
Albumin from Erythrina edulis (Pajuro) as a Promising Source of Multifunctional Peptides


 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Obtention of Seed Flour and Analysis of Its Proximate Composition
2.3. Protein Fractionation of Seed Flour
2.4. Enzymatic Hydrolysis of Pajuro Protein Fractions
2.5. Antioxidant Activity
2.6. Angiotensin-Converting Enzyme (ACE) Inhibitory Activity
2.7. In Vitro Anti-Diabetic Activity
2.8. Pancreatic Lipase Inhibitory Activity
2.9. Protective Effects in SH-SY5Y Cells
2.9.1. Cell Culture
2.9.2. Effects on Cell Viability
2.9.3. Protective Effects against Oxidative Stress Induced by FeSO4
2.9.4. Determination of Thiobarbituric Acid Reaction Substances (TBARS)
2.9.5. Determination of Lactate Dehydrogenase (LDH) Activity
2.10. Statistical Analysis
3. Results and Discussion
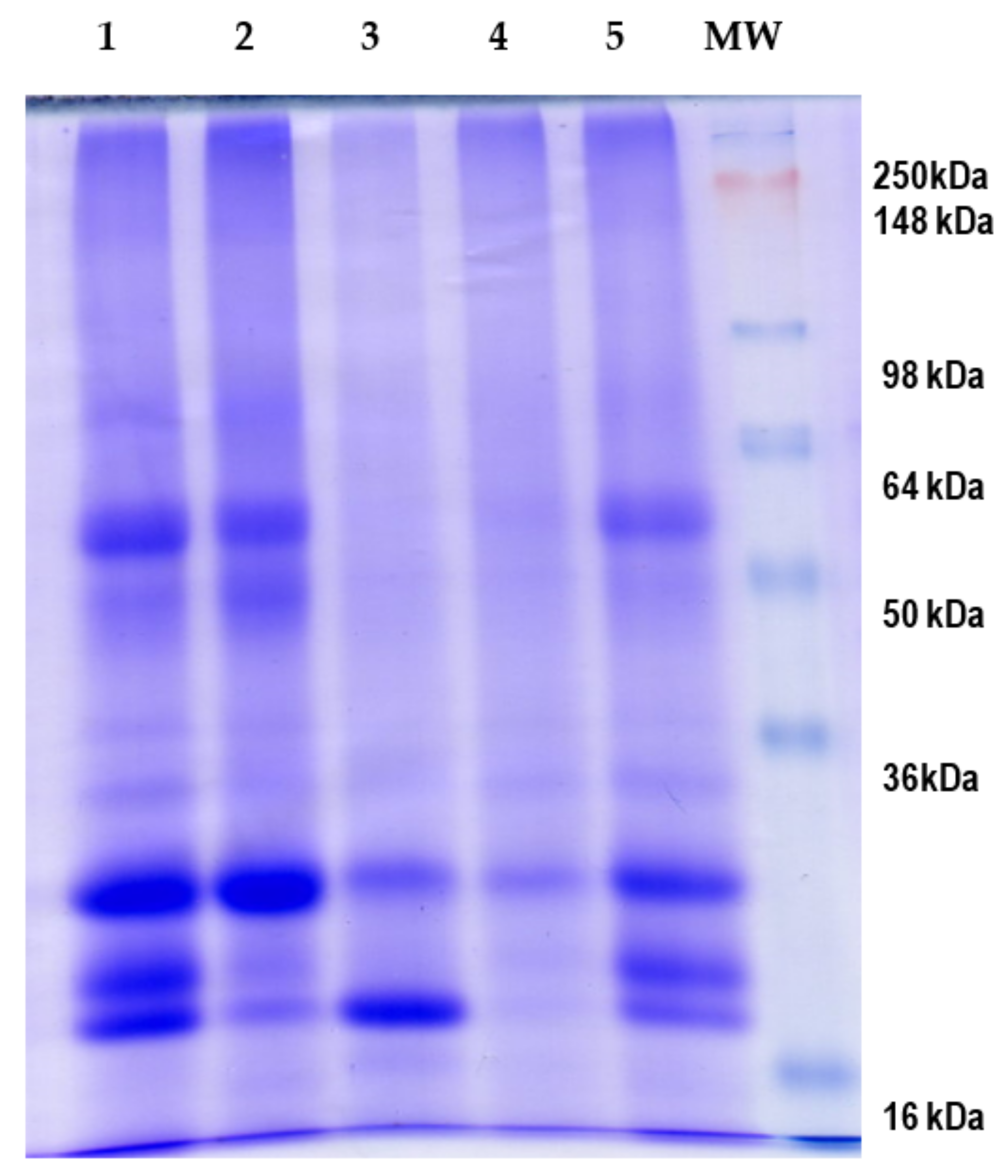
3.1. Obtention and Fractionation of E. edulis Seed Flour
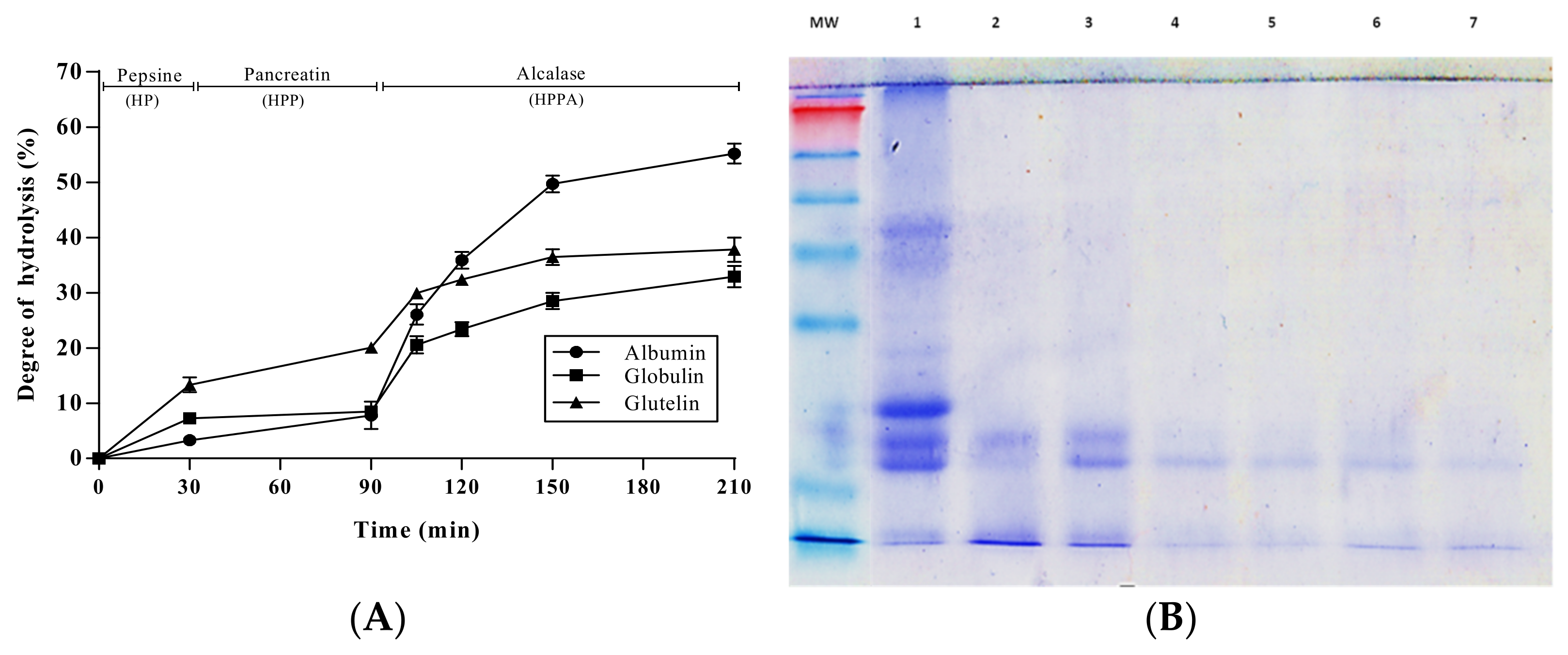
3.2. Enzymatic Hydrolysis of Pajuro Protein Fractions
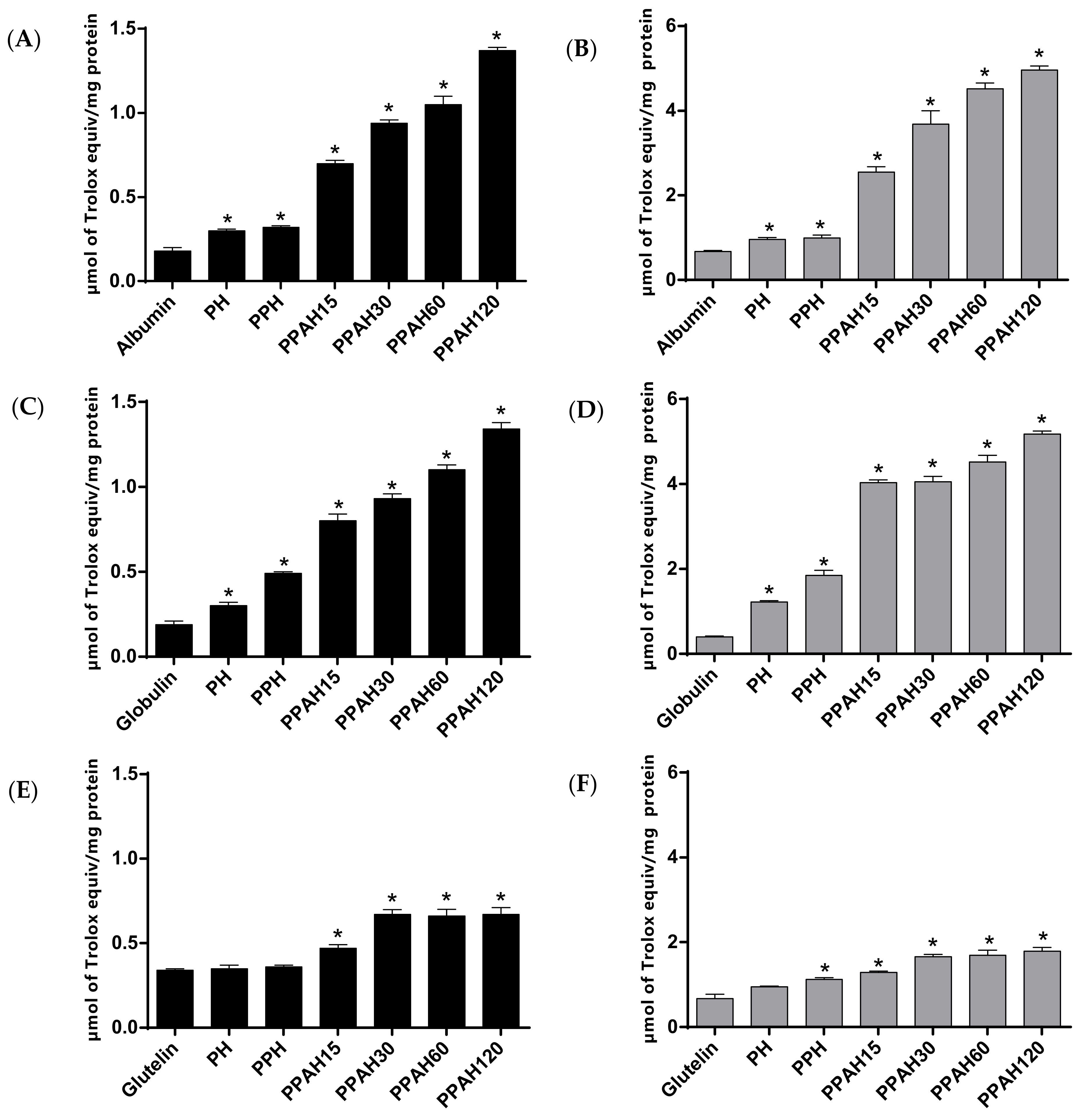
3.3. Antioxidant Activity of Pajuro Protein Fractions
3.4. In Vitro Anti-Hypertensive and Anti-Diabetic Activity of Pajuro Albumin Hydrolyzates
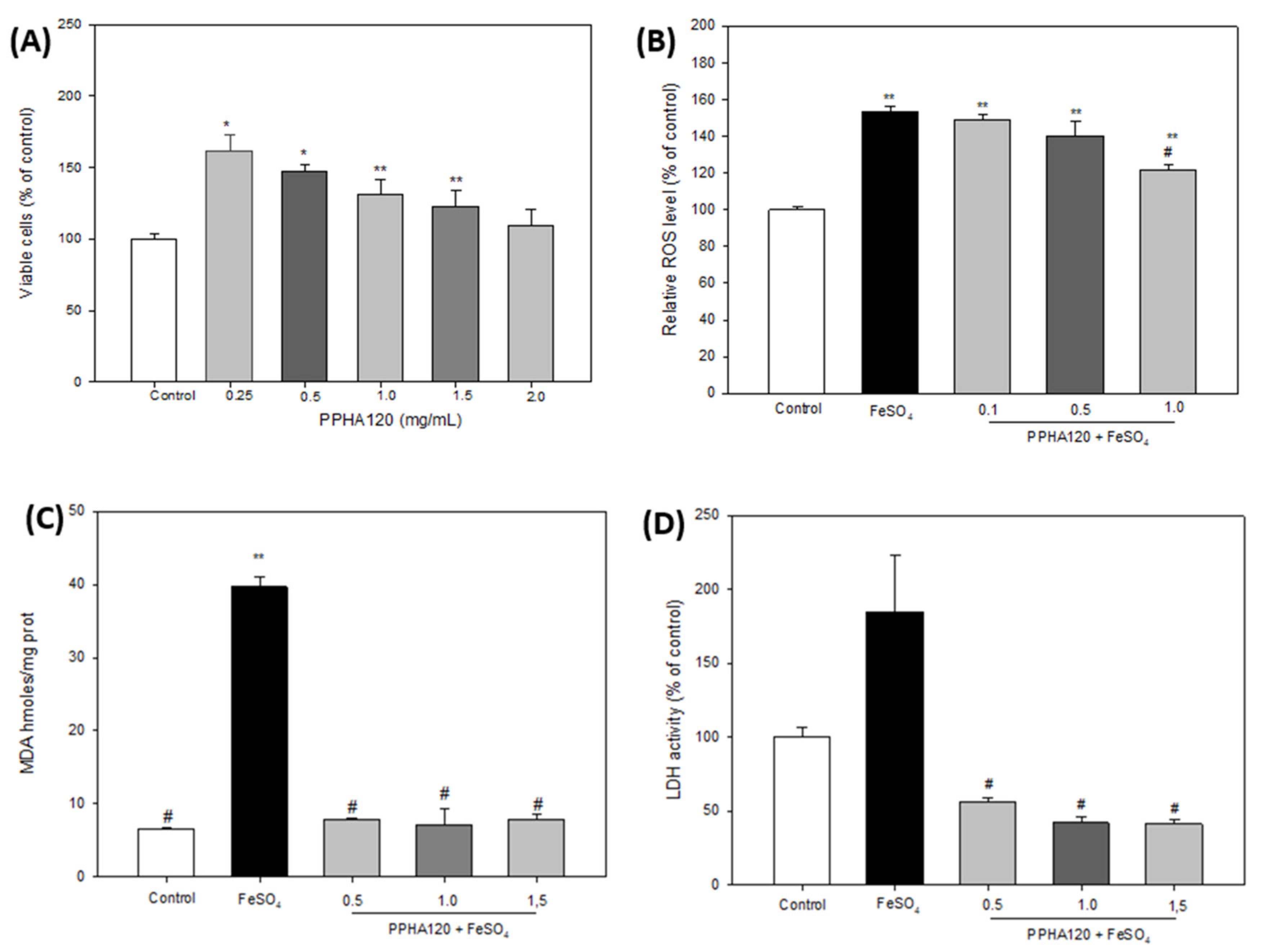
3.5. Neuroprotective Effects of Pajuro Albumin Hydrolyzates
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pihlanto, A.; Mattila, P.; Mäkinena, S.; Pajari, A.-M. Bioactivities of alternative protein sources and their potential health benefits. Food Funct. 2017, 8, 3443–3458. [Google Scholar] [CrossRef] [PubMed]
- Rutherfurd-Markwick, K.J. Food proteins as a source of bioactive peptides with diverse functions. Br. J. Nutr. 2012, 108, S149–S157. [Google Scholar] [CrossRef] [Green Version]
- Chelliah, R.; Wei, S.; Daliri, E.; Elahi, F.; Yeon, S.-J.; Tyagi, A.; Liu, S.; Madar, I.H.; Sultan, G.; Oh, D.-H. The role of bioactive peptides in diabetes and obesity. Foods 2021, 10, 2220. [Google Scholar] [CrossRef] [PubMed]
- Daroit, D.J.; Brandelli, A. In vivo bioactivities of food protein-derived peptides—A current review. Curr. Opin. Food Sci. 2021, 39, 120–129. [Google Scholar] [CrossRef]
- Shobako, N. Hypotensive peptides derived from plant proteins. Peptides 2021, 142, 170573. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Strategies for the discovery and identification of food protein-derived biologically active peptides. Trends Food Sci. Technol. 2017, 69, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, C. The current state of heart disease: Statins, cholesterol, fat and sugar. Int. J. Evid. Based Healthc. 2019, 17, 179–186. [Google Scholar] [CrossRef]
- Ghaedi, E.; Mohammadi, M.; Mohammadi, H.; Ramezani-Jolfaie, N.; Malekzadeh, J.; Hosseinzadeh, M.; Salehi-Abargouei, A. Effects of a paleolithic diet on cardiovascular disease risk factors: A systematic review and meta-analysis of randomized controlled trials. Adv. Nutr. 2019, 10, 634–646. [Google Scholar] [CrossRef]
- Pontes Oliveira de Almeida, A.S.; Silva de Almeida Rezende, M.; Dantas, S.H.; de Lima Silva, S.; Pinheiro Lúcio de Oliveira, J.C.; Araújo de Azevedo, F.L.A.; Ribeiro Alves, R.M.F.; Sales de Menezes, G.M.; Ferreira dos Santos, P.; Felisberto Gonçalves, T.A.; et al. Unveiling the role of inflammation and oxidative stress on age-related cardiovascular diseases. Oxid. Med. Cell. Longev. 2020, 2020, 1954398. [Google Scholar] [CrossRef]
- Daliri, E.B.; Oh, D.H.; Lee, B.H. Bioactive peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Lammi, C.; Aiello, G.; Boschin, G.; Arnoldi, A. Multifunctional peptides for the prevention of cardiovascular disease: A new concept in the area of bioactive food-derived peptides. J. Funct. Foods 2019, 55, 135–145. [Google Scholar] [CrossRef]
- Arango Bedoya, O.; Bolaños Patiño, V.; Ricaurte García, D.; Caicedo, M.; Guerrero, Y. Obtaining a protein extract from chachafruto flour (Erythrina edulis). Rev. Univ. Salud 2012, 14, 161–167. [Google Scholar]
- Intiquilla, I.; Jiménez-Aliaga, K.; Zavaleta, A.I.; Arnao, I.; Peña, C.; Chávez-Hidalgo, E.L.; Hernández-Ledesma, B. Erythrina edulis (pajuro) seed protein: A new source of antioxidant peptides. Nat. Prod. Commun. 2016, 11, 1934578X1601100620. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Chero, M.J.; Sánchez Chero, J.A.; Miranda Zamora, W. Technify and conserve the bioactive components of Pashul (Erythrina edulis) for human consumption. UCV HACER Rev. Inv. Cult. 2019, 8, 11–17. [Google Scholar]
- Intiquilla, A.; Jiménez-Aliaga, K.; Guzmán, F.; Álvarez, C.A.; Zavaleta, A.I.; Izaguirre, V.; Hernández-Ledesma, B. Novel antioxidant peptides obtained by alcalase hydrolysis of Erythrina edulis (pajuro) protein. J. Sci. Food Agric. 2019, 99, 2420–2427. [Google Scholar] [CrossRef]
- Chavan, U.D.; McKenzie, D.B.; Shahidi, F. Functional properties of protein isolates from beach pea (Lathyrus maritimus L.). Food Chem. 2001, 74, 177–187. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Re, R.; Pellergrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Dávalos, A.; Bartolomé, B.; Amigo, L. Preparation of antioxidant enzymatic hydrolysates from α-lactalbumin and β-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J. Agric. Food Chem. 2005, 53, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Hayakari, M.; Kondo, Y.; Izumi, H. A rapid and simple spectrophotometric assay of angiotensin-converting enzyme. Anal. Biochem. 1978, 84, 361–369. [Google Scholar] [CrossRef]
- Subramanian, R.; Asmawi, M.Z.; Sadikun, A. In vitro α-glucosidase and α-amylase enzyme inhibitory effects of Andrographis paniculata extract and andrographolide. Acta Biochim. Pol. 2008, 55, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Kumar Singla, R.; Singh, R.; Kumar Dubey, A. Important aspects of post-prandial antidiabetic drug, acarbose. Curr. Top. Med. Chem. 2016, 16, 2625–2633. [Google Scholar] [CrossRef]
- Johnson, M.; Lucius, A.; Meyer, T.; De Mejia, E. Cultivar evaluation and effect of fermentation on antioxidant capacity and in vitro inhibition of alpha-amylase and alpha-glucosidase by highbush blueberry (Vaccinium corombosum). J. Agric. Food Chem. 2011, 59, 8923–8930. [Google Scholar] [CrossRef]
- Liu, S.; Yu, Z.; Zhu, H.; Zhang, W.; Chen, Y. In vitro alpha-glucosidase inhibitory activity of isolated fractions from water extract of Qingzhuan dark tea. Bmc Complement. Altern. Med. 2016, 16, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Rathi, P.; Gupta, R. Simplified para-nitrophenyl palmitate assay for lipases and esterases. Anal. Biochem. 2002, 311, 98–99. [Google Scholar] [CrossRef]
- Dolenc, A.; Govedarica, B.; Dreu, R.; Kocbek, P.; Srčič, S.; Kristl, J. Nanosized particles of orlistat with enhanced in vitro dissolution rate and lipase inhibition. Int. J. Pharm. 2010, 396, 149–155. [Google Scholar] [CrossRef] [PubMed]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2′,7′-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, L.A.; Pérez, A.; Ruiz, R.; Guzmán, M.A.; Aranda-Olmedo, I.; Clemente, A. Characterization of pea (Pisum sativum) seed protein fractions. J. Sci. Food Agric. 2014, 94, 280–287. [Google Scholar] [CrossRef]
- Moscoso-Mujica, G.; Zavaleta, A.I.; Mujica, A.; Arnao, I.; Moscoso-Neira, C.; Santos, M.; Sánchez, J. Antimicrobial peptides purified from hydrolysates of kanihua (Chenopodium pallidicaule Aellen) seed protein fractions. Food Chem. 2021, 360, 129951. [Google Scholar] [CrossRef]
- Quist, E.E.; Phillips, R.D.; Saalia, F.K. The effect of enzyme systems and processing on the hydrolysis of peanut (Arachis hypogaea L.) protein. LWT Food Sci. Technol. 2009, 42, 1717–1721. [Google Scholar] [CrossRef]
- Castañeda-Pérez, E.; Jiménez-Morales, K.; Castellanos-Ruelas, A.; Chel-Guerrero, L.; Betancur-Ancona, D. Antidiabetic potential of protein hydrolysates and peptide fractions from lima bean (Phaseolus lunatus L): An in vitro study. Int. J. Pept. Res. Ther. 2021, 27, 1979–1988. [Google Scholar] [CrossRef]
- Zhan, Q.; Wang, Q.; Liu, Q.; Guo, Y.; Gong, F.; Hao, L.; Wu, H.; Dong, Z. The antioxidant activity of protein fractions from Sacha inchi seeds after a simulated gastrointestinal digestion. LWT Food Sci. Technol. 2021, 145, 111356. [Google Scholar] [CrossRef]
- Doucet, D.; Otter, D.E.; Gauthier, S.F.; Foegeding, E.A. Enzyme-induced gelation of extensively hydrolyzed whey proteins by Alcalase: Peptide identification and determination of enzyme specificity. J. Agric. Food Chem. 2003, 51, 6300–6308. [Google Scholar] [CrossRef] [PubMed]
- Balderas-León, I.; Baigts-Allende, D.; Cardador-Martínez, A. Antioxidant, angiotensin-converting enzyme, and α-amylase inhibitory activities of protein hydrolysates of Leucaena leucocephala seeds. CyTA J. Food 2021, 19, 349–359. [Google Scholar] [CrossRef]
- Segura Campos, M.R.; Chel Guerrero, L.A.; Betancur Ancona, D.A. Angiotensin-I converting enzyme inhibitory and antioxidant activities of peptide fractions extracted by ultrafiltration of cowpea Vigna unguiculata hydrolysates. J. Sci. Food Agric. 2010, 90, 2512–2518. [Google Scholar] [CrossRef]
- Quintero-Soto, M.F.; Chávez-Ontiveros, J.; Garzón-Tiznado, J.A.; Salazar-Salas, N.Y.; Pineda-Hidalgo, K.V.; Delgado-Vargas, F.; López-Valenzuela, J.A. Characterization of peptides with antioxidant activity and antidiabetic potential obtained from chickpea (Cicer arietinum L.) protein hydrolyzates. J. Food Sci. 2021, 86, 2962–2977. [Google Scholar] [CrossRef]
- Chirinos, R.; Cerna, E.; Pedreschi, R.; Calsin, M.; Aguilar-Galvez, A.; Campos, D. Multifunctional in vitro bioactive properties: Antioxidant, antidiabetic, and antihypertensive of protein hydrolyzates from tarwi (Lupinus mutabilis Sweet) obtained by enzymatic biotransformation. Cereal Chem. 2021, 98, 423–433. [Google Scholar] [CrossRef]
- Chen, C.; Sun-Waterhouse, D.; Zhang, Y.; Zhao, M.; Sun, W. The chemistry behind the antioxidant actions of soy protein isolate hydrolysates in a liposomal system: Their performance in aqueous solutions and liposomes. Food Chem. 2020, 323, 126789. [Google Scholar] [CrossRef]
- Chirinos, R.; Ochoa, K.; Aguilar-Galvez, A.; Carpentier, S.; Pedreschi, R.; Campos, D. Obtaining of peptides with in vitro antioxidant and angiotensin I converting enzyme inhibitory activities from cañihua protein (Chenopodium pallidicaule Aellen). J. Cereal Sci. 2018, 83, 139–146. [Google Scholar] [CrossRef]
- Chirinos, R.; Pedreschi, R.; Velásquez-Sánchez, M.; Aguilar-Galvez, A.; Campos, D. In vitro antioxidant and angiotensin-I converting enzyme inhibitory properties of enzymatically hydrolysed quinoa (Chenopodium quinoa) and kiwicha (Amaranthus caudatus) proteins. Cereal Chem. 2020, 97, 949–957, In press. [Google Scholar] [CrossRef]
- Jingbo, L.; Chen, Z.; He, J.; Zhang, Y.; Zhang, T.; Jiang, Y. Anti-oxidative and anti-apoptosis effects of egg white peptide, Trp-Asn-Trp-Ala-Asp, against H2O2-induced oxidative stress in human embryonic kidney 293 cells. Food Funct. 2014, 5, 3179–3188. [Google Scholar] [CrossRef]
- Wong, F.C.; Xiao, J.; Wang, S.; Ee, K.Y.; Chai, T.T. Advances on the antioxidant peptides from edible plant sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Kusumah, J.; Real Hernandez, L.M.; Gonzalez de Mejia, E. Antioxidant potential of mung bean (Vigna radiata) albumin peptides produced by enzymatic hydrolysis analyzed by biochemical and in silico methods. Foods 2020, 9, 1241. [Google Scholar] [CrossRef] [PubMed]
- Tom, B.; Dendorfer, A.; Jan Danser, A.H. Bradykinin, angiotensin-(1-7), and ACE inhibitors: How do they interact? Int. J. Biochem. Cell Biol. 2003, 35, 792–801. [Google Scholar] [CrossRef]
- Chin, Y.Y.; Chew, L.Y.; Toh, G.T.; Salampessy, J.; Azlan, A.; Ismail, A. Nutritional composition and angiotensin converting enzyme inhibitory activity of blue lupin (Lupinus angustifolius). Food Biosci. 2019, 31, 100401. [Google Scholar] [CrossRef]
- Esfandi, R.; Seidu, I.; Willmore, W.; Tsopmo, A. Antioxidant, pancreatic lipase, and α-amylase inhibitory properties of oat bran hydrolyzed proteins and peptides. J. Food Biochem. 2021, 00, e13762. [Google Scholar] [CrossRef]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in vitro activities of a-amylase, a-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Mojica, L.; de Mejia, E.G. Optimization of enzymatic production of antidiabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Liang, K.; Jin, Y.; Zhang, M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel a-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, S.O.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Effect of protease type and peptide size on the in vitro antioxidant, antihypertensive and anti-diabetic activities of eggplant leaf protein hydrolysates. Foods 2021, 10, 1112. [Google Scholar] [CrossRef]
- Holst, J.J.; Deacon, C.F. Glucagon-like peptide 1 and inhibitors of dipeptidyl peptidase IV in the treatment of type 2 diabetes mellitus. Curr. Opin. Pharmacol. 2004, 4, 589–596. [Google Scholar] [CrossRef]
- Mentlein, R. Therapeutic assessment of glucagon-like peptide-1 agonists compared with dipeptidyl peptidase IV inhibitors as potential antidiabetic drugs. Expert Opin. Investig. Drugs 2005, 14, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Mitani, H.; Takimoto, M.; Hughes, T.E.; Kimura, M. Dipeptidyl peptidase IV inhibition improves impaired glucose tolerance in high-fat diet-fed rats: Study using a Fischer 344 rat substrain deficient in its enzyme activity. Jpn. J. Pharmacol. 2002, 88, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Zhang, L.; Song, W.; Zhang, C.; Hua, Y.; Chen, Y.; Li, X. Separation, identification and molecular binding mechanism of dipeptidyl peptidase IV inhibitory peptides derived from walnut (Juglans regia L.) protein. Food Chem. 2021, 347, 129062. [Google Scholar] [CrossRef]
- Kamal, H.; Mudgil, P.; Bhaskar, B.; Fisayo, A.F.; Gan, C.-Y.; Maqsood, S. Amaranth proteins as potential source of bioactive peptides with enhanced inhibition of enzymatic markers linked with hypertension and diabetes. J. Cereal Sci. 2021, 101, 103308. [Google Scholar] [CrossRef]
- Velarde-Salcedo, A.J.; Barrera-Pacheco, A.; Lara-González, S.; Montero-Morán, G.M.; Díaz-Gois, A.; González de Mejia, E.; Barba de la Rosa, A. In vitro inhibition of dipeptidyl peptidase IV by peptides derived from the hydrolysis of amaranth (Amaranthus hypochondriacus L.) proteins. Food Chem. 2013, 136, 758–764. [Google Scholar] [CrossRef]
- Wang, F.; Yu, G.; Zhang, Y.; Zhang, B.; Fan, J. Dipeptidyl peptidase IV inhibitory peptides derived from oat (Avena sativa L.), buckwheat (Fagopyrum esculentum), and highland barley (Hordeum vulgare trifurcatum (L.) Trofim) proteins. J. Agric. Food Chem. 2015, 63, 9543–9549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, R.; Chen, X.; Zeng, Z.; Ma, H.; Chen, S. Dipeptidyl peptidase IV-inhibitory peptides derived from silver carp (Hy-pophthalmichthys molitrix Val.) proteins. J. Agric. Food Chem. 2016, 64, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Lunder, M.; Bratkovic, T.; Kreft, S.; Strukelj, B. Peptide inhibitor of pancreatic lipase selected by phage display using different elution strategies. J. Lipid Res. 2005, 46, 1512–1516. [Google Scholar] [CrossRef] [Green Version]
- Lunagariya, N.A.B.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897–921. [Google Scholar] [PubMed]
- Moreno Valdespino, C.; Gonzalez de Mejia, E.; Mojica, L.; Luna-Vital, D.; Camacho, R. Bioactive peptides from black bean proteins play a potential role in the prevention of adipogenesis. Curr. Dev. Nutr. 2019, 3 (Suppl. 1), P06-119-19. [Google Scholar] [CrossRef] [Green Version]
- Carrera, I.; Cacabelos, R. Current drugs and potential future neuroprotective compounds for Parkinson’s disease. Curr. Neuropharmacol. 2019, 17, 295–306. [Google Scholar] [CrossRef]
- Lee, E.J.; Lee, J.Y.; Seo, S.R.; Chung, K.C. Overexpression of DSCR1 blocks zincinduced neuronal cell death through the formation of nuclear aggregates. Mol. Cell. Neurosci. 2007, 35, 585–595. [Google Scholar] [CrossRef]
- Chan, S.; Kantham, S.; Rao, V.M.; Palanivelu, M.K.; Pham, H.L.; Shaw, P.N.; McGeary, R.P.; Ross, B.P. Metal chelation, radical scavenging and inhibition of Aβ42 fibrillation by food constituents in relation to Alzheimer’s disease. Food Chem. 2016, 199, 185–194. [Google Scholar] [CrossRef] [Green Version]
- González-Sarrías, A.; Núñez-Sánchez, M.Á.; Tomás-Barberán, F.A.; Espín, J.C. Neuroprotective effects of bioavailable polyphenol-derived metabolites against oxidative stress-induced cytotoxicity in human neuroblastoma SH-SY5Y cells. J. Agric. Food Chem. 2017, 65, 752–758. [Google Scholar] [CrossRef] [Green Version]
- Sheng, J.; Yang, X.; Chen, J.; Peng, T.; Yin, X.; Liu, W.; Liang, M.; Wan, J.; Yang, X. Antioxidative effects and mechanism study of bioactive peptides from defatted walnut (Juglans regia L.) meal hydrolysate. J. Agric. Food Chem. 2019, 67, 3305–3312. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Peng, F.; Wang, X.; Li, M.; Lei, H.; Xu, H. Identification and characterization of antioxidative peptides derived from simulated in vitro gastrointestinal digestion of walnut meal proteins. Food Res. Int. 2019, 116, 518–526. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hur, S.J. Neuroprotective effects of different molecular weight peptide fractions obtained from beef by hydrolysis with commercial enzymes in SH-SY5Y cells. Food Res. Int. 2019, 121, 176–184. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hur, S.J. Protective effect of a 3 kDa peptide obtained from beef myofibrillar protein using alkaline-AK on neuronal cells. Neurochem. Int. 2019, 129, 104459. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Shao, B.; Yu, H.; Xu, F.; Wang, P.; Yu, K.; Han, Y.; Song, M.; Li, Y.; Cao, Z. Neuroprotective role of hyperforin on aluminum maltolate-induced oxidative damage and apoptosis in PC12 cells and SH-SY5Y cells. Chem. Biol. Interact. 2019, 299, 15–26. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Content (g/100 g Protein) | |||||
|---|---|---|---|---|---|---|
| Seed Flour | Albumin | Globulin | Glutelin | Prolamin | FAO | |
| Essential | ||||||
| Lys | 2.06 | 4.39 | 4.48 | 3.20 | 1.88 | 5.2 |
| Trp | n.d. | n.d. | n.d. | n.d. | n.d. | 0.7 |
| Phe | 1.98 | 4.25 | 4.01 | 3.12 | 1.57 | 4.6 a |
| Tyr | 1.23 | 3.82 | 3.92 | 2.63 | 0.74 | |
| Met | 0.38 | 1.18 | 1.22 | 0.93 | 0.39 | 2.6 b |
| Cys | 0.89 | 1.02 | 1.03 | 0.49 | 0.45 | |
| Thr | 1.19 | 2.26 | 2.41 | 1.82 | 0.89 | 2.7 |
| Leu | 2.68 | 6.11 | 5.98 | 4.47 | 2.43 | 6.3 |
| Ile | 1.16 | 2.56 | 2.50 | 1.77 | 1.08 | 3.1 |
| Val | 2.11 | 3.62 | 3.38 | 2.36 | 1.68 | 4.2 |
| Non essential | ||||||
| Asx c | 4.78 | 9.57 | 8.99 | 6.50 | 3.44 | |
| Glx d | 5.50 | 11.98 | 11.65 | 8.05 | 4.15 | |
| Ser | 2.24 | 5.11 | 5.16 | 3.57 | 1.66 | |
| His | 1.50 | 1.90 | 1.88 | 1.59 | 0.68 | |
| Arg | 1.05 | 3.58 | 3.63 | 2.45 | 0.86 | |
| Ala | 1.68 | 3.30 | 2.99 | 2.62 | 1.59 | |
| Pro | 2.19 | 4.17 | 3.98 | 2.74 | 1.58 | |
| Gly | 1.69 | 3.83 | 3.45 | 2.55 | 1.43 | |
| TAA | 34.31 | 72.65 | 70.66 | 50.86 | 26.50 | |
| HAA | 14.30 | 30.03 | 29.01 | 21.13 | 11.51 | |
| AAA | 3.21 | 8.07 | 7.93 | 5.75 | 2.31 | |
| Sample | ACE Inhibition (%) a | α-Amylase Inhibition (%) b | α-Glucosidase Inhibition (%) b | DPP-IV Inhibition (%) c | Pancreatic Lipase Inhibition (%) a |
|---|---|---|---|---|---|
| Albumin | 0.10 ± 0.04 | 22.80 ± 6.85 | 1.61 ± 0.18 | 2.31 ± 1.48 | n.d. |
| Albumin PH | 13.45 ± 1.88 * | n.d. | n.d. | 16.16 ± 1.48 * | n.d. |
| Albumin PPH | 20.85 ± 1.06 * | n.d. | n.d. | 35.73 ± 1.35 * | n.d. |
| Albumin PPHA15 | 76.75 ± 4.48 ** | n.d. | n.d. | 60.69 ± 6.70 ** | n.d. |
| Albumin PPHA30 | 64.20 ± 6.15 ** | n.d. | n.d. | 58.19 ± 3.13 ** | n.d. |
| Albumin PPHA60 | 82.22 ± 5.51 ** | n.d. | n.d. | 58.61 ± 9.51 ** | n.d. |
| Albumin PPHA120 | 83.60 ± 0.09 ** | 14.60 ± 1.32 | 6.06 ± 1.61 | 64.81 ± 5.31 ** | n.d. |
| Positive control | 6.81 ± 0.04 | 87.44 ± 1.61 | 70.21 ± 0.47 | 0.86 ± 0.01 | 65.92 ± 2.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palma-Albino, C.; Intiquilla, A.; Jiménez-Aliaga, K.; Rodríguez-Arana, N.; Solano, E.; Flores, E.; Zavaleta, A.I.; Izaguirre, V.; Hernández-Ledesma, B. Albumin from Erythrina edulis (Pajuro) as a Promising Source of Multifunctional Peptides. Antioxidants 2021, 10, 1722. https://doi.org/10.3390/antiox10111722
Palma-Albino C, Intiquilla A, Jiménez-Aliaga K, Rodríguez-Arana N, Solano E, Flores E, Zavaleta AI, Izaguirre V, Hernández-Ledesma B. Albumin from Erythrina edulis (Pajuro) as a Promising Source of Multifunctional Peptides. Antioxidants. 2021; 10(11):1722. https://doi.org/10.3390/antiox10111722
Chicago/Turabian StylePalma-Albino, Cleni, Arturo Intiquilla, Karim Jiménez-Aliaga, Nathaly Rodríguez-Arana, Estela Solano, Eduardo Flores, Amparo Iris Zavaleta, Víctor Izaguirre, and Blanca Hernández-Ledesma. 2021. "Albumin from Erythrina edulis (Pajuro) as a Promising Source of Multifunctional Peptides" Antioxidants 10, no. 11: 1722. https://doi.org/10.3390/antiox10111722
APA StylePalma-Albino, C., Intiquilla, A., Jiménez-Aliaga, K., Rodríguez-Arana, N., Solano, E., Flores, E., Zavaleta, A. I., Izaguirre, V., & Hernández-Ledesma, B. (2021). Albumin from Erythrina edulis (Pajuro) as a Promising Source of Multifunctional Peptides. Antioxidants, 10(11), 1722. https://doi.org/10.3390/antiox10111722