Quercetin and Egg Metallome

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Determination of Egg Trace and Macro Elements
2.3. Statistical Analysis
3. Results
3.1. Concentration of Egg Elements
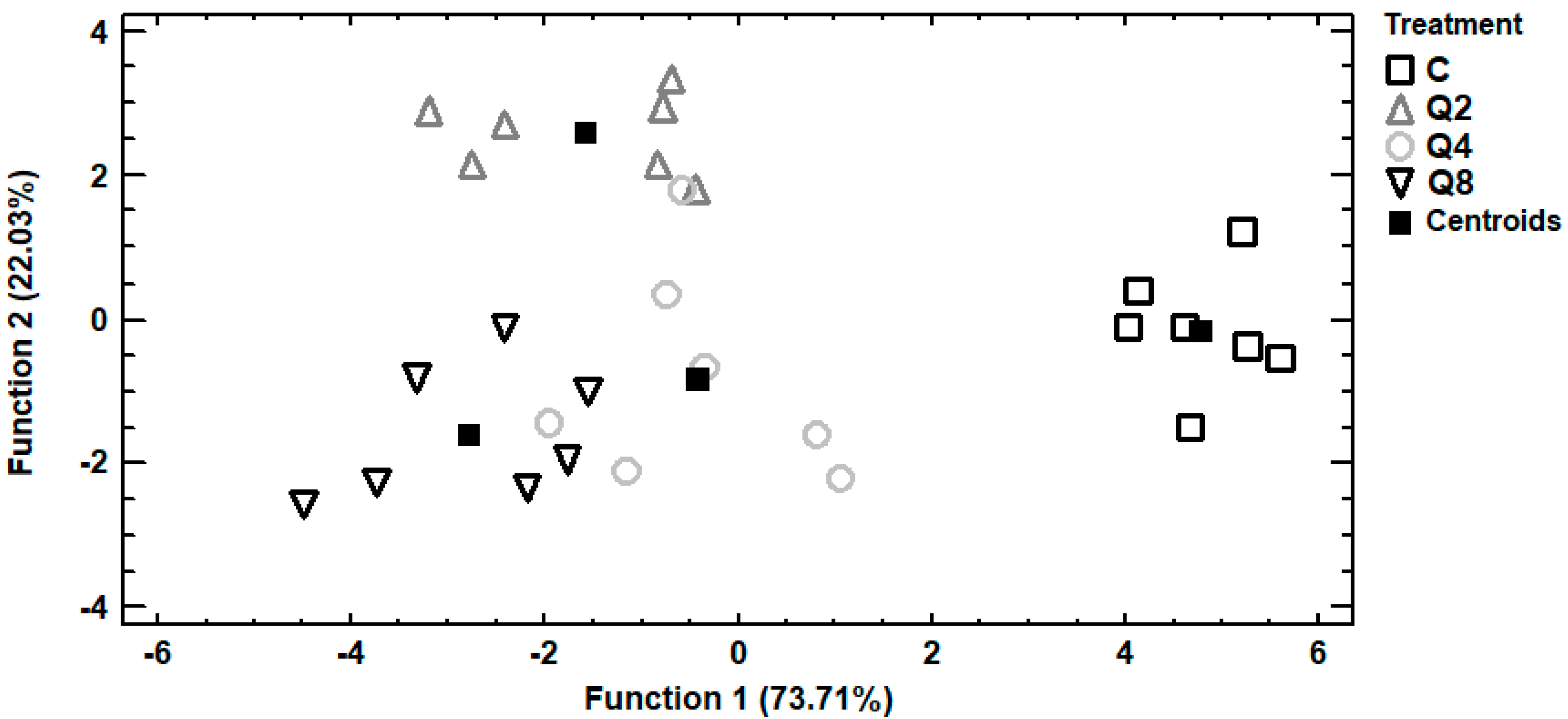
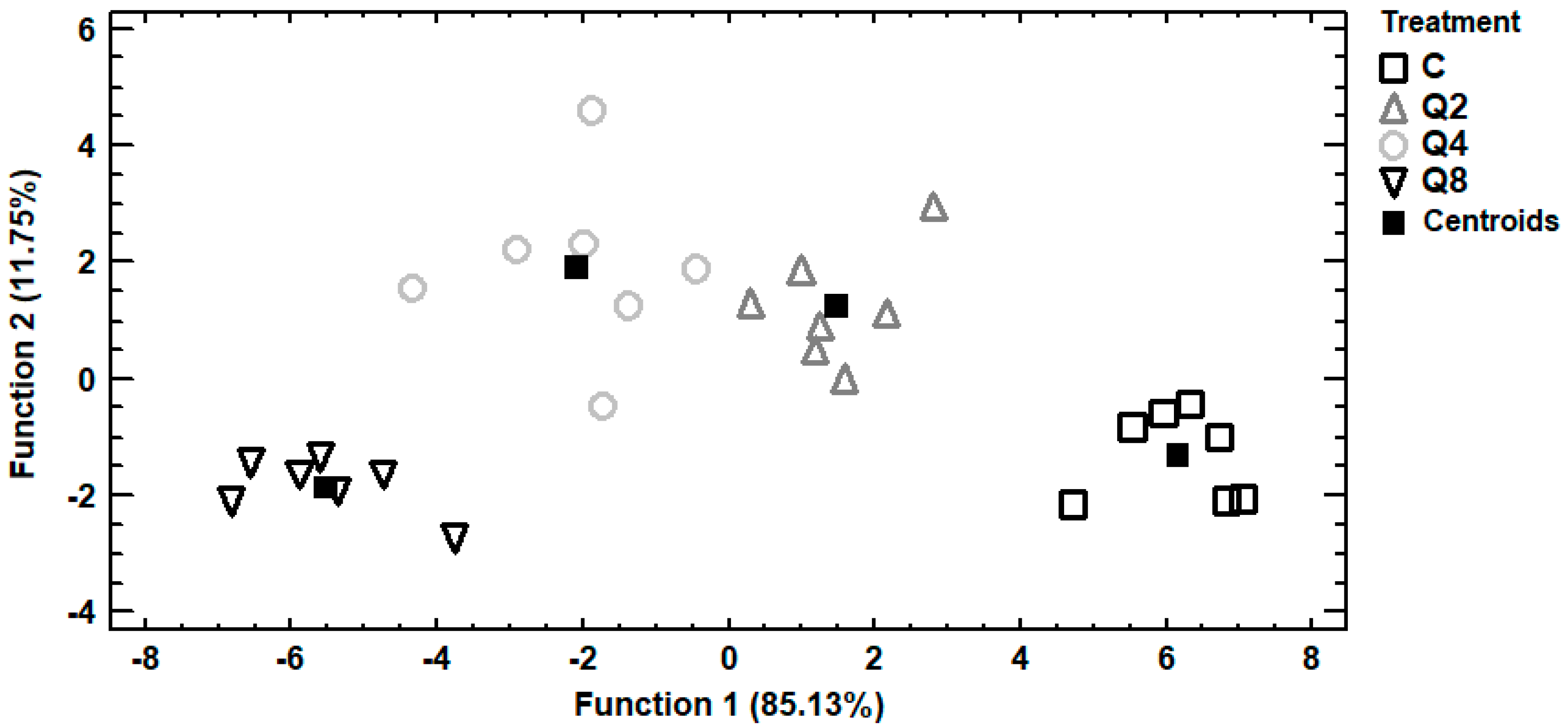
3.2. Principal Components Analysis
3.3. Discriminant Analysis
4. Discussion
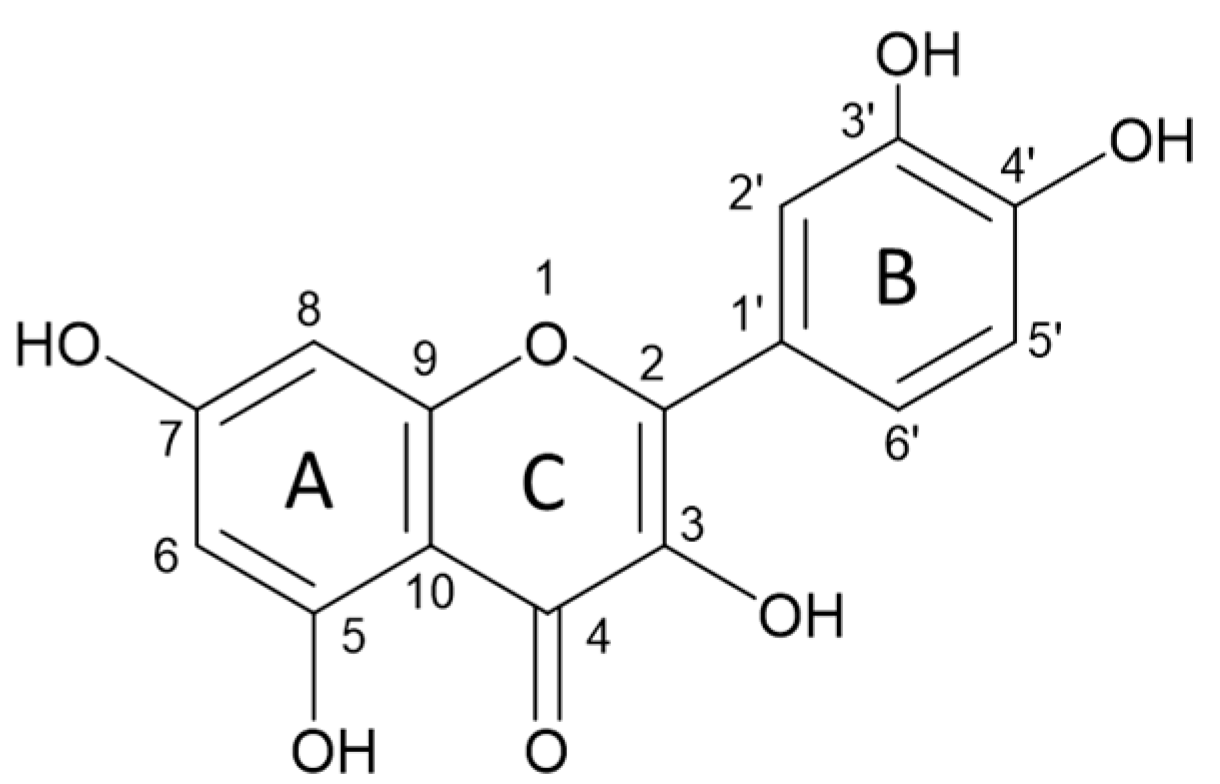
4.1. Flavonoids in Poultry Nutrition
4.2. Modulation of Egg Metallome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement

Conflicts of Interest
References
- Kamboh, A.A.; Leghari, R.A.; Khan, M.A.; Kaka, U.; Naseer, M.; Sazili, A.Q.; Malhi, K.K. Flavonoids supplementation-An ideal approach to improve quality of poultry products. Worlds Poult. Sci. J. 2019, 75, 115–126. [Google Scholar] [CrossRef]
- Batiha, G.-S.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The pharmacological activity, biochemical properties, and pharmacokinetics of the major natural polyphenolic flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, Y.; Liu, H.N.; Suo, Y.L.; Hu, L.L.; Feng, X.A.; Zhang, L.; Jin, F. Effect of quercetin on performance and egg quality during the late laying period of hens, Br. Poul. Sci. 2013, 54, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Naveed, M.; Arain, M.A.; Arif, M. Quercetin: Nutritional and beneficial effects in poultry. Worlds Poult. Sci. J. 2017, 73, 355–364. [Google Scholar] [CrossRef]
- Hager-Theodorides, A.L.; Goliomytis, M.; Delis, S.; Deligeorgis, S. Effects of dietary supplementation with quercetin on broiler immunological characteristics. Anim. Feed Sci. Technol. 2014, 198, 224–230. [Google Scholar] [CrossRef]
- Comalada, M.; Ballester, I.; Bailon, E.; Xaus, J.; Galvez, J.; De Medina, F.S.; Zarzuelo, A. Inhibition of pro-inflammatory markers in primary bone marrow-derived mousemacrophages by naturally occurring flavonoids: Analysis of the structure-activity relationship. Biochem. Pharmacol. 2006, 72, 1010–1021. [Google Scholar] [CrossRef]
- Malešev, D.; Kuntić, V. Investigation of metal-flavonoid chelates and the determination of flavonoids via metal-flavonoid complexing reactions. J. Serb. Chem. Soc. 2007, 72, 921–939. [Google Scholar] [CrossRef]
- Manach, C.; Texier, O.; Morand, C.; Crespy, V.; Régérat, F.; Demigné, C.; Rémésy, C. Comparison of the bioavailability of quercetin and catechin in rats. Free Radic. Biol. Med. 1999, 27, 1259–1266. [Google Scholar] [CrossRef]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2019, 1–14. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, S.J.; Yim, D.G.; Hur, S.J. Changes in the content and bioavailability of onion quercetin and grape resveratrol during in vitro human digestion. Foods 2020, 9, 694. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to quercetin and protection of DNA, proteins and lipids from oxidative damage (ID 1647), “cardiovascular system” (ID 1844), “mental state and performance” (ID 1845), and “liver, kidneys” (ID 1846) pursuant to Article 13 (1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2067. [Google Scholar] [CrossRef]
- Iqbal, M.; Cawthon, D.; Beers, K.; Wideman, R.F., Jr.; Bottje, W.G. Antioxidant enzyme activities and mitochondrial fatty acids in pulmonary hypertension syndrome (PHS) in broilers. Poult. Sci. 2002, 81, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, M.; Yamamoto, H.; Sato, T.; Mizuha, Y.; Kawai, Y.; Taketani, Y.; Kato, S.; Terao, J.; Inakuma, T.; Takeda, E. Dietary quercetin inhibits bone loss without effect on the uterus in ovariectomized mice. J. Bone Miner. Metabol. 2009, 27, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Faris, A.N.; Comstock, A.T.; Wang, Q.; Nanua, S.; Hershenson, M.B.; Sajjan, U.S. Quercetin inhibits rhinovirus replication in vitro and in vivo. Antivir. Res. 2012, 94, 258–271. [Google Scholar] [CrossRef]
- Kuo, S.M.; Leavitt, P.S.; Lin, C.P. Dietary flavonoids interact with trace metals and affect metallothionein level in human intestinal cells. Biol. Trace Elem. Res. 1998, 62, 135–153. [Google Scholar] [CrossRef]
- Xiao, L.; Luo, G.; Tang, Y.; Yao, P. Quercetin and iron metabolism: What we know and what we need to know. Food Chem. Toxicol. 2018, 114, 190–203. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, L.; Wang, J.; Tang, N. Antioxidative and anti-tumour activities of solid quercetin metal (II) complexes. Transit. Metal Chem. 2001, 26, 57–63. [Google Scholar] [CrossRef]
- Berroukeche, F.; Mokhtari-Soulimane, N.; Imessaoudene, A.; Cherrak, A.; Ronot, P.; Boos, A.; Belhandouz, A.; Merzouk, H.; Elhabiri, M. Oral supplementation effect of iron and its complex form with quercetin on oxidant status and on redistribution of essential metals in organs of streptozotocin diabetic rats. Rom. J. Diabetes Nutr. Metab. Dis. 2019, 26, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Simitzis, P.; Spanou, D.; Glastra, N.; Goliomytis, M. Impact of dietary quercetin on laying hen performance, egg quality and yolk oxidative stability. Anim. Feed Sci. Technol. 2018, 239, 27–32. [Google Scholar] [CrossRef]
- Pappas, A.C.; Zoidis, E.; Georgiou, C.A.; Demiris, N.; Surai, P.F.; Fegeros, K. Influence of organic selenium supplementation on the accumulation of toxic and essential trace elements involved in the antioxidant system of chicken. Food Addit. Contam. 2011, 28, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Georgiou, C.A.; Danezis, G.P. Comprehensive Analytical Chemistry; Pico, Y., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; Volume 68, Chapter 3; pp. 131–243. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.M.M.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Yu, Y.L.; Cheng, W.C.; OuYang, C.N.; Fu, E.; Chu, C.L. Immunosuppressive effect of quercetin on dendritic cell activation and function. J. Immunol. 2010, 184, 6815–6821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, Z.; Chadha, K.; Lieberman, A.; Hojnacki, D.; Drake, A.; Zamboni, P.; Rocco, P.; Grazioli, E.; Weinstock-Guttman, B.; Munschauer, F. Quercetin and interferon-beta modulate immune response(s) in peripheral blood mononuclear cells isolated from multiple sclerosis patients. J. Neuroimmunol. 2008, 205, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Schlachterman, A.; Valle, F.; Wall, K.M.; Azios, N.G.; Castillo, L.; Morell, L.; Washington, A.V.; Cubano, L.A.; Dharmawardhane, S.F. Combined resveratrol, quercetin, and catechin treatment reduces breast tumor growth in a nude mouse model. Transl. Oncol. 2008, 1, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupasinghe, H.P.V.; Ronalds, C.M.; Rathgeber, B.; Robinson, R.A. Absorption and tissue distribution of dietary quercetin and quercetin glycosides of apple skin in broiler chickens. J. Sci. Food Agric. 2010, 90, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Iskender, H.; Yenice, G.; Dokumacioglu, E.; Kaynar, O.; Hayirli, A.; Kaya, A. The effects of dietary flavonoid supplementation on the antioxidant status of laying hens. Braz. J. Poult. Sci. 2016, 18, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Kim, I.H. Effect of quercetin (flavonoid) supplementation on growth performance, meat stability, and immunological response in broiler chickens. Livest. Sci. 2020, 104286. [Google Scholar] [CrossRef]
- Liu, H.N.; Liu, Y.; Hu, L.L.; Suo, Y.L.; Zhang, L.; Jin, F.; Feng, X.A.; Teng, N.; Li, Y. Effects of dietary supplementation of quercetin on performance, egg quality, cecal microflora populations, and antioxidant status in laying hens. Poult. Sci. 2014, 93, 347–353. [Google Scholar] [CrossRef]
- Pappas, A.C.; Zoidis, E.; Goliomytis, M.; Simitzis, P.E.; Sotirakoglou, K.; Charismiadou, M.A.; Nikitas, C.; Danezis, G.; Deligeorgis, S.G.; Georgiou, C.A. Elemental Metabolomics: Modulation of egg metallome with flavonoids, an exploratory study. Antioxidants 2019, 8, 361. [Google Scholar] [CrossRef] [Green Version]
- Richards, M.P. Trace mineral metabolism in the avian embryo. Poult. Sci. 1997, 76, 152–164. [Google Scholar] [CrossRef]
- Miles, R.D. Trace minerals and avian embryo development. Ciênc. Anim. Bras. 2000, 2, 1–10. [Google Scholar]
- Siddique, S.; Firdous, S.; Durrani, A.I.; Khan, S.J.; Saeed, A. Hesperidin, a citrus flavonoid, increases the bioavailability of micronutrients of Gallus domesticus (chicken) eggshell: In vitro study. Chem. Spec. Bioavailab. 2016, 28, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, A.; Kambe, T. Mg, Zn and Cu transport proteins: A brief overview from physiological and molecular perspectives. J. Nutr. Sci. Vitaminol. 2015, 61, 116–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaassen, C.D.; Liu, J.; Choudhuri, S. Metallothionein: An intracellular protein to protect against cadmium toxicity. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 267–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganapathy, V.; Ganapathy, M.E.; Leibach, F.H. Intestinal transport of peptides and amino acids. Curr. Top. Membr. 2000, 50, 379–412. [Google Scholar] [CrossRef]
- Ferrali, M.; Signorini, C.; Caciotti, B.; Sugherini, L.; Ciccoli, L.; Giachetti, D.; Comporti, M. Protection against oxidative damage of erythrocyte membrane by the flavonoid quercetin and its relation to iron chelating activity. FEBS Lett. 1997, 416, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Dolatabadi, J.E.N. Molecular aspects on the interaction of quercetin and its metal complexes with DNA. Int. J. Biol. Macromol. 2011, 48, 227–233. [Google Scholar] [CrossRef]
- Jabeen, E.; Janjua, N.K.; Ahmed, S.; Murtaza, I.; Ali, T.; Hameed, S. Radical scavenging propensity of Cu2+, Fe3+ complexes of flavonoids and in-vivo radical scavenging by Fe3+-primuletin. Spectrochim. Acta Part A 2017, 171, 432–438. [Google Scholar] [CrossRef]
- Dehghan, G.; Dolatabadi, J.E.N.; Jouyban, A.; Zeynali, K.A.; Ahmadi, S.M.; Kashanian, S. Spectroscopic studies on the interaction of quercetin–terbium(iii) complex with calf thymus DNA. DNA Cell. Biol. 2011, 30, 195–201. [Google Scholar] [CrossRef]
- Jaccob, A.A.; Hussain, S.A.; Hussain, S.A. Effects of long-term use of flavonoids on the absorption and tissue distribution of orally administered doses of trace elements in rats. Pharmacol. Pharm. 2012, 3, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.R.; McCabe, G.P.; McCabe, L.; Jackson, G.S.; Horcajada, M.N.; Offord-Cavin, E.; Peacock, M.; Weaver, C.M. Effect of hesperidin with and without a calcium (Calcilock) supplement on bone health in postmenopausal women. J. Clin. Endocrinol. Metab. 2016, 101, 923–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saponara, S.; Testai, L.; Iozzi, D.; Martinotti, E.; Martelli, A.; Chericoni, S.; Sgaragli, G.; Fusi, F.; Calderone, V. (+/−)-Naringenin as large conductance Ca2+-activated K+ (BKCa) channel opener in vascular smooth muscle cells. Br. J. Pharmacol. 2006, 149, 1013–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Fang, X.; Wu, J. Flavonoids function as antioxidants: By scavenging reactive oxygen species or by chelating iron? Radiat. Phys. Chem. 1997, 50, 271–276. [Google Scholar] [CrossRef]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Helena Florêncio, M.H.; Jennings, K.R. Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radical Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef]
- Ghosh, N.; Chakraborty, T.; Mallick, S.; Mana, S.; Singha, D.; Ghosh, B.; Roy, S. Synthesis, characterization and study of antioxidant activity of quercetin–magnesium complex. Spectrochim. Acta Part A 2015, 151, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, H.; Liu, J.; Hong, H.; Yan, C. The influence of flavonoid amendment on the absorption of cadmium in Avicennia marina roots. Ecotoxicol. Environ. Saf. 2015, 120, 1–6. [Google Scholar] [CrossRef]
- Kim, E.Y.; Pai, T.K.; Han, O. Effect of bioactive dietary polyphenols on zinc transport across the intestinal Caco-2cell monolayers. J. Agric. Food Chem. 2011, 59, 3606–3612. [Google Scholar] [CrossRef] [Green Version]
- Martel, F.; Monteiro, R.; Calhau, C. Effect of polyphenols on the intestinal and placental transport of some bioactive compounds. Nutr. Res. Rev. 2010, 23, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Georgiou, C.A.; Brusic, V. Elemental metabolomics. Brief. Bioinform. 2017, 19, 524–536. [Google Scholar] [CrossRef]
- Aspuru, K.; Villa, C.; Bermejo, F.; Herrero, P.; López, S.G. Optimal management of iron deficiency anemia due to poor dietary intake. Int. J. Gen. Med. 2011, 4, 741–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | ||||||
|---|---|---|---|---|---|---|
| Elements (ppb) | C | Q2 | Q4 | Q8 | SEM | p-Value |
| As | 4.27 a | 4.40 a | 5.06 b | 5.31 b | 0.113 | <0.001 |
| Ca | 7.15 × 104 | 6.78 × 104 | 5.59 × 104 | 5.19 × 104 | 7.41 × 103 | 0.214 |
| Cd | 2.77 a | 3.73 b | 3.14 ab | 3.67 b | 0.233 | 0.022 |
| Co | 1.31 | 1.60 | 1.33 | 1.33 | 0.099 | 0.147 |
| Cr | 89.5 c | 87.2 c | 81.2 b | 76.3 a | 1.386 | <0.001 |
| Cu | 276.7 | 244.3 | 246.7 | 248.7 | 15.81 | 0.446 |
| Fe | 5.21 × 103 | 7.31 × 103 | 6.65 × 103 | 5.50 × 103 | 806.6 | 0.246 |
| Mg | 1.06 × 105 | 0.908 × 105 | 1.02 × 105 | 0.959 × 105 | 4.74 × 103 | 0.154 |
| Mn | 58.5 | 65.4 | 61.7 | 62.7 | 9.014 | 0.959 |
| Ni | 40.7 | 37.0 | 37.9 | 40.8 | 2.207 | 0.519 |
| Sb | 0.019 a | 0.014 a | 0.585 a | 2.69 b | 0.426 | 0.001 |
| Se | 88.6c | 84.2 bc | 67.7 ab | 58.0 a | 6.672 | 0.011 |
| Sr | 150.3 | 152.9 | 132.4 | 146.4 | 9.593 | 0.454 |
| V | 78.8 | 81.6 | 82.9 | 84.3 | 1.631 | 0.129 |
| Zn | 1.68 × 103 | 1.41 × 103 | 1.18 × 103 | 1.49 × 103 | 143.8 | 0.129 |
| Treatment | ||||||
|---|---|---|---|---|---|---|
| Elements (ppb) | C | Q2 | Q4 | Q8 | SEM | p-value |
| As | 3.73 | 3.61 | 3.69 | 3.71 | 0.110 | 0.887 |
| Ca | 1.03 × 106 | 1.11 × 106 | 1.13 × 106 | 1.11 × 106 | 3.73 × 104 | 0.291 |
| Cd | 3.93 | 5.47 | 4.61 | 4.31 | 0.437 | 0.108 |
| Co | 3.30 | 3.44 | 3.73 | 3.41 | 0.134 | 0.164 |
| Cr | 148.1 | 155.7 | 152.6 | 150.7 | 4.072 | 0.614 |
| Cu | 1436 | 1473 | 1578 | 1509 | 43.46 | 0.153 |
| Fe | 6.89 × 104 | 7.20 × 104 | 7.06 × 104 | 6.55 × 104 | 3.86 × 103 | 0.672 |
| Mg | 1.25 × 105 | 1.35 × 105 | 1.29 × 105 | 1.33 × 105 | 3.77 × 103 | 0.307 |
| Mn | 1141 | 1169 | 1221 | 1179 | 79.01 | 0.914 |
| Ni | 66.7 | 70.1 | 67.9 | 68.0 | 2.220 | 0.754 |
| Sb | 0.077 a | 0.408 ab | 0.582 b | 0.613 b | 0.116 | 0.012 |
| Se | 232.9 | 316.4 | 236.8 | 301.5 | 29.76 | 0.125 |
| Sr | 696.9 | 752.9 | 725.0 | 697.4 | 34.28 | 0.618 |
| V | 104.7 a | 114.2 b | 114.4 b | 112.7 b | 2.173 | 0.016 |
| Zn | 4.00 × 104 | 4.15 × 104 | 4.22 × 104 | 4.20 × 104 | 1.21 × 103 | 0.577 |
| Treatment | ||||||
|---|---|---|---|---|---|---|
| Elements (ppb) | C | Q2 | Q4 | Q8 | SEM | p-value |
| As | 16.63 d | 14.21 c | 9.84 b | 7.24 a | 0.857 | <0.001 |
| Ca | 2.27 × 108 a | 2.39 × 108 ab | 2.44 × 108 b | 2.39 × 108 ab | 4.00 × 106 | 0.040 |
| Cd | 70.2 c | 63.8 bc | 54.5 b | 37.7 a | 4.098 | <0.001 |
| Co | 270.6 | 281.7 | 288.3 | 270.6 | 5.699 | 0.097 |
| Cr | 692.9 b | 663.1 ab | 642.9 a | 625.2 a | 14.99 | 0.024 |
| Cu | 1218 b | 996.0 ab | 843.3 a | 905.1 a | 80.16 | 0.016 |
| Fe | 1.79 × 106 a | 1.96 × 106 b | 2.00 × 106 b | 1.93 × 106 b | 4.20 × 104 | 0.033 |
| Mg | 2.87 × 106 a | 2.99 × 106 a | 3.28 × 106 b | 3.06 × 106 a | 8.66 × 104 | 0.020 |
| Mn | 491.1 | 632.1 | 514.6 | 488.6 | 48.70 | 0.148 |
| Ni | 9.49 × 103 a | 1.02 × 104 b | 1.07 × 104 b | 1.06 × 104 b | 192.3 | <0.001 |
| Sb | 33.86 a | 47.70 a | 72.26 b | 94.88 c | 6.907 | <0.001 |
| Se | 273.9 b | 221.9 ab | 156.8 a | 154.2 a | 27.78 | 0.015 |
| Sr | 1.05 × 105 | 1.11 × 105 | 1.10 × 105 | 1.07 × 105 | 3.42 × 103 | 0.506 |
| V | 663.9 c | 654.7 c | 618.4 b | 572.2 a | 9.019 | <0.001 |
| Zn | 6.95 × 103 | 9.02 × 103 | 6.51 × 103 | 6.11 × 103 | 888.4 | 0.124 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoidis, E.; Pappas, A.C.; Goliomytis, M.; Simitzis, P.E.; Sotirakoglou, K.; Tavrizelou, S.; Danezis, G.; Georgiou, C.A. Quercetin and Egg Metallome. Antioxidants 2021, 10, 80. https://doi.org/10.3390/antiox10010080
Zoidis E, Pappas AC, Goliomytis M, Simitzis PE, Sotirakoglou K, Tavrizelou S, Danezis G, Georgiou CA. Quercetin and Egg Metallome. Antioxidants. 2021; 10(1):80. https://doi.org/10.3390/antiox10010080
Chicago/Turabian StyleZoidis, Evangelos, Athanasios C. Pappas, Michael Goliomytis, Panagiotis E. Simitzis, Kyriaki Sotirakoglou, Savvina Tavrizelou, George Danezis, and Constantinos A. Georgiou. 2021. "Quercetin and Egg Metallome" Antioxidants 10, no. 1: 80. https://doi.org/10.3390/antiox10010080
APA StyleZoidis, E., Pappas, A. C., Goliomytis, M., Simitzis, P. E., Sotirakoglou, K., Tavrizelou, S., Danezis, G., & Georgiou, C. A. (2021). Quercetin and Egg Metallome. Antioxidants, 10(1), 80. https://doi.org/10.3390/antiox10010080