Challenges and Opportunities of Light-Emitting Diode (LED) as Key to Modulate Antioxidant Compounds in Plants. A Review

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
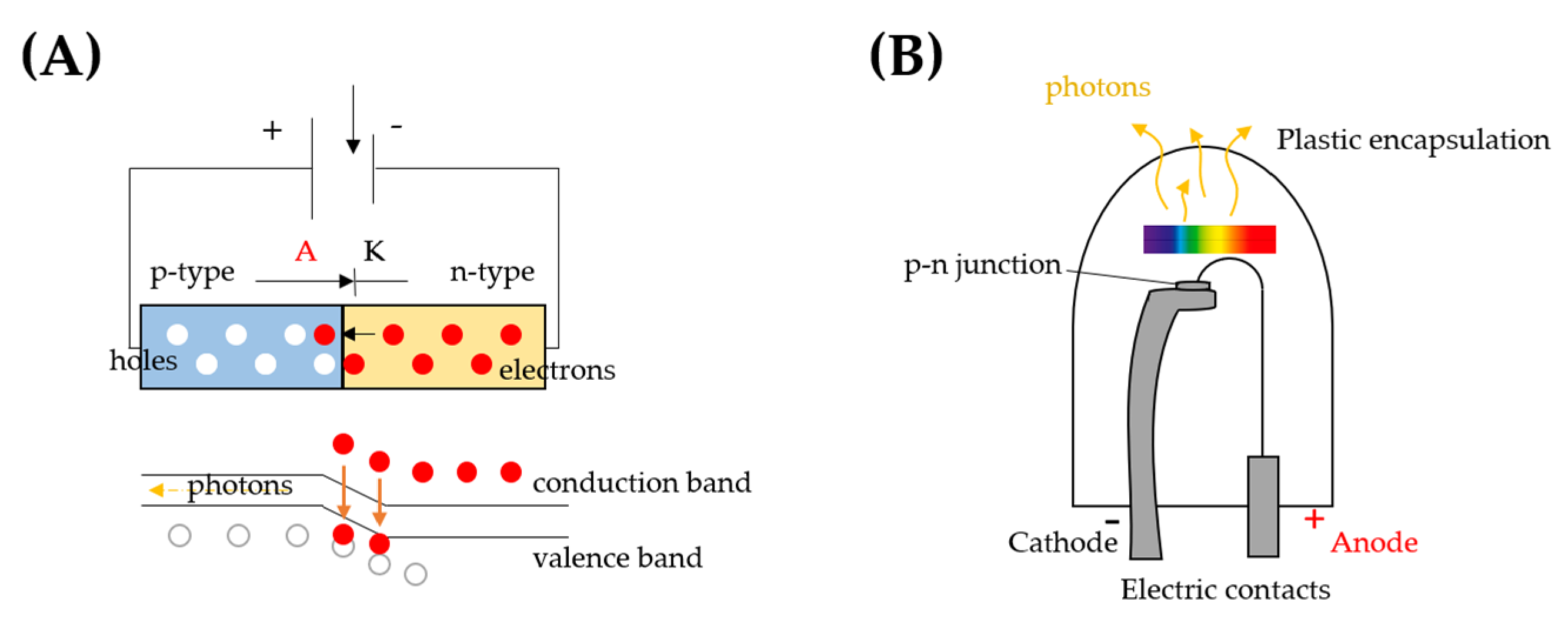
2. LED Technology
3. Plant Photoreceptors
4. LED Effects on the Antioxidants and Phytonutrients of Plants
4.1. Vitamin C
4.2. Polyphenols
4.3. Photosynthetic Pigments
4.4. Glucosinolates
5. LED Lighting: Advantages and Challenges in Plant Growth and Postharvest Management
6. LEDs as a Tool to Improve Microbiological Food Safety
7. Perspectives
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gupta, D.K.; Palma, J.M.; Corpas, F.J. Antioxidants and Antioxidant Enzymes in Higher Plants, 1st ed.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Alrifai, O.; Hao, X.; Marcone, M.F.; Tsao, R. Current review of the modulatory effects of LED lights on photosynthesis of secondary metabolites and future perspectives of microgreen vegetables. J. Agric. Food Chem. 2019, 67, 6075–6090. [Google Scholar] [CrossRef] [PubMed]
- Rienks, J.; Barbaresko, J.; Nöthlings, U. Association of polyphenol biomarkers with cardiovascular disease and mortality risk: A systematic review and meta-analysis of observational studies. Nutrients 2017, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Micek, A.; Godos, J.; Pajak, A.; Sciacca, S.; Galvano, F.; Giovannucci, E.L. Dietary flavonoid and lignan intake and mortality in prospective cohort studies: Systematic review and dose-response meta-analysis. Am. J. Epidemiol. 2017, 185, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Micek, A.; Godos, J.; Sciacca, S.; Pajak, A.; Martínez-González, M.A.; Giovannucci, E.L.; Galvano, F. Coffee consumption and risk of all-cause, cardiovascular, and cancer mortality in smokers and non-smokers: A dose-response meta-analysis. Eur. J. Epidemiol. 2016, 31, 1191–1205. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to cocoa flavanols and protection of lipids from oxidative damage (ID 652, 1372, 1506, 3143), and maintenance of normal blood pressure (ID 1507) pursuant to Article 13(1) of Regulation (EC) No. 1924/2006. EFSA J. 2010, 8, 1792–1813. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to polyphenols in olive and protection of LDL particles from oxidative damage (ID 1333, 1638, 1639, 1696, 2865), maintenance of normal blood HDL cholesterol concentrations (ID 1639), maintenance of normal blood pressure (ID 3781),“anti-inflammatory properties” (ID 1882), “contributes to the upper respiratory tract health” (ID 3468), “can help to maintain a normal function of gastrointestinal tract” (3779), and “contributes to body defences against external agents” (ID 3467) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2033. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Biswas, K.; Adhikari, S.; Tarafdar, A.; Kumar, R.; Saha, S.; Ghosh, P. Reactive Oxygen Species and Antioxidant Defence Systems in Plants: Role and Crosstalk Under Biotic Stress. In Sustainable Agriculture in the Era of Climate Change; Roychowdhury, R., Choudhury, S., Hasanuzzaman, M., Srivastava, S., Eds.; Springer: Cham, Switzerland, 2020; Chapter 12; pp. 265–292. [Google Scholar] [CrossRef]
- Kumar, N.; Singh, K.; Sharma, S.K. Antioxidants: Responses and Importance in Plant Defense System. In Sustainable Agriculture in the Era of Climate Change; Roychowdhury, R., Choudhury, S., Hasanuzzaman, M., Srivastava, S., Eds.; Springer: Cham, Switzerland, 2020; Chapter 11; pp. 251–264. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Effects of light quality on growth and phytonutrient accumulation of herbs under controlled environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G. Plant responses to light. In Plant Factory, 2nd ed.; Kozai, T., Niu, G., Takagaki, G., Eds.; Academic Press: London, UK, 2020; Chapter 9; pp. 153–166. [Google Scholar] [CrossRef]
- Yao, H.; Zhang, Y.; Yi, X.; Zuo, W.; Lei, Z.; Sui, L.; Zhang, W. Characters in light-response curves of canopy photosynthetic use efficiency of light and N in responses to plant density in field-grown cotton. Field Crops Res. 2017, 203, 192–200. [Google Scholar] [CrossRef]
- Stagnari, F.; Di Mattia, C.; Galieni, A.; Santarelli, V.; D’Egidio, S.; Pagnani, G.; Pisante, M. Light quantity and quality supplies sharply affect growth, morphological, physiological and quality traits of basil. Ind. Crop. Prod. 2018, 122, 277–289. [Google Scholar] [CrossRef]
- Taulavuori, K.; Hyöky, V.; Oksanen, J.; Taulavuori, E.; Julkunen-Tiitto, R. Species-specific differences in synthesis of flavonoids and phenolic acids under increasing periods of enhanced blue light. Environ. Exp. Bot. 2016, 121, 145–150. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morela, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Pinnola, A.; Bassi, R. Molecular mechanisms involved in plant photoprotection. Biochem. Soc. Trans. 2018, 46, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Paik, I.; Huq, E. Plant photoreceptors: Multi-functional sensory proteins and their signaling networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Ibaraki, Y. LED supplementary lighting. In Light Emitting Diodes for Agriculture, 1st ed.; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; Chapter 2; pp. 27–36. [Google Scholar] [CrossRef]
- Oncel, S.S.; Kose, A.; Vardar, F.; Torzillo, G. From the ancient tribes to modern societies, microalgae evolution from a simple food to an alternative fuel source. In Handbook of Marine Microalgae, 1st ed.; Se-Kwon, K., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 127–144. [Google Scholar]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Lee, S.W.; Seo, J.M.; Lee, M.K.; Chun, J.H.; Antonisamy, P.; Arasu, M.V.; Suzuki, T.; Al-Dhabi, N.A.; Kim, S.J. Influence of different LED lamps on the production of phenolic compounds in common and Tartary buckwheat sprouts. Ind. Crops Prod. 2014, 54, 320–326. [Google Scholar] [CrossRef]
- Wilson, J.; Hawkes, J. Display devices. In Optoelectronics, an Introduction, 3rd ed.; Prentice Hall PTR: Saddle River, NJ, USA, 1998; pp. 129–167. ISBN 0521771293. [Google Scholar]
- Schubert, E.F. LED basics: Optical properties. In Light-Emitting Diodes, 2nd ed.; Cambridge University Press: Cambridge, UK, 2006; pp. 86–190. ISBN 9780521865388. [Google Scholar]
- Rosencher, E.; Vinter, B. Light emitting diodes and laser diodes. In Optoelectronics, English Edition; Cambridge University Press: Cambridge, UK, 2004; pp. 613–656. ISBN B001FB6DT0. [Google Scholar]
- Pimputkar, S.; Speck, J.; Denbaars, S. Prospects for LED lighting. Nat. Photonics 2009, 3, 180–182. [Google Scholar] [CrossRef]
- How Do LEDs Work?—Patterned Sapphire Substrates. Available online: https://www.mikewoodconsulting.com/articles/Protocol%20Fall%202016%20-%20Patterned%20Sapphire%20Substrates.pdf (accessed on 4 November 2020).
- Franklin, K.A.; Quail, P.H. Phytochrome functions in Arabidopsis development. J. Exp. Bot. 2010, 61, 11–24. [Google Scholar] [CrossRef]
- Lin, C.; Shalitin, D. Cryptochrome Structure and Signal Transduction. Annu. Rev. Plant Biol. 2003, 54, 469–496. [Google Scholar] [CrossRef]
- Christie, J.M. Phototropin blue-light receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef]
- Kim, W.Y.; Fujiwara, S.; Suh, S.S.; Kim, J.; Kim, Y.; Han, L.; David, K.; Putterill, J.; Nam, H.G.; Somers, D.E. ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature 2007, 449, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Zoltowski, B.D.; Imaizumi, T. Structure and function of the ZTL/FKF1/LKP2 group proteins in Arabidopsis. Enzymes 2014, 35, 213–239. [Google Scholar] [CrossRef] [PubMed]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schäfer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Tilbrook, K.; Arongaus, A.B.; Binkert, M.; Heijde, M.; Yin, R.; Ulm, R. The UVR8 UV-B Photoreceptor: Perception, Signaling and Response. Arab. Book 2013, 11, e0164. [Google Scholar] [CrossRef] [PubMed]
- Christie, J.M.; Blackwood, L.; Petersen, J.; Sullivan, S. Plant flavoprotein photoreceptors. Plant Cell Physiol. 2015, 56, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.I.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef]
- Wang, Q.; Zuo, Z.; Wang, X.; Liu, Q.; Gu, L.; Oka, Y.; Lin, C. Beyond the photocycle—How cryptochromes regulate photoresponses in plants? Curr. Opin. Plant Biol. 2018, 45, 120–126. [Google Scholar] [CrossRef]
- Fiorucci, A.S.; Fankhauser, C. Plant Strategies for Enhancing Access to Sunlight. Curr. Biol. 2017, 27, R931–R940. [Google Scholar] [CrossRef]
- Nozue, K.; Covington, M.; Duek, P.; Lorrain, S.; Fankhauser, C.; Harmer, S.L.; Maloof, J.N. Rhythmic growth explained by coincidence between internal and external cues. Nature 2007, 448, 358–361. [Google Scholar] [CrossRef]
- Kinoshita, T.; Doi, M.; Suetsugu, N.; Kagawa, T.; Wada, M.; Shimazaki, K.I. phot1 and phot2 mediate blue light regulation of stomatal opening. Nature 2001, 414, 656–660. [Google Scholar] [CrossRef]
- Imaizumi, T.; Tran, H.G.; Swartz, T.E.; Briggs, W.R.; Kay, S.A. FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis. Nature 2003, 426, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Ito, S.; Song, Y.H.; Strait, A.A.; Kiba, T.; Lu, S.; Henriques, R.; Pruneda-Paz, J.L.; Chua, N.H.; Tobin, E.M.; et al. F-box proteins FKF1 and LKP2 act in concert with ZEITLUPE to control Arabidopsis clock progression. Plant Cell 2010, 22, 606–622. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, G.; Wang, H.; Wang Deng, X. Phytochrome signaling mechanisms. Arab. Book 2011, 9, e0148. [Google Scholar] [CrossRef] [PubMed]
- Yano, A.; Fujiwara, K. Plant lighting system with five wavelength-band light-emitting diodes providing photon flux density and mixing ratio control. Plant Methods 2012, 8, 46. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxygen processing in photosynthesis: Regulation and signalling. New Phytol. 2000, 146, 359–388. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Sign. 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Gallie, D.R. The role ofL-ascorbic acid recycling in responding to environmental stress and in promoting plant growth. J. Exp. Bot. 2013, 64, 433–443. [Google Scholar] [CrossRef]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in Plants: From Functions to Biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef]
- Ortiz-Espín, A.; Sánchez-Guerrero, A.; Sevilla, F.; Jiménez, A. The role of ascorbate in plant growth and development. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M.A., Munnè-Bosch, S., Burritt, D.J., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 25–45. [Google Scholar]
- Wheeler, G.L.; Jones, M.A.; Smirnoff, N. The biosynthetic pathway of vitamin C in higher plants. Nature 1998, 393, 365–369. [Google Scholar] [CrossRef]
- Chambial, S.; Dwivedi, S.; Shukla, K.K.; John, P.J.; Sharma, P. Vitamin C in disease prevention and cure: An overview. Indian J. Clin. Biochem. 2013, 28, 314–328. [Google Scholar] [CrossRef]
- Williamson, G.; Rhodes, M.J.C.; Parr, A.J. Disease prevention and plant dietary substances. In Chemicals from Plants. Perspectives on Plant Secondary Products; Waltonand, N.J., Brown, D.E., Eds.; Imperial College Press and World Scientific Publishing Co. Pte. Ltd.: London, UK, 1999; pp. 251–276. ISBN 981-02-2773-6. [Google Scholar]
- Bartoli, C.G.; Tambussi, E.A.; Diego, F.; Foyer, C.H. Control of ascorbic acid synthesis and accumulation and glutathione by the incident light red/far red ratio in Phaseolus vulgaris leaves. FEBS Lett. 2009, 583, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Mastropasqua, L.; Borraccino, G.; Bianco, L.; Paciolla, C. Light qualities and dose influence ascorbate pool size in detached oat leaves. Plant Sci. 2012, 183, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Zhang, L.; Setiawan, C.K.; Yamawaki, K.; Asai, T.; Nishikawa, F.; Maezawa, S.; Sato, H.; Kanemitsu, N.; Kato, M. Effect of red and blue LED light irradiation on ascorbate content and expression of genes related to ascorbate metabolism in postharvest broccoli. Postharvest Biol. Technol. 2014, 94, 97–103. [Google Scholar] [CrossRef]
- Loi, M.; Liuzzi, V.C.; Fanelli, F.; De Leonardis, S.; Creanza, T.M.; Ancona, N.; Paciolla, C.; Mulè, G. Effect of different light-emitting diode (LED) irradiation on the shelf life and phytonutrient content of broccoli (Brassica oleracea L. var. italica). Food Chem. 2019, 283, 206–214. [Google Scholar] [CrossRef]
- Mastropasqua, L.; Tanzarella, P.; Paciolla, C. Effects of postharvest light spectra on quality and health–related parameters in green Asparagus officinalis L. Postharvest Biol. Technol. 2016, 112, 143–151. [Google Scholar] [CrossRef]
- Mastropasqua, L.; Dipierro, N.; Paciolla, C. Effects of Darkness and Light Spectra on Nutrients and Pigments in Radish, Soybean, Mung Bean and Pumpkin Sprouts. Antioxidants 2020, 9, 558. [Google Scholar] [CrossRef]
- Dutilleul, C.; Garmier, M.; Noctor, G.; Mathieu, C.; Chétrit, P.; Foyer, C.H.; de Paepe, R. Leaf mitochondria modulate whole cell redox homeostasis, set antioxidant capacity, and determine stress resistance through altered signaling and diurnal regulation. Plant Cell 2003, 15, 1212–1226. [Google Scholar] [CrossRef]
- Tamaoki, M.; Mukai, F.; Asai, N.; Nakajima, N.; Kubo, A.; Aono, M.; Saji, H. Light-controlled expression of a gene encodingL-galactono-lactone dehydrogenase which affects ascorbate pool size in Arabidopsis thaliana. Plant Sci. 2003, 164, 1111–1117. [Google Scholar] [CrossRef]
- Massot, C.; Génard, M.; Stevens, R.; Gautier, H. Fluctuations in sugar content are not determinant in explaining variations in vitamin C in tomato fruit. Plant Physiol. Biochem. 2010, 48, 751–757. [Google Scholar] [CrossRef]
- Truffault, V.; Fry, S.C.; Stevens, R.G.; Gautier, H. Ascorbate degradation in tomato leads to accumulation of oxalate, threonate and oxalyl threonate. Plant J. 2017, 89, 996–1008. [Google Scholar] [CrossRef]
- Smirnoff, N.; Pallanca, J.E. Ascorbate metabolism in relation to oxidative stress. Biochem. Soc Trans. 1996, 24, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, Y.; Zhang, Z.; Quan, R.; Zhang, H.; Ma, L.; Deng, X.W.; Huang, R. Arabidopsis CSN5B interacts with VTC1 and modulates ascorbic acid synthesis. Plant Cell 2013, 25, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Yabuta, Y.; Mieda, T.; Rapolu, M.; Nakamura, A.; Motoki, T.; Maruta, T.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Light regulation of ascorbate biosynthesis is dependent on the photosynthetic electron transport chain but independent of sugars in Arabidopsis. J. Exp. Bot. 2007, 58, 2661–2671. [Google Scholar] [CrossRef] [PubMed]
- Di Matteo, A.; Sacco, A.; Anacleria, M.; Pezzotti, M.; Delledonne, M.; Ferrarini, A.; Frusciante, L.; Barone, A. The ascorbic acid content of tomato fruits is associated with the expression of genes involved in pectin degradation. BMC Plant Biol. 2010, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Ntagkas, N.; Woltering, E.; Bouras, S.; de Vos, R.C.; Dieleman, J.A.; Nicole, C.C.; Labrie, C.; Marcelis, L.F. Light-induced vitamin c accumulation in tomato fruits is independent of carbohydrate availability. Plants 2019, 8, 86. [Google Scholar] [CrossRef]
- Müller-Moulé, P.; Golan, T.; Niyogi, K.K. Ascorbate-deficient mutants of Arabidopsis grow in high light despite chronic photo-oxidative stress. Plant Physiol. 2014, 133, 748–760. [Google Scholar] [CrossRef]
- Karpinska, B.; Zhang, K.; Rasool, B.; Pastok, D.; Morris, J.; Verrall, S.; Hedley, P.E.; Hancock, R.D.; Foyer, C.H. The redox state of the apoplast influences the acclimation of photosynthesis and leaf metabolism to changing irradiance. Plant Cell Environ. 2017, 41, 1083–1097. [Google Scholar] [CrossRef]
- Massot, C.; Stevens, R.; Genard, M.; Longuenesse, J.J.; Gautier, H. Light affects ascorbate content and ascorbate-related gene expression in tomato leaves more than in fruits. Planta 2012, 235, 153–163. [Google Scholar] [CrossRef]
- Tabata, K.; Takaoka, T.; Esaka, M. Gene expression of ascorbic acid-related enzymes in tobacco. Phytochemistry 2002, 61, 631–635. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, G.; Yamawaki, K.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Ohta, S.; et al. Regulation of ascorbic acid metabolism by blue LED light irradiation in citrus juice sacs. Plant Sci. 2015, 233, 134–142. [Google Scholar] [CrossRef]
- Song, Y.; Qiu, K.; Gao, J.; Kuai, B. Molecular and physiological analyses of the effects of red and blue LED light irradiation on postharvest senescence of pak choi. Postharvest Biol. Technol. 2020, 164, 111155. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. The Brassica rapa Genome Sequencing Project Consortium. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Shi, L.; Chen, W.; Cao, S.; Su, Z.; Yang, Z. Effect of blue light treatment on fruit quality, antioxidant enzymes and radical-scavenging activity in strawberry fruit. Sci. Hortic. 2014, 175, 181–186. [Google Scholar] [CrossRef]
- Yan, Z.N.; He, D.X.; Niu, G.H.; Zhou, Q.; Qu, Y.H. Growth, nutritional quality, and energy use efficiency in two lettuce cultivars as influenced by white plus red versus red plus blue LEDs. Int. J. Agric. Biol. Eng. 2020, 13, 33–40. [Google Scholar] [CrossRef]
- Zha, L.; Liu, W.; Yang, Q.; Zhang, Y.; Zhou, C.; Shao, M. Regulation of Ascorbate Accumulation and Metabolism in Lettuce by the Red:Blue Ratio of Continuous Light Using LEDs. Front. Plant Sci. 2020, 11, 704. [Google Scholar] [CrossRef]
- Li, H.M.; Tang, C.M.; Xu, Z.G.; Liu, X.Y.; Han, X.L. Effects of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262–273. [Google Scholar] [CrossRef]
- Yang, X.J.; Liu, S.Q.; Zhang, Z.K.; Ma, L.; Zhang, Y.; Wei, H. Effects of different light emitting diode sources on nutritional quality of garlic seedling. Acta Nutr. Sin. 2010, 32, 518–520. [Google Scholar]
- Zhang, L.W.; Liu, S.Q.; Zhang, Z.K.; Yang, R.; Yang, X.J. Dynamic effects of different light qualities on pea sprouts quality. North. Hortic. 2010, 8, 4–7. [Google Scholar]
- Samuolienė, G.; Urbonavičiūtė, A.; Brazaitytė, A.; Šabajevienė, G.; Sakalauskaitė, J.; Duchovskis, P. The impact of LED illumination on antioxidant properties of sprouted seeds. Cent. Eur. J. Biol. 2011, 6, 68–74. [Google Scholar] [CrossRef]
- Ntagkas, N.; Min, Q.; Woltering, E.J.; Labrie, C.; Nicole, C.C.S.; Marcelis, L.F.M. Illuminating tomato fruit enhances fruit vitamin C content. Acta Hortic. 2016, 1134, 351–356. [Google Scholar] [CrossRef]
- Saengtharatip, S.; Goto, N.; Kozai, T.; Yamori, W. Green light penetrates inside crisp head lettuce leading to chlorophyll and ascorbic acid content enhancement. Acta Hortic. 2020, 1273, 261–270. [Google Scholar] [CrossRef]
- Kokalj, D.; Hribar, J.; Cigic, B.; Zlatic, E.; Demsar, L.; Sinkovic, L.; Sircelj, H.; Bizjak, G.; Vidrih, R. Influence of yellow light-emitting diodes at 590 nm on storage of apple, tomato and bell pepper fruit. Food Technol. Biotechnol. 2016, 54, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Ha, J.Y.; Oh, J.E.; Cho, M.S. The effect of LED irradiation on the quality of cabbage stored at a low temperature. Food Sci. Biotechnol. 2014, 23, 1087–1093. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 5, 270–278. [Google Scholar] [CrossRef]
- Xu, Z.; Rothstei, S.J. ROS-Induced anthocyanin production provides feedback protection by scavenging ROS and maintaining photosynthetic capacity in Arabidopsis. Plant Signal Behav. 2018, 13. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Li, X.X.; Chu, Y.N.; Zhang, M.X.; Wen, Y.Q.; Duan, C.Q.; Pan, Q.H. Three types of ultraviolet irradiation differentially promote expression of shikimate pathway genes and production of anthocyanins in grape berries. Plant. Physiol. Biochem. 2012, 57, 74–83. [Google Scholar] [CrossRef]
- Xu, J.; Su, X.; Li, Y.; Sun, X.; Wang, D.; Wang, W. Response of bioactive phytochemicals in vegetables and fruits to environmental factors. Eur. J. Nutr. Food Saf. 2019, 9, 233–247. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Acharya, J.; Rechner, O.; Neugart, S.; Schreiner, M.; Poehling, H.M. Effects of light-emitting diode treatments on Brevicoryne brassicae performance mediated by secondary metabolites in Brussels sprouts. J. Plant Dis. Prot. 2016, 123, 321–330. [Google Scholar] [CrossRef]
- Nam, T.G.; Kim, D.O.; Eom, S.H. Effects of light sources on major flavonoids and antioxidant activity in common buckwheat sprouts. Food Sci. Biotechnol. 2018, 27, 169–176. [Google Scholar] [CrossRef]
- Taulavuori, K.; Pyysalo, A.; Taulavuori, E.; Julkunen-Tiitto, R. Responses of phenolic acid and flavonoid synthesis to blue and blue-violet light depends on plant species. Environ. Exp. Bot. 2018, 150, 183–187. [Google Scholar] [CrossRef]
- Kim, E.; Park, S.; Park, B.; Lee, Y.; Oh, M.M. Growth and antioxidant phenolic compounds in cherry tomato seedlings grown under monochromatic light-emitting diodes. Hortic. Environ. Biotechnol. 2014, 55, 506–513. [Google Scholar] [CrossRef]
- Wang, P.; Chen, S.; Gu, M.; Chen, X.; Chen, X.; Yang, J.; Ye, N. Exploration of the Effects of Different Blue LED Light Intensities on Flavonoid and Lipid Metabolism in Tea Plants via Transcriptomics and Metabolomics. Int. J. Mol. Sci. 2020, 21, 4606. [Google Scholar] [CrossRef] [PubMed]
- Amoozgar, A.; Mohammadi, A.; Sabzalian, M.R. Impact of light-emitting diode irradiation on photosynthesis, phytochemical composition and mineral element content of lettuce cv. Grizzly. Photosynthetica 2017, 55, 85–95. [Google Scholar] [CrossRef]
- Xie, D.; Chen, L.; Zhou, C.; Tarin, M.W.K.; Yang, D.; Ren, K.; Zheng, Y. Transcriptomic and metabolomic profiling reveals the effect of LED light quality on morphological traits, and phenylpropanoid-derived compounds accumulation in Sarcandra glabra seedlings. BMC Plant Biol. 2020, 20, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hu, L.; Jiang, D.; Xi, W. Effect of Post-Harvest LED and UV Light Irradiation on the Accumulation of Flavonoids and Limonoids in the Segments of Newhall Navel Oranges (Citrus sinensis Osbeck). Molecules 2019, 24, 1755. [Google Scholar] [CrossRef]
- Li, H.; Lin, Y.; Chen, X.; Bai, Y.; Wang, C.; Xu, X.; Lai, Z. Effects of blue light on flavonoid accumulation linked to the expression of miR393, miR394 and miR395 in longan embryogenic calli. PLoS ONE 2018, 13, e0191444. [Google Scholar] [CrossRef]
- Kokalj, D.; Zlatić, E.; Cigić, B.; Vidrih, R. Postharvest light-emitting diode irradiation of sweet cherries (Prunus avium L.) promotes accumulation of anthocyanins. Postharvest Biol. Technol. 2019, 148, 192–199. [Google Scholar] [CrossRef]
- Maroga, G.M.; Soundy, P.; Sivakumar, D. Different Postharvest Responses of Fresh-Cut Sweet Peppers Related to Quality and Antioxidant and Phenylalanine Ammonia Lyase Activities during Exposure to Light-Emitting Diode Treatments. Foods 2019, 8, 359. [Google Scholar] [CrossRef]
- Wilawan, N.; Ngamwonglumlert, L.; Devahastin, S.; Chiewchan, N. Changes in enzyme activities and amino acids and their relations with phenolic compounds contents in okra treated by LED lights of different colors. Food Bioprocess Technol. 2019, 12, 1945–1954. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Kim, S.A.; Yun, H.K. Inhibition of Botrytis cinerea and accumulation of stilbene compounds by light-emitting diodes of grapevine leaves and differential expression of defense-related genes. Eur. J. Plant Pathol. 2015, 143, 753–765. [Google Scholar] [CrossRef]
- Gam, D.T.; Khoi, P.H.; Ngoc, P.B.; Linh, L.K.; Hung, N.K.; Anh, P.T.L.; Ha, C.H. LED Lights Promote Growth and Flavonoid Accumulation of Anoectochilus roxburghii and Are Linked to the Enhanced Expression of Several Related Genes. Plants 2020, 9, 1344. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.D.; Pradhan, S. Regulation of gene expression by LED lighting. In Light Emitting Diodes for Agriculture, 1st ed.; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; Chapter 10; pp. 237–258. [Google Scholar]
- Liu, J.; Osbourn, A.; Ma, P. MYB transcription factors as regulators of phenylpropanoid metabolism in plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Xu, F.; Zhou, W. Effect of LED irradiation on the ripening and nutritional quality of postharvest banana fruit. J. Sci. Food Agric. 2018, 98, 5486–5493. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Yao, D.; Xu, F.; Wang, H.; Zheng, Y. Effect of light on quality and bioactive compounds in postharvest broccoli florets. Food Chem. 2015, 172, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Samuolienė, G.; Viršilė, A.; Brazaitytė, A.; Jankauskienė, J.; Sakalauskienė, S.; Vaštakaitė, V.; Novickovas, A.; Viškeliene, A.; Sasnauskas, A.; Duchovskisa, P.; et al. Blue light dosage affects carotenoids and tocopherols in microgreens. Food Chem. 2017, 228, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Son, K.H.; Oh, M.M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Ouzounis, T.; Razi Parjikolaei, B.; Fretté, X.; Rosenqvist, E.; Ottosen, C.O. Predawn and high intensity application of supplemental blue light decreases the quantum yield of PSII and enhances the amount of phenolic acids, flavonoids, and pigments in Lactuca sativa. Front. Plant Sci. 2015, 6, 19. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.; Hu, T.; Zhang, S.; Zhang, Y.; Zhao, T.; Kang, Y. The influence of light-emitting diodes on the phenolic compounds and antioxidant activities in pea sprouts. J. Funct. Food 2016, 25, 459–465. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, N.S.; Park, J.S.; Lee, S.Y.; Lee, J.W.; Park, S.U. Effects of light-emitting diodes on the accumulation of glucosinolates and phenolic compounds in sprouting canola (Brassica napus L.). Foods 2019, 8, 76. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Gross, J. Pigments in Vegetables: Chlorophylls and Carotenoids; Springer Science & Business Media: New York, NY, USA, 2012; ISBN 1461520339. [Google Scholar]
- Croft, H.; Chen, J.M. Leaf pigment content. In Reference Module in Earth Systems and Environmental Sciences; Elsevier Inc.: Oxford, UK, 2017; pp. 1–22. [Google Scholar]
- Von Wettstein, D.; Gough, S.; Kannangara, C.G. Chlorophyll biosynthesis. Plant Cell 1995, 7, 1039–1057. [Google Scholar] [CrossRef]
- Willows, R.D. Biosynthesis of chlorophylls from protoporphyrin IX. Nat. Prod. Rep. 2003, 20, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Fujita, Y. Regulation and evolution of chlorophyll metabolism. Photochem. Photobiol. Sci. 2008, 7, 1131–1149. [Google Scholar] [CrossRef] [PubMed]
- Stenbaek, A.; Jensen, P.E. Redox regulation of chlorophyll biosynthesis. Phytochemistry 2010, 71, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam, R.; Radhakrishnan, R.; Kim, J.K.; Park, S.U. An update on biosynthesis and regulation of carotenoids in plants. S. Afr. J. Bot. 2020, 1–13. [Google Scholar] [CrossRef]
- Hajiboland, R. Reactive oxygen species and photosynthesis. In Oxidative Damage to Plants; Ahmad, A., Ed.; Academic Press: Cambridge, MA, USA, 2014; Chapter 1; pp. 1–63. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Rad. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Lado, J.; Alós, E.; Manzi, M.; Cronje, P.J.; Gómez-Cadenas, A.; Rodrigo, M.J.; Zacarías, L. Light regulation of carotenoid biosynthesis in the peel of mandarin and sweet orange fruits. Front. Plant Sci. 2019, 10, 1288. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Hormann, H.; Neubauer, C. Chlorophyll fluorescence as diagnostic tool: Basics and some aspects of practical relevance. In Photosynthesis: A Comprehensive Treatise; Raghavendra, A.S., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 320–336. [Google Scholar]
- Cavender-Bares, J.; Bazzaz, F.A. From leaves to ecosystems: Using chlorophyll fluorescence to assess photosynthesis and plant function in ecological studies. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 737–755. [Google Scholar]
- Van Iersel, M.W. Optimizing LED lighting in controlled environment agriculture. In Light Emitting Diodes for Agriculture: Smart Lighting; Gupta, S.D., Ed.; Springer: Singapore, 2017; pp. 59–80. [Google Scholar]
- Pizarro, L.; Stange, C. Light-dependent regulation of carotenoid biosynthesis in plants. Cien. Investig. Agrar. 2009, 36, 143–162. [Google Scholar] [CrossRef]
- Li, C.X.; Chang, S.X.; Khalil-Ur-Rehman, M.; Xu, Z.G.; Tao, J.M. Effect of irradiating the leaf abaxial surface with supplemental light-emitting diode lights on grape photosynthesis. Aust. J. Grape Wine Res. 2017, 23, 58–65. [Google Scholar] [CrossRef]
- Wu, C.; Sun, L.; Lv, Y.; Cui, H.; Wang, X.; Gao, P.; Luan, F. Functional Characterization and in Silico Analysis of Phytoene Synthase Family Genes Responsible for Carotenoid Biosynthesis in Watermelon (Citrullus lanatus L.). Agronomy 2020, 10, 1077. [Google Scholar] [CrossRef]
- Dhingra, A.; Bies, D.H.; Lehner, K.R.; Folta, K.M. Green light adjusts the plastid transcriptome during early photomorphogenic development. Plant Physiol. 2006, 142, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.K.; Holalu, S.V.; Casal, J.J.; and Finlayson, S.A. Abscisic acid regulates axillary bud outgrowth responses to the ratio of red to far-red light. Plant Physiol. 2013, 163, 1047–1058. [Google Scholar] [CrossRef]
- Li, C.X.; Xu, Z.G.; Dong, R.Q.; Chang, S.X.; Wang, L.Z.; Khalil-Ur-Rehman, M.; Tao, J.M. An RNA-seq analysis of grape plantlets grown in vitro reveals different responses to blue, green, red LED light, and white fluorescent light. Front. Plant Sci. 2017, 8, 78. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of blue and red LED light irradiation on β-cryptoxanthin accumulation in the flavedo of citrus fruits. J. Agric. Food Chem. 2012, 60, 197–201. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of the combination of ethylene and red LED light irradiation on carotenoid accumulation and carotenogenic gene expression in the flavedo of citrus fruit. Postharvest Biol. Technol. 2015, 99, 99–104. [Google Scholar] [CrossRef]
- Llorente, B.; Martinez-Garcia, J.F.; Stange, C.; Rodriguez-Concepcion, M. Illuminating colors: Regulation of carotenoid biosynthesis and accumulation by light. Curr. Opin. Plant Biol. 2017, 37, 49–55. [Google Scholar] [CrossRef]
- Tuan, P.A.; Park, C.H.; Park, W.T.; Kim, Y.B.; Kim, Y.J.; Chung, S.O.; Kim, J.K.; Park, S.U. Expression levels of carotenoid biosynthetic genes and carotenoid production in the callus of scutellaria baicalensis exposed to white, blue, and red light-emitting diodes. Appl. Biol. Chem. 2017, 60, 591–596. [Google Scholar] [CrossRef]
- Jeon, Y.M.; Son, K.H.; Kim, S.M.; Oh, M.M. Growth and bioactive compounds as affected by irradiation with various spectrum of light-emitting diode lights in dropwort. Hortic. Environ. Biotechnol. 2017, 58, 467–478. [Google Scholar] [CrossRef]
- Frede, K.; Schreiner, M.; Baldermann, S. Light quality-induced changes of carotenoid composition in pak choi Brassica rapa ssp. chinensis. J. Photochem. Photobiol. B Biol. 2019, 193, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Pola, W.; Sugaya, S.; Photchanachai, S. Color development and phytochemical changes in mature green chili (Capsicum annuum L.) exposed to red and blue light-emitting diodes. J. Agric. Food Chem. 2019, 68, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zang, J.; Xu, Z.; Guo, S.; Jiao, X.; Liu, X.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol. Plant. 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Lobiuc, A.; Vasilache, V.; Oroian, M.; Stoleru, T.; Burducea, M.; Pintilie, O.; Zamfirache, M.M. Blue and Red LED Illumination Improves Growth and Bioactive Compounds Contents in Acyanic and Cyanic Ocimum basilicum L. Microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef]
- Pennisi, G.; Orsini, F.; Blasioli, S.; Cellini, A.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Resource use efficiency of indoor lettuce (Lactuca sativa L.) cultivation as affected by red:blue ratio provided by LED lighting. Sci. Rep. 2019, 9, 14127. [Google Scholar] [CrossRef]
- Ghaffari, Z.; Rahimmalek, M.; Sabzalian, M.R. Variation in the primary and secondary metabolites derived from the isoprenoid pathway in the Perovskia species in response to different wavelengths generated by light emitting diodes (LEDs). Ind. Crop Prod. 2019, 140, 1–8. [Google Scholar] [CrossRef]
- Fang, L.; Ma, Z.; Wang, Q.; Nian, H.; Ma, Q.; Huang, Q.; Mu, Y. Plant growth and photosynthetic characteristics of soybean seedlings under different led lighting quality conditions. J. Plant Growth Regul. 2020, 1–11. [Google Scholar] [CrossRef]
- Chen, X.L.; Guo, W.Z.; Xue, X.Z.; Wang, L.C.; Qiao, X.J. Growth and quality responses of ‘Green Oak Leaf’ lettuce as affected by monochromic or mixed radiation provided by fluorescent lamp (FL) and light-emitting diode (LED). Sci. Hortic. 2014, 172, 168–175. [Google Scholar] [CrossRef]
- Chen, X.L.; Xue, X.Z.; Guo, W.Z.; Wang, L.C.; Qiao, X.J. Growth and nutritional properties of lettuce affected by mixed irradiation of white and supplemental light provided by light-emitting diode. Sci. Hortic. 2016, 200, 111–118. [Google Scholar] [CrossRef]
- Kasim, M.U.; Kasim, R. While continuous white LED lighting increases chlorophyll content (SPAD), green LED light reduces the infection rate of lettuce during storage and shelf-life conditions. J. Food Process. Preserv. 2017, 41, 1–7. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Sakalauskienė, S.; Samuolienė, G.; Jankauskienė, J.; Viršilė, A.; Novičkovas, A.; Sirtautas, R.; Miliauskienė, J.; Vaštakaitė, V.; Dabašinskas, L.; et al. The effects of LED illumination spectra and intensity on carotenoid content in Brassicaceae microgreens. Food Chem. 2015, 173, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Darko, É.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. 2014, 369, 20130243. [Google Scholar] [CrossRef] [PubMed]
- Loconsole, D.; Cocetta, G.; Santoro, P.; Ferrante, A. Optimization of LED lighting and quality evaluation of romaine lettuce grown in an innovative indoor cultivation system. Sustainability 2019, 11, 841. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Sirtautas, R.; Viršilė, A.; Sakalauskaitė, J.; Sakalauskienė, S.; Duchovskis, P. LED illumination affects bioactive compounds in romaine baby leaf lettuce. J. Sci. Food Agric. 2013, 93, 3286–3291. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yang, C.; Zhang, L.; Feng, J.; Xi, W. Effect of light-emitting diodes and ultraviolet irradiation on the soluble sugar, organic acid, and carotenoid content of postharvest sweet oranges (Citrus sinensis (L.) Osbeck). Molecules 2019, 24, 3440. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar] [CrossRef] [PubMed]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Mari, M.; Leoni, O.; Iori, R.; Cembali, T. Antifungal vapour-phase activity of allylisothiocyanate against Penicillium expansum on pears. Plant Pathol. 2002, 51, 231–236. [Google Scholar] [CrossRef]
- Smith, B.J.; Kirkegaard, J.A. In vitro inhibition of soil microorganisms by 2-phenylethyl isothiocyanate. Plant Pathol. 2002, 51, 585–593. [Google Scholar] [CrossRef]
- Wittstock, U.; Kliebenstein, D.J.; Lambrix, V.M.; Reichelt, M.; Gershenzon, J. Glucosinolate hydrolysis and its impact on generalist and specialist insect herbivores. In Integrative Phytochemistry: From Ethnobotany to Molecular Ecology; Romeo, J.T., Ed.; Elsevier: Amsterdam, The Netherlands, 2003; pp. 101–125. [Google Scholar]
- Traka, M.H. Health benefits of glucosinolates. In Glucosinolates, Advances in Botanical Research; Kopriva, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 80, pp. 247–279. [Google Scholar]
- Fahey, J.W.; Olson, M.E.; Stephenson, K.K.; Wade, K.L.; Chodur, G.M.; Odee, D.; Nouman, W.; Massiah, M.; Alt, J.; Egner, P.A.; et al. The diversity of chemoprotective glucosinolates in Moringaceae (Moringa spp.). Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Kim, J.S. Anti-carcinogenic glucosinolates in cruciferous vegetables and their antagonistic effects on prevention of cancers. Molecules 2018, 23, 2983. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Wang, M.; Jian, Y.; Zhang, F.; Zhu, J.; Wang, Q.; Sun, B. Health-promoting phytochemicals and antioxidant capacity in different organs from six varieties of Chinese kale. Sci. Rep. 2019, 9, 20344. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.M.; Juvik, J.A. The role of glucosinolate hydrolysis products from Brassica vegetable consumption in inducing antioxidant activity and reducing cancer incidence. Diseases 2016, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Groenbaek, M.; Jensen, S.; Neugart, S.; Schreiner, M.; Kidmose, U.; Lakkenborg Kristensen, H. Influence of cultivar and fertilizer approach on curly kale (Brassica oleracea L. var. sabellica). 1. Genetic diversity reflected in agronomic characteristics and phytochemical concentration. J. Agric. Food Chem. 2014, 62, 11393–11402. [Google Scholar] [CrossRef]
- Moon, J.; Jeong, M.J.; Lee, S.I.; Lee, J.G.; Hwang, H.; Yu, J.; Kim, Y.-R.; Park, S.W.; Kim, J.A. Effect of LED mixed light conditions on the glucosinolate pathway in brassica rapa. J. Plant Biotechnol. 2015, 42, 245–256. [Google Scholar] [CrossRef]
- Qian, H.; Liu, T.; Deng, M.; Miao, H.; Cai, C.; Shen, W.; Wang, Q. Effects of light quality on main health-promoting compounds and antioxidant capacity of Chinese kale sprouts. Food Chem. 2016, 196, 1232–1238. [Google Scholar] [CrossRef]
- Groenbaek, M.; Tybirk, E.; Kristensen, H.L. Glucosinolate and carotenoid content of white-and yellow-flowering rapeseed grown for human consumption as sprouts and seedlings under light emitting diodes. Eur. Food Res. Technol. 2018, 244, 1121–1131. [Google Scholar] [CrossRef]
- Mithen, R.; Clarke, J.; Lister, C.; Dean, C. Genetics of aliphatic glucosinolates. III. Side chain structure of aliphatic glucosinolates in Arabidopsis thaliana. Heredity 1995, 74, 210–215. [Google Scholar] [CrossRef]
- Giamoustaris, A.; Mithen, R. Genetics of aliphatic glucosinolates IV Side-chain modification in Brassica oleracea. Theor. Appl. Genet. 1996, 93, 1006–1010. [Google Scholar] [CrossRef]
- Augustine, R.; Bisht, N. Biofortification of oilseed Brassica juncea with the anti-cancer compound glucoraphanin by suppressing GSL-ALK gene family. Sci. Rep. 2016, 5, 18005. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J.; Lambrix, V.M.; Reichelt, M.; Gershenzon, J.; Mitchell-Olds, T. Gene duplication in the diversification of secondary metabolism: Tandem 2-oxoglutarate–dependent dioxygenases control glucosinolate biosynthesis in Arabidopsis. Plant Cell 2001, 13, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.S.; Fredericks, D.P.; Griffiths, C.A.; Neale, A.D. The characterisation of AOP2: A gene associated with the biosynthesis of aliphatic alkenyl glucosinolates in Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Kopsell, D.A.; Sams, C.E.; Barickman, T.C.; Morrow, R.C. Sprouting broccoli accumulate higher concentrations of nutritionally important metabolites under narrow-band light-emitting diode lighting. J. Am. Soc. Hortic. Sci. 2014, 139, 469–477. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hortic. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef]
- Abe, K.; Kido, S.; Maeda, T.; Kami, D.; Matsuura, H.; Shimura, H.; Suzuki, T. Glucosinolate profiles in Cardamine fauriei and effect of light quality on glucosinolate concentration. Sci. Hortic. 2015, 189, 12–16. [Google Scholar] [CrossRef]
- Tan, W.K.; Goenadie, V.; Lee, H.W.; Lianga, X.; Loh, C.S.; Ong, C.N.; Tan, H.T.W. Growth and glucosinolate profiles of a common Asian green leafy vegetable, Brassica rapa subsp. Chinensis var. Parachinensis (choy sum), under LED lighting. Sci. Hortic. 2020, 261, 108922. [Google Scholar] [CrossRef]
- Mølmann, J.A.; Hansen, E.; Johansen, T.J. Effects of supplemental LED light quality and reduced growth temperature on swede (Brassica napus L. ssp. rapifera Metzg.) root vegetable development and contents of glucosinolates and sugars. J. Sci. Food Agric 2020. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Agarwal, A. Artificial Lighting System for Plant Growth and Development: Chronological Advancement, Working Principles, and Comparative Assessment. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; Chapter 1; pp. 1–25. [Google Scholar] [CrossRef]
- Both, A.J.; Bugbee, B.; Kubota, C.; Lopez, R.G.; Mitchell, C.; Runkle, E.S.; Wallace, C. Proposed product label for electric lamps used in the plant sciences. HortTechnology 2017, 27, 544–549. [Google Scholar] [CrossRef]
- Nassarawa, S.S.; Abdelshafy, A.M.; Xu, Y.; Li, L.; Luo, Z. Effect of Light-Emitting Diodes (LEDs) on the Quality of Fruits and Vegetables During Postharvest Period: A Review. Food Bioprocess Technol. 2020, 1–27. [Google Scholar] [CrossRef]
- Cavaiuolo, M.; Ferrante, A. Nitrates and glucosinolates as strong determinants of the nutritional quality in rocket leafy salads. Nutrients 2014, 6, 1519–1538. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, C.; Yuk, H.-G.; Khoo, G.H.; Zhou, W. Application of light-emitting diodes in food production, postharvest preservation, and microbiological food safety. Compr. Rev. Food Sci. Food Saf. 2015, 14, 719–740. [Google Scholar] [CrossRef]
- Shi, L.Y.; Cao, S.F.; Chen, W.; Yang, Z.F. Blue light induced anthocyanin accumulation and expression of associated genes in Chinese bayberry fruit. Sci. Hortic. 2014, 179, 98–102. [Google Scholar] [CrossRef]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-light supplementation for enhanced lettuce growth under red-and blue-light-emitting diodes. HortScience 2004, 39, 1617–1622. [Google Scholar] [CrossRef]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Vogelmann, T.C. Plant tissue optics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 231–251. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.N.; Yoshihara, T. Effect of green light wavelength and intensity on photomorphogenesis and photosynthesis in Lactuca sativa. Environ. Exp. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- ANSI/ASABE S640: Quantities and Units of Electromagnetic Radiation for Plants. Photosynthetic Organisms; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2017.
- ANSI/ASABE S642: Recommended Methods for Measurement and Testing of LED Products for Plant Growth and Development; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2018.
- Paucek, I.; Appolloni, E.; Pennisi, G.; Quaini, S.; Gianquinto, G.; Orsini, F. LED Lighting systems for horticulture: Business growth and global distribution. Sustainability 2020, 12, 7516. [Google Scholar] [CrossRef]
- Thomas, A.; Nair, A.S. Experimental study on the effect of junction temperature on power LED’S. Int. J. Curr. Eng. Sci. Res. 2015, 2, 14–19. [Google Scholar]
- U.S. Department of Energy. Life-Cycle Assessment of Energy and Environmental Impacts of LED Lighting Products, Part 2: LED Manufacturing and Performance. 30 January 2012. Available online: www.yole.fr (accessed on 30 January 2019).
- Zheng, Y.J.; Zhang, Y.T.; Liu, H.C.; Li, Y.M.; Liu, Y.L.; Hao, Y.W.; Lei, B.F. Supplemental blue light increases growth and quality of greenhouse pak choi depending on cultivar and supplemental light intensity. J. Integr. Agric. 2018, 17, 2245–2256. [Google Scholar] [CrossRef]
- Jones-Baumgardt, C.; Llewellyn, D.; Ying, Q.; Zheng, Y. Intensity of sole-source light-emitting diodes affects growth, yield, and quality of Brassicaceae microgreens. HortScience 2019, 54, 1168–1174. [Google Scholar] [CrossRef]
- Sipos, L.; Boros, I.F.; Csambalik, L.; Székely, G.; Jung, A.; Balázs, L. Horticultural lighting system optimalization: A review. Sci. Hort. 2020, 273, 109631. [Google Scholar] [CrossRef]
- Braidot, E.; Petrussa, E.; Peresson, C.; Patui, S.; Bertolini, A.; Tubaro, F.; Wählby, U.; Coan, M.; Vianello, A.; Zancani, M. Low-intensity light cycles improve the quality of lamb’s lettuce (Valerianella olitoria [L.] Pollich) during storage at low temperature. Postharvest Biol. Technol. 2014, 90, 15–23. [Google Scholar] [CrossRef]
- Yano, A. Configuration, Function, and Operation of LED Lighting Systems. In LED Lighting for Urban Agriculture; Kozai, T., Fujiwara, K., Runkle, E.S., Eds.; Springer: Singapore, 2016; pp. 403–415. [Google Scholar]
- Hernandez, E.; Timmons, M.B.; Mattson, N.S. Quality, Yield, and Biomass Efficacy of Several Hydroponic Lettuce (Lactuca sativa L.) Cultivars in Response to High Pressure Sodium Lights or Light Emitting Diodes for Greenhouse Supplemental Lighting. Horticulturae 2020, 6, 7. [Google Scholar] [CrossRef]
- Alrifai, O.; Hao, X.; Liu, R.; Lu, Z.; Marcone, M.F.; Tsao, R. Amber, red and blue LEDs modulate phenolic contents and antioxidant activities in eight Cruciferous microgreens. J. Food Bioact. 2020, 11. [Google Scholar] [CrossRef]
- Purschwitz, J.; Müller, S.; Kastner, C.; Fischer, R. Seeing the rainbow: Light sensing in fungi. Curr. Opin. Microbiol. 2006, 9, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Gupta, A.; Huang, Y.Y.; Yin, R.; Murray, C.K.; Vrahas, M.S.; Sherwood, M.E.; Tegos, G.P.; Hamblin, M.R. Blue light rescues mice from potentially Pseudomonas aeruginosa burn infection: Efficacy, safety, and mechanism of action. Antimicrob. Agents Chemother. 2013, 57, 1238–1245. [Google Scholar] [CrossRef]
- Huang, L.; Xuan, Y.; Koide, Y.; Zhiyentayev, T.; Sanaka, M.; Hamblin, M.R. Type I and Type II mechanisms of antimicrobial photodynamic therapy. An in vitro study on gram-negative and gram-positive bacteria. Lasers Surg. Med. 2012, 44, 490–499. [Google Scholar] [CrossRef]
- Endarko, E.; Maclean, M.; Timoshkin, I.V.; MacGregor, S.J.; Anderson, J.G. High-intensity 405 nm light inactivation of Listeria monocytogenes. Photochem. Photobiol. 2012, 88, 1280–1286. [Google Scholar] [CrossRef]
- Imada, K.; Tanaka, S.; Ibaraki, Y.; Yoshimura, K.; Ito, S. Antifungal effect of 405-nm light on Botrytis cinerea. Lett. Appl. Microbiol. 2014, 59, 670–676. [Google Scholar] [CrossRef]
- Maclean, M.; McKenzie, K.; Anderson, J.G.; Gettinby, G.; MacGregor, S.J. 405 nm light technology for the inactivation of pathogens and its potential role for environmental disinfection and infection control. J. Hospital Infect. 2014, 88, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ghate, V.; Kim, M.J.; Zhou, W.; Khoo, G.H.; Yuk, H.G. Inactivation and changes in metabolic profile of selected foodborne bacteria by 460 nm LED illumination. Food Microbiol. 2017, 63, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Ballario, P.; Vittorioso, P.; Magrelli, A.; Talora, C.; Cabibbo, A.; Macino, G. White collar-1, a central regulator of blue light responses in Neurospora, is a zinc finger protein. EMBO J. 1996, 15, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Ballario, P.; Macino, G. White-collar proteins: PASsing the light signal in Neurospora crassa. Trends Microbiol. 1997, 5, 458–462. [Google Scholar] [CrossRef]
- Alferez, F.; Liao, H.L.; Burns, J.K. Blue light alters infection by Penicillium digitatum in tangerines. Postharvest Biol. Technol. 2012, 63, 11–15. [Google Scholar] [CrossRef]
- Liao, H.L.; Alferez, F.; Burns, J.K. Assessment of blue light treatments on citrus postharvest diseases. Postharvest Biol. Technol. 2013, 81, 81–88. [Google Scholar] [CrossRef]
- Lafuente, M.T.; Alferez, F. Effect of LED Blue Light on Penicillium digitatum and Penicillium italicum strains. Photochem. Photobiol. 2015, 91, 1412–1421. [Google Scholar] [CrossRef]
- Yamaga, I.; Takahashi, T.; Ishii, K.; Kato, M.; Kobayashi, Y. Antifungal Effect of Blue LED Irradiation on the Blue Mold, Penicillium italicum, in Satsuma Mandarin Fruits. Hortic. Res. 2015, 14, 83–87. [Google Scholar] [CrossRef][Green Version]
- D’Hallewin, G.; Schirra, M.; Manueddu, E.; Piga, A.; Ben Yehoshua, S. Scoparone and scopoletin accumulation and ultraviolet-C induced resistance to postharvest decay in oranges as influenced by harvest date. J. Am. Soc. Hortic. Sci. 1999, 124, 702–707. [Google Scholar] [CrossRef]
- Marcos, J.F.; González-Candelas, L.; Zacarías, L. Involvement of ethylene biosynthesis and perception in the susceptibility of citrus fruit to Penicillium digitatum infection and the accumulation of defense-related mRNAs. J. Exp. Bot. 2005, 56, 2183–2193. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Gonzalez-Candelas, L.; Alamar, S.; Sanchez-Torres, P.; Zacarias, L.; Marcos, J. A transcriptomic approach highlights induction of secondary metabolism in citrus fruit in response to Penicillium digitatum infection. BMC Plant Biol. 2010, 10, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.R.; Lafuente, M.T.; de Vos, R.C.H.; Bovy, A.G.; González-Candelas, L. Citrus phenylpropanoids and defense against pathogens. Part I: Metabolic profiling in elicited fruits. Food Chem. 2013, 136, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.R.; Lafuente, M.T. LED Blue Light-induced changes in phenolics and ethylene in citrus fruit: Implication in elicited resistance against Penicillium digitatum infection. Food Chem. 2017, 218, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.M.; Lee, Y.H. Effect of light quality on Bacillus amyloliquefaciens JBC36 and its biocontrol efficacy. Biol. Control 2013, 64, 203–210. [Google Scholar] [CrossRef]
- Logrieco, A.; Bottalico, A.; Mulè, G.; Moretti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some Mediterranean crops. Eur. J. Plant Pathol. 2003, 109, 645–667. [Google Scholar] [CrossRef]
- Yu, J.J.; Keller, N. Regulation of secondary metabolism in filamentous fungi. Annu. Rev. Phytopathol. 2005, 43, 437–458. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Ruefer, C.; Raupp, F.; Bruchmann, A.; Perrone, G.; Geisen, R. Influence of light on food relevant fungi with emphasis on ochratoxin producing species. Int. J. Food Microbiol. 2011, 145, 229–237. [Google Scholar] [CrossRef]
- Fanelli, F.; Schmidt-Heydt, M.; Haidukowski, M.; Geisen, R.; Logrieco, A.; Mulè, G. Influence of light on growth, fumonisin biosynthesis and FUM1 gene expression by Fusarium proliferatum. Int. J. Food Microbiol. 2012, 153, 148–153. [Google Scholar] [CrossRef]
- Ntagkas, N.; de Vos, R.C.; Woltering, E.J.; Nicole, C.; Labrie, C.; Marcelis, L.F. Modulation of the Tomato Fruit Metabolome by LED Light. Metabolites 2020, 10, 266. [Google Scholar] [CrossRef]
- Loi, M.; De Leonardis, S.; Mulè, G.; Logrieco, A.F.; Paciolla, C. A Novel and Potentially Multifaceted Dehydroascorbate Reductase Increasing the Antioxidant Systems is Induced by Beauvericin in Tomato. Antioxidants 2020, 9, 435. [Google Scholar] [CrossRef]
- Loi, M.; Paciolla, C.; Logrieco, A.F.; Mulè, G. Plant Bioactive Compounds in Pre-and Postharvest Management for Aflatoxins Reduction. Front. Microbiol. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Bashir, T.; Ghosh, R.; Lee, S.K.; Bae, H. An overview of LEDs’ effects on the production of bioactive compounds and crop quality. Molecules 2017, 22, 1420. [Google Scholar] [CrossRef] [PubMed]
- Alsanius, B.W.; Karlsson, M.; Rosberg, A.K.; Dorais, M.; Naznin, M.T.; Khalil, S.; Bergstrand, K.J. Light and Microbial Lifestyle: The Impact of Light Quality on Plant–Microbe Interactions in Horticultural Production Systems—A Review. Horticulturae 2019, 5, 41. [Google Scholar] [CrossRef]
- Nadalini, S.; Zucchi, P.; Andreotti, C. Effects of blue and red LED lights on soilless cultivated strawberry growth performances and fruit quality. Eur. J. Hortic. Sci. 2017, 82, 12–20. [Google Scholar] [CrossRef]
- Díaz-Galián, M.V.; Torres, M.; Sanchez-Pagán, J.D.; Navarro, P.J.; Weiss, J.; Egea-Cortines, M. Enhancement of strawberry production and fruit quality by blue and red LED lights in research and commercial greenhouses. S. Afr. J. Bot. 2020. [Google Scholar] [CrossRef]
- Lakhiar, I.A.; Gao, J.; Syed, T.N.; Chandio, F.A.; Buttar, N.A. Modern plant cultivation technologies in agriculture under controlled environment: A review on aeroponics. J. Plant Interact. 2018, 13, 338–352. [Google Scholar] [CrossRef]
- Kim, H.J.; Yang, T.; Choi, S.; Wang, Y.J.; Lin, M.Y.; Liceaga, A.M. Supplemental intracanopy far-red radiation to red LED light improves fruit quality attributes of greenhouse tomatoes. Sci. Hortic. 2020, 261, 108985. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | Formula | Wavelengths [nm] | Light Color | Forward Voltage [V] |
|---|---|---|---|---|
| Gallium-Phosphide Aluminium-Gallium-Arsenide Gallium-Arsenide-Phosphide Aluminium-Gallium-Indium-Phosphide | GaP AlGaAs GaAsP AlGaInP | 610 < < 770 | Red | 1.6–2.0 |
| Gallium-Phosphide Gallium-Arsenide-Phosphide Aluminium-Gallium-Indium-Phosphide | GaP GaAsP AlGaInP | 590 < < 610 | Orange | 2.0–2.1 |
| Gallium-Arsenide-Phosphide Gallium-Phosphide | GaAsP GaP | 570 < < 590 | Yellow | 2.1–2.2 |
| Gallium-Phosphide Aluminium-Gallium-Phosphide Aluminium-Gallium-Indium-Phosphide | GaP AlGaP AlGaInP | 500 < < 570 | Green | 1.9–4 |
| Silicon carbide as substrate Zinc Sulfide | SiC ZnS | 450 < < 500 | Blue | 2.4–3.7 |
| Gallium-Nitride Indium-Gallium-Nitride | GaN InGaN | 400 < < 450 | Violet | 2.7–4.0 |
| Blue diode with yellow phosphor | Broad spectrum | White | 3.5 |
| Plant Species | LED (Wavelength, Intensity), Photoperiod, and Experimental Parameters | Type of Application | Effect | Reference |
|---|---|---|---|---|
| Satsuma mandarin (Citrus unshiu Marc.), Valencia orange (C. sinensis Osbeck), Lisbon lemon (C. limon Burm.f.) | BL (470 nm, 50 μmol m−2·s−1) + RL (660 nm, 50 μmol m−2·s−1) for 4 weeks; BL (50 and 100 μmol m−2·s−1) for 4 weeks; continuous and pulsed BL (100 μmol m−2·s−1) for 4 weeks | Postharvest | BL increased Vit C content RL did not affect Vit C content | [72] |
| Brassica rapa ssp. Chinensis | RL (35 μmol m−2·s−1) for 8 h/per day at 20 °C | Postharvest | RL increased Vit C, RL delayed senescence, BL no effect | [73] |
| Brassica oleracea L. Var. Italica | BL (470 nm, 50 μmol m−2·s−1) at 20 °C and RH > 95%, Modified WL (RL 660 nm > BL) | Postharvest | RL delayed senescence, modified WL (RL>BL) increased Vit C, BL no effect | [55] |
| Fragaria ananassa Duch. Cv. Fengguang | BL (470 nm, 40 μmol m−2·s−1) for 12 days at 5 °C, RH 80–90%. | Postharvest | BL increased Vit C | [75] |
| Lactuca sativa L. Cv. Lvdie | WL (80 RL + 20 BL, 250 μmol m–2·s–1) | Growth | 5 GL + 80 RL + 15 BL decreased Vit C | [76] |
| Lettuce plants | 25 RL + 75 BL (200 μmol m–2·s–1) 12 days under continuous irradiation); 23 °C (RH 50–60%) | Growth | 25 RL + 75 BL increased Vit C | [77] |
| Brassica campestris L | BL + RL (B/R = 1:8) (80 μmol m–2s–1) 12-h photoperiod; 24–26°C; RH 40–50% | Seedlings | BL + RL increased Vit C | [78] |
| Lentil (Lens esculenta Moenh), wheat (Triticum aestivum L.), radish (Raphanus sativus L.) | GL (510 nm,100 μmol m−2·s−1; 12-h photoperiod; 27 °C) for lentil and wheat; GL + Amber Light for radish | Postharvest | GL increased Vit C (lentil and wheat); GL + Amber Light increased Vit C (radish) | [81] |
| Solanum lycopersicum | WL (263 μmol m−2·s−1) for 13 days nm | Postharvest | WL increased Vit C (plus 32%) | [82] |
| Lactuca sativa L. Var. Capitata | BL (480 nm), GL (520 nm), RL (630 nm), WL (455 and 570 nm), 400 μmol m−2·s−1 | Postharvest | supplemented GL either 4 days or 43 days enhanced Vit C inside the crisp head lettuce | [83] |
| Apple (Malus domestica ‘Granny Smith’), red tomato (Solanum lycopersicum L.), bell pepper (Capsicum annuum) | Constant YL (590 nm, 1.81 W/m2) 10 °C for 7 days | Postharvest | YL not significantly increased Vit C | [84] |
| Brassica rapa ‘Dongdori’ | WL, GL (524 nm), BL (436 nm), RL (664 nm) stored at 4–5 °C for 18 days | Postharvest | BL increased Vit C with BL > WL > GL > RL > non-irradiate | [85] |
| Plant Species | LED (Wavelength, Intensity), Photoperiod, and Experimental Parameters | Type of Application | Effect | Reference |
|---|---|---|---|---|
| Broccoli (Brassica oleracea L. Var. Italica) | BL (467 nm, 21 µmol m−2·s−1), GL (522 nm,24 µmol m−2·s−1), YL (587 nm, 27 µmol m−2·s−1) RL (625 nm, 66 µmol m−2·s−1) WL (3000 K color temperature, 40 µmol m−2·s−1); continuous irradiation at 4 °C | Postharvest | RL > YL > BL increased the TPC throughout the storage period | [56] |
| Brussels sprouts (Brassica oleracea var. gemmifera) | UV-B (300 nm, 0.02 µmol m−2·s−1), UV-A (365 nm, 14 µmol m−2·s−1), BL (470 nm, 11 µmol m−2·s−1), RL (660 nm, 2 µmol m−2·s−1); 16-h photoperiod | Sprouting | Compound specific effects | [91] |
| Buckwheat sprouts (Fagopyrum esculentum Moench) | BL (460nm), RL (625nm), FL (35 µmol m−2·s−1); 16 h of light; supplement and 8 h of dark per day | Sprouting | BL > RL > FL increasedTPC, TFC, and antioxidant capacity using ABTS radical cation assay. BL increased orientin, isoorientin, vitexin, isovitexin, rutin, and quercetin-3-O-robinobioside | [92] |
| Basil (Ocimum basilicum), arugula (Eruca sativa), and bloody dock (Rumex sanguineus) | BL (450 nm, 300 µmol m−2·s−1), BVL (440–420 nm, 300 µmol m−2·s−1), HPS (600nm, 400 W); | Growth | BL and BVL increased the phenolic acid content in basil, BV the flavonoids in arugula. No increase was registered in bloody dock with either BL or BV | [93] |
| Cherry tomato seedlings (Solanum lycopersicum L. ‘Cuty’) | RL (655 nm, 200 µmol m−2·s−1), BL (456 nm, 200 µmol m−2·s−1), GL (518 nm, 200 µmol m−2·s−1), WL (456 nm, 200 µmol m−2·s−1) | Growth | BL > WL > RL > GL increased the TPC and TFC | [94] |
| Tea (Camellia sinensis cv. Fujian Shuixian) | BL (450nm) at low (50 µmol m−2·s−1) medium (100 µmol m−2·s−1) and high (200 µmol m−2·s−1) intensity, WL (100 µmol m−2·s−1); 12-h photoperiod, 14 days | Growth | High-intensity BL down-regulated several genes involved in flavonoid biosynthesis; no effects elicited by low- and medium-intensity BL. BL increased the levels of 3′,5,6-trihydroxy-3,4′,7,8-tetramethoxyflavone 3-glucoside, galangin 3-[galactosyl-(1-4)-rhamnoside], and neocarthamin, and reduced the levels of 3-(2-caffeoylsophoroside) 7-glucoside, quercetin 3-(4‘-acetylrhamnoside) 7-rhamnoside, and spinacetin 3-(2‘- feruloylgentiobioside | [95] |
| Navel oranges (Citrus sinensis Osbeck) | UVC (10–280 nm, 100 µmol m−2·s−1), UVB (270–315 nm, 100 µmol m−2·s−1), UVA (315–400 nm, 100 µmol m−2·s−1), BL (470 nm, 200 µmol m−2·s−1), RL (660 nm, 150 µmol m−2·s−1), WL (100 µmol m−2·s−1); Continuous irradiation for 6 days; sampling after 0, 6, and 15 days | Postharvest | BL and RL retained more PC during the irradiation period, while WL, UVB, and UVC stimulated their accumulation after the irradiation period. RL and BL maintained the levels of diosmin, diosmetin 6,8-di-C-glucoside, hesperidin, didymin, neoeriocitrin, and narirutin; all Uvs increased narirutin, neoeriocitrin, and didymin | [98] |
| Sweet pepper (Capsicum annuum L cv. California Wonder, King of the North, Citrine F1 Hybrid) | RL (660 nm, 150 µmol m−2·s−1) BL (450 nm, 100 µmol m−2·s−1); 8-h photoperiod | Postharvest | RLwas more effective than BL in increasing the TFC, and PAL activity | [101] |
| Okra (Abelmoschus esculentus L.) | RL (630 nm, 17.28 W/m2), BL (470 nm, 17.28 W/m2), GL (560 nm, 17.28 W/m2); 8-h photoperiod | Postharvest | WL and BL increased the TFC content and the activity of key enzymes involved in the biosynthesis of phenolics | [102] |
| Bananas (Musa acuminata cv. Berangan) | BL (464–474 nm, 3920 µmol m−2·s−1), GL (515–525 nm, 4340 µmol m−2·s−1) equivalent to photosynthetic intensity of 100 W m−2, RL (617–627 nm, 5200 µmol m−2·s−1); continuous irradiation for 8 days | Postharvest | BL was more effective than RL and GL in increasing in TPC | [107] |
| Broccoli (Brassica oleracea L. Var. Italica cv. Chaoda No. 1) | WFL (300–700nm, (12–13 µmol m−2·s−1), GL (520 nm, 12–13 µmol m−2·s−1); 12-h photoperiod | Postharvest | GL increased the TPC | [108] |
| Red leaf lettuce (Lactuca sativa L., cv. Sunmang) and green leaf lettuce (Lactuca sativa L., cv. Grand Rapid TBR) | Various combinations of BL (456 nm) and RL (655 nm) BL: RL = 0:100%, 13%:87%, 26%:74%, 35%:65%, 47%:53%, 59%:41%, (171 µmol m−2·s−1), 12-h photoperiod | Postharvest | Intermediate BL/RL ratios (35%:65% > 47%:53% > 59%:41% in cv. Sunmang) stimulated the accumulation of TPC | [110] |
| Pea sprouts (Pisum sativum L.) | RL (635 nm), BL (460 nm), YL (585 nm), WL (30 µmol m−2·s−1); 12-h photoperiod | Sprouting | BL was more effective than WL, RL, and YL in increasingd the TPC and TFC with respect to the dark. BL increased the content of chlorogenic acid, p-hydroxybenzoic acid, caffeic acid, gallic acid, p-coumaric acid, ferulic acid, rutin, and resveratrol; WL increased the content of chlorogenic acid, p-hydroxybenzoic acid, caffeic acid, gallic acid, ferulic acid, rutin, and resveratrol; RL increased the content of p-hydroxybenzoic acid, caffeic acid, p-coumaric acid, ferulic acid, and rutin; YL increased p-hydroxybenzoic acid, caffeic acid, gallic acid, p-coumaric acid, ferulic acid, rutin, and resveratrol. | [112] |
| Canola sprouts (Brassica napus L.) | WL (380 nm, 50 µmol m−2·s−1), BL (470 nm, 50 µmol m−2·s−1), RL (660 nm, 50 µmol m−2·s−1), BL + RL; 16-h photoperiod | Sprouting | BL was more effective than WL, BL + RL, and RL in increasing the content of phenolic acids, flavonoids, and catechin. BL increased the content of benzoic acid, (+)-catechin, caffeic acid, and (−)-epicatechin; WL increased the contents of rutin. | [113] |
| Lettuce (Lactuca sativa L. Cv. ‘Grizzly’ | WL (380–760 nm, 300 µmol m−2·s−1), RL (650–665 nm, 300 µmol m−2·s−1), BL (460–475 nm, 300 µmol m−2·s−1) or RL + BL (70%:30%), 14-h photoperiod | Growth | WL > BL increased the TPC | [96] |
| Longan (Dimocarpus longan Lour.) | BL (457 nm, 32 µmol m−2·s−1), GL (515 nm, 32 µmol m−2·s−1), WL 32 µmol m−2·s−1), and RL (660 nm, 32 µmol m−2·s−1) 12-h photoperiod; different BL intensities (16, 32,64, 128, and 256 µmol m−2·s−1), 12-h photoperiod; BL at 32 µmol m-2·s-1 with different photoperiods: 8, 12, 16, 20, and 24 h | Growth | BL was more effective than GL and WL in increasing the TFC; 32 µmol m−2·s−1 was the most effective intensity to increase the TFC; 12-h photoperiod was the most effective in increasing TFC; BL promoted the accumulation of epicatechin, but inhibitedthe synthesis of rutin. | [99] |
| Sarcandra Herb (Sarcandra glabra) | WL (380–760 nm, 80 µmol m−2·s−1), RL, 656 nm, 80 µmol m−2·s−1), BL, 450 nm80 µmol m−2·s−1); 16-h photoperiod | Growth | BL increased the levels of cinnamic acid, 4-coumaric acid, chalcone, naringenin quercitin,kaempferol, and rutin, while it reduced the caffeic acid content. BL increased the expression of of key enzymes involved in the biosynthesis of phenolics (PAL, FLS) | [97] |
| Van’sweet cherry (Prunus avium L.) | UV-B (310nm, 0.046 W m−2), BL (444 nm, 1 W m−2), WBG composed by BL (470nm), GL (520 nm) and WL with a total 3.6 radiant flux W m2 | Postharvest | BL increased the content of cyanidin 3-O-glucoside, cyanidin 3-O-rutinoside; BL was more effective than WBG in increasing PAL activity | [100] |
| Plant Species | LED (Wavelength, Intensity), Photoperiod, and Experimental Parameters | Type of Application | Effect | Reference |
|---|---|---|---|---|
| Chinese cabbage seeds (Brassica campestris L. Teaiqing) | YL (590 nm, 150 µmol m−2·s−1), GL (520 nm, 150 µmol m−2·s−1), RL (658 nm, 150 µmol m−2·s−1), BL (460 nm, 150 µmol m−2·s−1), RL + BL (6:1 ratio); 12-h photoperiod | Growth | All LEDs except RL + BL led to a decrease in total chl and carotenoids | [142] |
| Lettuce (Lactuca sativa L. cv. ‘Grizzly’ | WL (380–760 nm, 300 µmol m−2·s−1), RL (650–665 nm, 300 µmol m−2·s−1), BL (460–475 nm, 300 µmol m−2·s−1) orRL + BL (70%:30%), 14-h photoperiod | Growth | RL + BL > RL > BL increased the total chl; RL+BL increased the total carotenoids. All LEDs improved the photosynthetic rate | [96] |
| Grape (Vitis vinifera L. × V. labrusca L cv. Summer Black) | RL (660nm, 100 µmol m−2·s−1), BL (460nm, 100 µmol m−2·s−1), RL + BL (660 + 630 + 460 nm) | Growth | BL increased the photosynthetic rate | [130] |
| Satsuma mandarin (Citrus unshiu Marc.) | BL (470 nm, 50 µmol m−2·s−1), RL (660 nm) 50 µmol m−2·s−1); continuous irradiation for 6 days at 20°C | Postharvest | BL increased the expression of carotenoid biosynthetic genes and carotenoids content | [135] |
| Pak choi (Brassica rapa ssp. chinensis cv. Black Behi) | BL (453nm, 80 µmol m−2·s−1), RL (633nm, 80 µmol m−2·s−1), WL (404–789nm, 80 µmol m−2·s−1); 12-h photoperiod | Growth | WL > RL > BL increased the carotenoid and chlorophyll contents | [140] |
| Sweet oranges (Citrus sinensis (L.) Osbeck) | RL (660 nm, 150 µmol m−2·s−1), BL (470 nm, 200 µmol m−2·s−1), WL (100 µmol m−2·s−1), UVA (315–400 nm, 100 µmol m−2·s−1), UVB (270–315 nm, 100 µmol m−2·s−1), UVC (100–280 nm, 100 µmol m−2·s−1); continuous irradiation for 6 days | Postharvest | Dark > RL increased the carotenoid content | [154] |
| Mustard (Brassica juncea L., ‘Red Lion’), beet (Beta vulgaris L., ‘Bulls Blood’), and parsley (Petroselinum crispum Mill., ‘Plain Leaved or French) | Different combinations of BL (447 nm, 300 µmol m−2·s−1) RL (638nm, 300 µmol m−2·s−1), FRL (731 nm, 300 µmol m−2·s−1); 16-h photoperiod | Growth | BL led to the accumulation of chlorophylls, carotenoids, carotenes, lutein, violaxanthin, and zeathin | [109] |
| Broccoli (Brassica oleracea L. var. italica) | BL (467 nm, 21 µmol m−2·s−1), GL (522 nm, 24 µmol m−2·s−1), YL (587 nm, 27 µmol m−2·s−1), RL (625 nm, 66 µmol m−2·s−1), WL (3000 K color temperature, 40 µmol m2·s−1); continuous irradiation at 4 °C | Postharvest | GL increased the chlorophyll content; no effect on carotenoids elicited by all LEDs | [56] |
| Green Oak Leaf lettuce (Lactuca sativa var. crispa) | WFL (400–700 nm, 133 µmol m−2·s−1), BL (460 nm, 133 µmol m−2·s−1), RL (630 nm, 133 µmol m−2·s−1), FBL (WFL + BL), FRL (WFL + RL); 14-h photoperiod | Growth | FLR > FLB > RB increased total chlorophyll content and total carotenoids | [147] |
| Green Oak Leaf lettuce (Lactuca sativa var. crispa) | WFL + FRL (850 nm, 135 µmol m−2·s−1), WFL + RL (660 nm, 135 µmol m−2·s−1), WFL + YL (596 nm, 135 µmol m−2·s−1), WFL + GL (522 nm, 135 µmol m−2·s−1), WFL + BL (450 nm, 135 µmol m−2·s−1) | Growth | WFL + RL and WFL + BL increased the total chlorophyll and carotenoid content | [148] |
| Baikal skullcap (Scutellaria baicalensis) | RL (660 nm, 50 µmol m−2·s−1), BL (470 nm, 50 µmol m−2·s−1), WL (380 nm, 50 µmol m−2·s−1); 16-h photoperiod | Growth | BL > RL > WL increased the expression of carotenoids photosynthetic genes; BL increased the zeaxanthin, β-carotene, and 9-cis-β-carotene content | [138] |
| Green Chili (Capsicum annuum L.) | RL (660 nm, 50 µmol m−2·s−1), BL (470 nm, 50 µmol m−2·s−1), continuous irradiation for 3 days | Postharvest | RL induced the expression of lycopene-β-cyclase (Lcyb), β-carotene hydroxylase (CrtZ), and capsanthin/capsolubin synthase (Ccs); BL induced the expression of PS; RL > BL increased the carotene, free-capsanthin, and total carotenoid contents | [141] |
| Plant Species | LED (Wavelength, Intensity), Photoperiod, and Experimental Parameters | Type of Application | Effect | Reference |
|---|---|---|---|---|
| Broccoli (Brassica oleracea L. var. italica cv. Chaoda No. 1) | WFL (300–700 nm, (12–13 (µmol m−2·s−1)GL (520 nm, 12–13 µmol m−2·s−1) 12-h photoperiod | Postharvest | GL increased GLSs contents | [108] |
| Sprouting Canola (Brassica napus L.) | WL (380 nm, 50 µmol m−2·s−1) BL (470 nm, 50 µmol m−2·s−1) RL (660 nm, 50 µmol m−2·s−1) BL + RL; 16-h photoperiod | Growth | BL + RL determined the lowest levels of total GLSs. RL increased the level of sinigrin, glucobrassicin, and 4-methoxy glucobrassicin, while BL increased the level of glucoraphanin. WL and BL increased the level of glucoalyssin and gluconapin | [113] |
| kale sprouts (Brassica oleacea var. alboglabra Bailey) | WL (440-660 nm, 30 µmol m−2·s−1) RL (660 nm, 30 µmol m−2·s−1) BL (470 nm, 30 µmol m−2·s−1). 16-h photoperiod | Growth | No effects elicited by WL and RL; BL inhibited the accumulation of GLSs in shoots. | [168] |
| white- and yellow-flowering rapeseed (Brassica napus L.) | HB (31.7% blue/66.3% red, 121 ± 7 µmol m−2·s−1) LB (14.8% blue/81.3% red, 121 ± 7 µmol m−2·s−1)14-h photoperiod | Growth | HB light decreased the contents of progoitrin. BL/RL light ratios slightly affect GLSs content. | [169] |
| Broccoli (Brassica oleracea var. italica) | (A) FL (B) BL/RL (5% 447 nm/95% 627 nm) (C) BL/RL/GL (5%/85%/10% 530 nm) (D) BL/RL (20%/80%) (E) BL/RL/GL (20%/70%/10%). 250 µmol m−2·s−1 ; 16-h photoperiod | Growth | D > C > B > E > A increased the level of aliphatic, indoles, and total GLS. | [175] |
| Broccoli (Brassica oleracea var. italica) | BL + RL (12% 470 nm/88% 627 nm; 350 µmol m−2·s−1); BL (470 nm; 41 ± 2 µmol m−2·s−1). 24-h photoperiod | Growth | BL increased the level of glucoraphenin, epiprogoitrin, and aliphatic GLSs, while indole GLSs were not impacted by the LED lighting treatment. | [176] |
| Cardamine fauriei | RL (660 nm), BL (445 nm), GL (520 nm) RL + BL (1:1), 75 µmol m−2·s−1; 16-h photoperiod | Growth | BL increased accumulation of aliphatic GSLs and decreased accumulation of indolic GSL | [177] |
| Choy sum (Brassica rapa subsp. chinensis var. parachinensis) | RB: 160 μmol m−2·s−1 and 80 μmol m−2·s−1; WL: 160 W and 80 W; BL (450 nm) RL (660 nm). 12-h photoperiod | Growth | No significant difference in the total GSL content was observed in response to the various LED light treatments within the same growth stage. The changes were more pronounced across growth stages. | [178] |
| (Brassica napus L. ssp. rapifera Metzg) | RL (660 nm), FR (740 nm), RL/FRL (1:1), BL (460 nm); 24-h photoperiod;10 μmol m−2·s−1.These treatments were given in combination with temperature regimes of either constant 15 °C (100 day) or 15 °C (65 day) followed by 9 °C (66–100 day) | Growth | Total GLS concentrations were not different among treatments, progoitrin was present in highest concentration under LEDs containing far-red light, and in lower concentration at 9 °C compared to 15 °C | [179] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loi, M.; Villani, A.; Paciolla, F.; Mulè, G.; Paciolla, C. Challenges and Opportunities of Light-Emitting Diode (LED) as Key to Modulate Antioxidant Compounds in Plants. A Review. Antioxidants 2021, 10, 42. https://doi.org/10.3390/antiox10010042
Loi M, Villani A, Paciolla F, Mulè G, Paciolla C. Challenges and Opportunities of Light-Emitting Diode (LED) as Key to Modulate Antioxidant Compounds in Plants. A Review. Antioxidants. 2021; 10(1):42. https://doi.org/10.3390/antiox10010042
Chicago/Turabian StyleLoi, Martina, Alessandra Villani, Francesco Paciolla, Giuseppina Mulè, and Costantino Paciolla. 2021. "Challenges and Opportunities of Light-Emitting Diode (LED) as Key to Modulate Antioxidant Compounds in Plants. A Review" Antioxidants 10, no. 1: 42. https://doi.org/10.3390/antiox10010042
APA StyleLoi, M., Villani, A., Paciolla, F., Mulè, G., & Paciolla, C. (2021). Challenges and Opportunities of Light-Emitting Diode (LED) as Key to Modulate Antioxidant Compounds in Plants. A Review. Antioxidants, 10(1), 42. https://doi.org/10.3390/antiox10010042