panshin, A.J.; DeZeeuw, C. Textbook of Wood Technology, 3rd ed.; McGraw-Hill: New York, NY, USA, 1970; 705p. [Google Scholar]
- Wheeler, E.A. InsideWood—A web resource for hardwood anatomy. IAWA J. 2011, 32, 199–211. [Google Scholar]
- Kirkby, R. Bounty of Bruneau—Fossil treasures in southern Idaho. Gems Miner 1998, 28, 52–55. [Google Scholar]
- Nash, B.; University of Utah, Salt Lake City, UT, USA. Personal communication, 2015.
- Morgan, L.A.; McIntosh, W.C. Timing and Development of the Heise Volcanic Field, Snake River Plain, Idaho, Western USA; Geological Society of America Bulletin: Boulder, CO, USA, 2005; Volume 117, pp. 288–306. [Google Scholar] [CrossRef]
- Anders, M.H.; Saltzman, J.; Hemming, S.R. Neogene Tephra Correlations in Eastern Idaho and Wyoming Yellowstone for Hotspot-Related Volcanism and Tectonic Activity; Geological Society of America Bulletin: Boulder, CO, USA, 2009; Volume 121, pp. 837–856. [Google Scholar]
- Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.-X. The ICS International Chronostratigraphic Chart. Episodes 2013, 36, 199. [Google Scholar]
- Elliot, J.C. Structure and Chemistry of the Apatites and Other Orthophosphates. Studies in Inorganic Chemistry 18; Elsevier: New York, NY, USA, 1994; 371p. [Google Scholar]
- Hughes, J.M.; Rakovan, J. The crystal structure of apatite Ca5(PO4)3(F, OH, Cl). In Reviews in Mineralogy and Geochemistry; Kohn, M.J., Rakovan, J., Hughes, J.M., Eds.; Mineralogical Society of America: Washington, DC, USA, 2002; Volume 48, pp. 1–12. [Google Scholar]
- Godinot, C.; Tribolet, A.; Grover, R.; Ferrier-Pagès, C. Bioerosion by euendoliths decreases in phosphate-enriched skeletons of living corals. Biogeosciences 2012, 9, 2377–2384. [Google Scholar] [CrossRef] [Green Version]
- Golubic, S.; Friedmann, I.; Schneider, J. The lithobiontic ecological niche, with special reference to microorganisms. J. Sediment. Res. 1981, 5, 475–478. [Google Scholar]
- Yancey, T.M.; Mustoe, G.E.; Leopold, E.B.; Heizler, M.T. Mudflow disturbance in latest Miocene forests in Lewis County, Washington. PALAIOS 2013, 28, 343–358. [Google Scholar] [CrossRef]
- White, J.D.; Manville, V.; Wilson, C.J.N.; Houghon, B.F.; Riggs, N.R.; Ort, M. Settling and deposition of AD 181 Taupo pumice in lacustrine and associated environments. In Volcaniclastic Sedimentation in Lacustrine Settings; White, J.D.L., Riggs, N.R., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 1999; pp. 141–150. [Google Scholar]
- Furniss, R.L.; Carolin, V.M. Western Forest Insects; U.S. Department of Agriculture, Forest Service, Miscellaneous Publication: Washington, DC, USA, 1977; No. 1339; 346p.
- Andrews, H.N.; Lenz, L.W. Fossil polypores from Idaho. Ann. Mo. Bot. Gard. 1947, 34, 113–114. [Google Scholar] [CrossRef]
- Gauch, H.G. Scientific Method in Practice; Cambridge University Press: Cambridge, UK, 2003; ISBN 0–521–01708–4. [Google Scholar]
- Sandell, E.B.; Hay, M.H.; McConnell, D. The composition of francolite. Mineral. Mag. 1939, 25, 395–401. [Google Scholar] [CrossRef]
- Lucas, J.; Prévôt, L.E. Phosphates and fossil preservation. In Taphonomy: Releasing the Data Locked in the Fossil Record; Allison, P.A., Briggs, D.E.G., Eds.; Plenum Press: New York, NY, USA, 1991; pp. 389–409. [Google Scholar]
- Swirydczuk, K.; Wilkinson, B.H.; Smith, G.R. The Pliocene Glenns Ferry oolite: Lake-margin carbonate deposition in the southwestern Snake River plain. J. Sediment. Res. 1979, 49, 114–1129. [Google Scholar]
- Swirydczuk, K.; Wilkinson, B.H.; Smith, G.T. Synsedimentary lacustrine phosphorites from the Pliocene Glenns Ferry Formation of southwestern Idaho. J. Sediment. Res. 1981, 51, 1205–1214. [Google Scholar]
- Wedepohl, K.H. The composition of the continental crust. Geochim. Cosmochim. Acta 1995, 59, 1217–1232. [Google Scholar] [CrossRef]
- Fleischer, M. Recent Estimates of the Abundances of Elements in the Earth’s Crust; U.S. Geological Survey Circular 285; USGS: Reston, VA, USA, 1953; pp. 1–3.
- Dubrowsky, N.M.; Burrow, K.R.; Clark, G.M.; Gronberg, J.M.; Hamilton, P.A.; Hitt, K.J.; Mueller, D.K.; Munn, M.D.; Nolan, B.T.; Puckett, L.J.; et al. The quality of our Nation’s waters—Nutrients in the Nation’s streams and groundwater, 1992–2004; U.S. Geological Survey Circular 1350; USGS: Reston, VA, USA, 2010; 174p.
- Dunne, T.; Leopold, L.B. Water in Environmental Planning, 1st ed.; W.H. Freeman: New York, NY, USA, 1978. [Google Scholar]
- Arena, D.A. Exceptional preservation of plants and invertebrates by phosphatization, Riversleigh, Australia. PALAIOS 2008, 23, 495–502. [Google Scholar] [CrossRef]
- Sheldon, R.P.; Warner, M.A.; Thompson, M.E.; Pierce, H.W. Stratigraphic Sections of the Phosphoria Formation in Idaho, 1949. Part 1; U.S. Geological Survey Circular 304; USGS: Reston, VA, USA, 1949.
- McKelvey, V.E.; Armstrong, R.A.; Gulbrandsen, R.A.; Campbell, R.M. Stratigraphic sections of the Phosphoria Formation in Idaho, 1947–48, Part 2; Geological Survey Information Circular 301; USGS: Reston, VA, USA, 1953.
- Petrun, R.M. Field guide to the southeast Idaho phosphate district. In Guidebook to the Geology of Eastern Idaho; Hughes, S.S., Thackray, G.D., Eds.; Idaho Museum of Natural History: Moscow, ID, USA, 1999; pp. 269–280. [Google Scholar]
- Piper, D.Z.; Link, P.K. An Upwelling Model for the Phosphoria Sea: A Permian, Ocean-Margin Sea in the Northwest United States. Am. Assoc. Petroleum Geol. Bull. 2002, 86, 1217–1235. [Google Scholar]
- Gluskoter, H.J.; Pierard, L.H.; Pfefferkorn, H.W. Apatite petrifacations in Pennsylvanian shales of Illinois. J. Sediment. Res. 1970, 40, 1363–1366. [Google Scholar] [CrossRef]
- Krajewski, K.P. Organic geochemistry of a phophorite to black shale transgressive succession: Wilhelmøya and Janusfjellet Formations (Rhætian-Jurassic) in Central Spitsbergen, Arctic Ocean. Chem. Geol. 1989, 11, 331–334. [Google Scholar]
- Reolid, M.; Philippe, M.; Nagy, J.; Abad, I. Preservation of phosphatic wood remains in marine deposits of the Brentskardhaugen Bed (Middle Jurassic) from Svalbard (Boreal Realm). Facies 2010, 56, 549–566. [Google Scholar] [CrossRef]
- Scott, A.C.; Collinson, M.E. Non-destructive multiple approaches to interpret the preservation of plant fosils: Implications for calcium-rich permineralizations. J. Geol. Soc. 2003, 160, 857–862. [Google Scholar] [CrossRef]
- Jefferson, T.H. The preservation of conifer wood: Examples from the Lower Cretaceous of Antarctica. Paleontology 1987, 30, 232–249. [Google Scholar]
- Sweeney, I.J.; Chin, K.; Hower, J.C.; Budd, D.A.; Wolfe, D.G. Fossil wood from the middle Cretaceous Moreno Hill Formation: unique expressions of wood mineralization and implications for the process of wood preservation. Int. J. Coal Geol. 2009, 79, 1–17. [Google Scholar] [CrossRef]
- Pailler, D.; Flicoteaux, R.; Ambrosi, J.P.; Médus, J. Les bois fossils mio-pliocènes de Nkondo (Lac Albert, Ouganda), composition minéralogique et mode de formation. C R Acad. Sci. Paris Sci. Terre et Planets 2000, 331, 279–286. [Google Scholar]
- Koeniguer, J.C. Sur une liane plio-quaternaire du Tchad. Bulletin du Muséum National d’Histoire Naturelle, Sciences de la Terre 1973, 172, 81–91. [Google Scholar]
- Goldberg, E.D.; Parker, R.H. Phosphatized wood from the Pacific sea floor. Bull. Geol. Soc. Am. 1960, 71, 631–632. [Google Scholar] [CrossRef]
- Mustoe, G.E. Late Tertiary petrified wood from Nevada, USA: Evidence of multiple silicification pathways. Geosciences 2015, 5, 286–309. [Google Scholar] [CrossRef]
- Viney, M.; Dietrich, D.; Mustoe, G.; Link, P.; Lampke, T.; Götze, J.; Rößler, R. Multi-stage silicification of Pliocene wood: Re-examination of an 1895 Discovery from Idaho, USA. Geosciences 2016, 6, 21. [Google Scholar] [CrossRef]
- Crosby, C.H.; Bailey, J. The role of microbes in the formation of modern and ancient phosphatic mineral deposits. Front. Microbiol. 2012, 3, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Omelon, S.; Ariganello, M.; Bonucci, E.; Grynpas, M.; Nanci, A. A review of phosphate mineral nucleation in biology and geobiology. Calcif. Tissue Int. 2013, 93, 382–396. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Renaut, R.W.; Rosen, M.R. Biogenicity of silica precipitation around geysers and hot-spring vents, North Island, New Zealans. J. Sediment. Res. 1997, 67, 88–104. [Google Scholar]
- Walter, M.R.; McKloughlin, S.; Drinnan, A.N.; Farmer, J.D. Paleozoic hot spring sinter in the Drumond Basin, Queensland, Australia. Econ. Geol. 1998, 84, 135–143. [Google Scholar]
- Konhauser, K.O.; Phoenix, V.R.; Bottrell, S.H.; Adams, D.G.; Head, I.M. Microbial-silica interactions in Icelandic hot spring sinter: Possible analogues for some Precambrian iron stromatolites. Sedimentology 2001, 48, 425–433. [Google Scholar] [CrossRef]
- Guidry, S.A.; Chafetz, H.S. Depositional facies and diagentic alteration in a relict siliceous hot-spring accumulation: Examples from Yellowstone National Park, U.S.A. J. Sediment. Res. 2003, 73, 806–823. [Google Scholar] [CrossRef]
- Hinman, N.W.; Walter, M.R. Textural preservation in siliceous hot spring deposits during early diagenesis: Examples from Yellowstone National Park and Nevada, U.S.A. J. Sediment. Res. 2005, 75, 200–215. [Google Scholar] [CrossRef]
- Knoll, A.H. Exceptional preservation of photosynthetic organisms in silicified carbonates and silicified peats. Philos. Trans. R. Soc. Lond. 1985, B311, 111–122. [Google Scholar] [CrossRef]
- Harding, I.C.; Chant, L.S. Self-cemented diatom mats as agents of exceptional fossil preservation in the Oligocene Florissant lake beds, Colorado, United States. Geology 2000, 28, 195–198. [Google Scholar] [CrossRef]
- O’Brien, N.R. The Role of Biofilm in Fossil Preservation in the Late Eocene Florissant Forest, Colorado; Geological Society of America Special Paper 435; Geological Society of America: Golden, CO, USA, 2008; pp. 19–31. [Google Scholar]
- Briggs, D.E.K. The role of decay and mineralization in the preservation of soft-bodied fossils. Annu. Rev. Earth Planet. Sci. 2003, 31, 275–301. [Google Scholar] [CrossRef]
- Briggs, D.E.H.; Wilby, P.R. The role of the calcium carbonate-calcium phosphate switch in the mineralization of soft-bodied fossils. J. Geol. Soc. Lond. 1996, 153, 665–668. [Google Scholar] [CrossRef]
- Briggs, D.E.G.; Kear, A.J.; Maretill, D.M.; Wilby, P.R. Phosphatization of soft tissue in experiments and fossils. J. Geol. Soc. Lond. 1993, 150, 1035–1038. [Google Scholar] [CrossRef]
- Wilson, P.; Parry, L.A.; Vinther, J.; Edgecomb, G.D. Unveiling biases in soft-tissue phosphatization: Extensive preservation of musculature in the Cretaceous (Cenomanian) polychaete Rollinschaeta myoplena (Annelida: Amphinomidae). Palaeontology 2016, 59, 463–479. [Google Scholar] [CrossRef]
- Saggemann, J.; Bale, S.J.; Briggs, D.G.; Parkes, R.J. Controls on formation of authigentic minerals in association with decaying organic matter: An experimental approach. Geochim. Cosmochim. Acta 1999, 63, 1083–1095. [Google Scholar] [CrossRef]
- Ballhaus, C.; Gee, C.T.; Bockrath, C.; Greef, K.; Mansekldt, T.; Rhede, D. The silicification of trees in volcanic ash—An experimental study. Geochim. Cosmochim. Acta 2012, 84, 62–74. [Google Scholar] [CrossRef]
- Leo, R.F.; Barghoorn, E.S. Silicification of Wood. Bot. Mus. Leafl. Harv. Univ. 1976, 25, 1–47. [Google Scholar]
- Zachos, J.C.; Pagani, M.; Sloan, L.; Billups, K.; Thomas, E. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Graham, A. Late Cretaceous and Cenozoic History of North American Vegetation; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | SiO2 | Al2O3 | TiO2 | Fe2O3 * | MnO | MgO | CaO | K2O | Na2O | P2O5 | LOI ** |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 14.53 | 0.12 | 0.00 | 1.88 | 0.45 | 0.98 | 44.36 | 0.01 | 0.94 | 37.79 | 3.83 |
| 2 | 13.69 | 0.04 | 0.00 | 1.73 | 0.50 | 0.55 | 45.67 | 0.00 | 0.00 | 33.33 | 5.83 |
| Sample | Weight % | Atomic % | Ca:P Ratio | ||
|---|---|---|---|---|---|
| CaO | P2O5 | Ca | P | ||
| 1 | 44.36 | 37.79 | 31.72 | 16.48 | 1.92 |
| 2 | 45.67 | 33.33 | 32.65 | 14.53 | 2.25 |
| Location | Age | Setting | Reference |
|---|---|---|---|
| Illinois, USA | Jurassic | marine | [52] |
| Svalbard, Boreal Realm | Jurassic | marine | [53,54] |
| Swindon, U.K. | Jurassic | marine | [55] |
| Antarctica | Cretaceous | continental | [56] |
| New Mexico, USA | Cretaceous | continental | [57] |
| California, USA | Eocene | continental | This report |
| Australia | Miocene/Oligocene | continental | [47] |
| Nevada, USA | Miocene | continental | This report |
| Idaho, USA | Miocene | continental | This report |
| Uganda | Miocene/Pliocene | continental | [58] |
| Florida, USA | Neogene | continental | This report |
| France | Pliocene/Pleistocene | continental | [59] |
| Pacific sea floor | Holocene | marine | [60] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viney, M.; Mustoe, G.E.; Dillhoff, T.A.; Link, P.K. The Bruneau Woodpile: A Miocene Phosphatized Fossil Wood Locality in Southwestern Idaho, USA. Geosciences 2017, 7, 82. https://doi.org/10.3390/geosciences7030082
Viney M, Mustoe GE, Dillhoff TA, Link PK. The Bruneau Woodpile: A Miocene Phosphatized Fossil Wood Locality in Southwestern Idaho, USA. Geosciences. 2017; 7(3):82. https://doi.org/10.3390/geosciences7030082
Chicago/Turabian StyleViney, Mike, George E. Mustoe, Thomas A. Dillhoff, and Paul K. Link. 2017. "The Bruneau Woodpile: A Miocene Phosphatized Fossil Wood Locality in Southwestern Idaho, USA" Geosciences 7, no. 3: 82. https://doi.org/10.3390/geosciences7030082
APA StyleViney, M., Mustoe, G. E., Dillhoff, T. A., & Link, P. K. (2017). The Bruneau Woodpile: A Miocene Phosphatized Fossil Wood Locality in Southwestern Idaho, USA. Geosciences, 7(3), 82. https://doi.org/10.3390/geosciences7030082