
Antioxidant Properties and Cytoprotective Effect of Pistacia lentiscus L. Seed Oil against 7β-Hydroxycholesterol-Induced Toxicity in C2C12 Myoblasts: Reduction in Oxidative Stress, Mitochondrial and Peroxisomal Dysfunctions and Attenuation of Cell Death

 ,
,  , , ,
, , ,  , , ,
, , ,  ,
, 
 ,
, 
Abstract
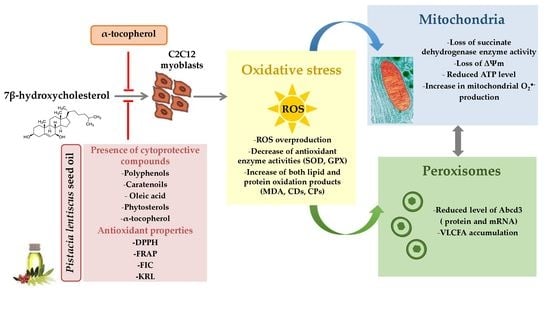
:
1. Introduction
2. Material and Methods
2.1. Chemical Profile of Pistacia lentiscus L. Seed Oil
2.1.1. Seed Material and Oil Extraction
2.1.2. Colorimetric Determination of Total Phenolics, Flavonoids, and Carotenoids Contents of Pistacia lentiscus L. Seed Oil
2.1.3. Polyphenols Analysis
2.1.4. Fatty Acids Analysis
2.1.5. Phytosterols Analysis
2.1.6. α-Tocopherol Analysis
2.2. Antioxidant Activity of Pistacia lentiscus L. Seed Oil
2.2.1. Free Radical Scavenging Activity with DPPH Assay
2.2.2. Ferric Reducing Antioxidant Power (FRAP) Assay
2.2.3. Ferrous-Ion Chelating (FIC) Assay
2.2.4. KRL Test
2.3. In Vitro Study
2.3.1. Cell Culture and Treatments
2.3.2. Evaluation of Cell Morphology by Phase-Contrast Microscopy
2.3.3. Evaluation of Cell Viability with the MTT Assay
2.3.4. Measurement of Cell Viability with the Fluorescein Diacetate Assay
2.3.5. Measurement of Plasma Membrane Permeability with Propidium Iodide
2.3.6. Measurement of Oxidative Stress
Evaluation of Reactive Oxygen Species Production with Dihydroethidium
Quantification of Antioxidant Enzymes Activities: Glutathione Peroxidase (GPx) and Superoxide Dismutase (SOD)
Measurement of Lipid Peroxidation Products: Malondialdehyde (MDA) and Conjugated Dienes (CDs)
2.3.7. Evaluation of Mitochondrial Function
Measurement of Transmembrane Mitochondrial Potential with DiOC6(3)
Measurement of ATP Levels
Measurement of Mitochondrial Reactive Oxygen Species with MitoSOX-Red
2.3.8. Determination of the Peroxisomal Status
Evaluation of the Level and Topography of Abcd3 Peroxisomal Transporter by Structured Illumination Microscopy (Apotome)
Flow Cytometric Quantification of Abcd3 Peroxisomal Transporter
Gas Chromatography—Mass Spectrometry Analysis of Fatty Acids
Transmission Electron Microscopy Analysis
Analysis of Abcd3 Peroxisomal Transporter mRNA by Real-Time Quantitative Polymerase Chain Reaction
2.4. Gas Chromatography—Mass Spectrometry Analysis of Cholesterol and Oxysterols Oxidized at C7 (7-Ketocholesterol, 7β-Hydroxycholesterol) in the Plasma of Sarcopenic Patients
2.5. Statistical Analysis
3. Results
3.1. Biochemical Composition of Pistacia lentiscus L. Seed Oil
3.2. Evaluation of the Antioxidant Properties of Pistacia lentiscus L. Seed Oil
3.3. Evaluation of 7-Ketocholesterol and 7β-Hydroxycholesterol Plasma Levels in Sarcopenic and Non-Sarcopenic Subjects
3.4. Evaluation of the Effects of Pistacia lentiscus L. Seed Oil on 7β-Hydroxycholesterol-Induced Morphological Changes and Cell Death
3.5. Evaluation of the Effects of Pistacia lentiscus L. Seed Oil on 7β-Hydroxycholesterol-Induced Oxidative Stress
3.6. Evaluation of the Effects of Pistacia lentiscus L. Seed Oil on 7β-Hydroxycholesterol-Induced Mitochondrial Damages
3.7. Evaluation of the Effects of Pistacia lentiscus L. Seed Oil on 7β-Hydroxycholesterol-Induced Peroxisomal Damages
3.8. Evaluation by Transmission Electron Microscopy of the Effects of Pistacia lentiscus L. Seed Oil on 7β-Hydroxycholesterol-Induced Cellular, Mitochondrial, and Peroxisomal Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knapowski, J.; Wieczorowska-Tobis, K.; Witowski, J. Pathophysiology of ageing. J. Physiol. Pharmacol. 2002, 53, 135–146. [Google Scholar] [PubMed]
- Rosenberg, I.H.; Roubenoff, R. Stalking sarcopenia. Ann. Intern. Med. 1995, 123, 727–728. [Google Scholar] [CrossRef] [PubMed]
- Mankhong, S.; Kim, S.; Moon, S.; Kwak, H.B.; Park, D.H.; Kang, J.H. Experimental Models of Sarcopenia: Bridging molecular mechanism and therapeutic strategy. Cells 2020, 9, 1385. [Google Scholar] [CrossRef] [PubMed]
- Troen, B.R. The biology of aging. Mt. Sinai J. Med. 2003, 70, 3–22. [Google Scholar] [PubMed]
- Melton, L.J.; Khosla, S.; Crowson, C.S.; O'Connor, M.K.; O’Fallon, W.M.; Riggs, B.L. Epidemiology of sarcopenia. J. Am. Geriatr. Soc. 2000, 48, 625–630. [Google Scholar] [CrossRef]
- Pereira, A.F.; Silva, A.J.; Matos Costa, A.; Monteiro, A.M.; Bastos, E.M.; Cardoso Marques, M. Muscle tissue changes with aging. Acta Med. Port. 2013, 26, 51–55. [Google Scholar]
- Marquez, D.X.; Hoyem, R.; Fogg, L.; Bustamante, E.E.; Staffileno, B.; Wilbur, J. Physical activity of urban community-dwelling older latino adults. J. Phys. Act. Health 2011, 8, S161–S170. [Google Scholar] [CrossRef]
- Kirchengast, S.; Huber, J. Gender and age differences in lean soft tissue mass and sarcopenia among healthy elderly. Anthropol. Anz. 2009, 67, 139–151. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritchevsky, S.B.; Nevitt, M.; Schwartz, A.V.; Simonsick, E.M.; Tylavsky, F.A.; Visser, M.; Newman, A.B. The loss of skeletal muscle strength, mass, and quality in older adults: The health, aging and body composition study. J. Gerontol. Med. Sci. 2006, 61, 1059–1064. [Google Scholar] [CrossRef]
- Rathnayake, N.; Alwis, G.; Lenora, J.; Lekamwasam, S. Concordance between appendicular skeletal muscle mass measured with DXA and estimated with mathematical models in middle-aged women. J. Physiol. Anthropol. 2018, 37, 19. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, A.S. Proteomics of skeletal muscle: Focus on insulin resistance and exercise biology. Proteomes 2016, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Gransee, H.M.; Mantilla, C.B.; Sieck, G.C. Respiratory muscle plasticity. Compr. Physiol. 2012, 2, 1441–1462. [Google Scholar]
- Periasamy, M.; Herrera, J.L.; Reis, F.C.G. Skeletal muscle thermogenesis and its role in whole body energy metabolism. Diabetes Metab. J. 2017, 41, 327–336. [Google Scholar] [CrossRef]
- Thomson, D.M.; Winder, W.W. AMPK control of fat metabolism in skeletal muscle. Acta Physiol. 2009, 196, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Muscat, G.E.; Wagner, B.L.; Hou, J.; Tangirala, R.K.; Bischoff, E.D.; Rohde, P.; Petrowski, M.; Li, J.; Shao, G.; Macondray, G.; et al. Regulation of cholesterol homeostasis and lipid metabolism in skeletal muscle by liver X receptors. J. Biol. Chem. 2002, 25, 40722–40728. [Google Scholar] [CrossRef] [Green Version]
- Evans, P.L.; McMillin, S.L.; Weyrauch, L.A.; Witczak, C.A. Regulation of skeletal muscle glucose transport and glucose metabolism by exercise training. Nutrients 2019, 11, 2432. [Google Scholar] [CrossRef] [Green Version]
- Kelley, D.E.; Mandarino, L.J. Hyperglycemia normalizes insulin-stimulated skeletal-muscle glucose-oxidation and storage in noninsulin-dependent diabetes-mellitus. J. Clin. Investig. 1990, 86, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and age-related diseases: From mechanisms to therapeutic strategies. Biogerontology 2021, 22, 165–187. [Google Scholar] [CrossRef]
- Crowe, A.V.; McArdle, A.; McArdle, F.; Pattwell, D.M.; Bell, G.M.; Kemp, G.J.; Bone, J.M.; Griffiths, R.D.; Jackson, M.J. Markers of oxidative stress in the skeletal muscle of patients on haemodialysis. Nephrol. Dial. Transplant. 2007, 22, 1177–1183. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, T.; Ulfhake, B. Sarcopenia: What is the origin of this aging-induced disorder? Front. Genet. 2021, 12, 686526. [Google Scholar] [CrossRef]
- Gomes, M.J.; Martinez, P.F.; Pagan, L.U.; Damatto, R.L.; Cezar, M.D.M.; Lima, A.R.R.; Okoshi, K.; Okoshi, M.P. Skeletal muscle aging: Influence of oxidative stress and physical exercise. Oncotarget 2017, 8, 20428–20440. [Google Scholar] [CrossRef] [Green Version]
- Fulle, S.; Protasi, F.; Di Tano, G.; Pietrangelo, T.; Beltramin, A.; Boncompagni, S.; Vecchiet, L.; Fano, G. The contribution of reactive oxygen species to sarcopenia and muscle ageing. Exp. Gerontol. 2004, 39, 17–24. [Google Scholar] [CrossRef]
- Seo, E.; Kang, H.; Choi, H.; Choi, W.; Jun, H.S. Reactive oxygen species-induced changes in glucose and lipid metabolism contribute to the accumulation of cholesterol in the liver during aging. Aging Cell 2019, 18, e12895. [Google Scholar] [CrossRef] [Green Version]
- Simons, K.; Ikonen, E. How cells handle cholesterol. Science 2000, 290, 1721–1726. [Google Scholar] [CrossRef] [Green Version]
- Iuliano, L. Pathways of cholesterol oxidation via non-enzymatic mechanisms. Chem. Phys. Lipids 2011, 164, 457–468. [Google Scholar] [CrossRef]
- Zerbinati, C.; Iuliano, L. Cholesterol and related sterols autoxidation. Free Radic. Biol. Med. 2017, 111, 151–155. [Google Scholar] [CrossRef]
- Anderson, A.; Campo, A.; Fulton, E.; Corwin, A.; Jerome III, W.G.; O’Connor, M.S. 7-Ketocholesterol in disease and aging. Redox Biol. 2020, 29, 101380. [Google Scholar] [CrossRef]
- Nury, T.; Yammine, A.; Ghzaiel, I.; Sassi, K.; Zarrouk, A.; Brahmi, F.; Samadi, M.; Rup-Jacques, S.; Vervandier-Fasseur, D.; de Barros, J.P.; et al. Attenuation of 7-ketocholesterol- and 7β-hydroxycholesterol-induced oxiapoptophagy by nutrients, synthetic molecules and oils: Potential for the prevention of age-related diseases. Ageing Res. Rev. 2021, 68, 101324. [Google Scholar] [CrossRef]
- Mitic, T.; Andrew, R.; Walker, B.R.; Hadoke, P.W.F. 11b-Hydroxysteroid dehydrogenase type 1 contributes to the regulation of 7-oxysterol levels in the arterial wall through the inter-conversion of 7-ketocholesterol and 7b-hydroxycholesterol. Biochimie 2013, 95, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Vejux, A.; Abed-Vieillard, D.; Hajji, K.; Zarrouk, A.; Mackrill, J.J.; Ghosh, S.; Nury, T.; Yammine, A.; Zaibi, M.; Mihoubi, W.; et al. 7-Ketocholesterol and 7β-hydroxycholesterol: In vitro and animal models used to characterize their activities and to identify molecules preventing their toxicity. Biochem. Pharmacol. 2020, 173, 113648. [Google Scholar] [CrossRef] [Green Version]
- Zarrouk, A.; Vejux, A.; Mackrill, J.; O'Callaghan, Y.; Hammami, M.; O'Brien, N.; Lizard, G. Involvement of oxysterols in age-related diseases and ageing processes. Ageing Res. Rev. 2014, 18, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Mutemberezi, V.; Guillemot-Legris, O.; Muccioli, G.G. Oxysterols: From cholesterol metabolites to key mediators. Prog. Lipid Res. 2016, 64, 152–169. [Google Scholar] [CrossRef] [PubMed]
- Samadi, A.; Sabuncuoglu, S.; Samadi, M.; Isikhan, S.Y.; Chirumbolo, S.; Peana, M.; Lay, I.; Yalcinkaya, A.; Bjorklund, G. A Comprehensive Review on Oxysterols and Related Diseases. Curr. Med. Chem. 2021, 28, 110–136. [Google Scholar] [CrossRef] [PubMed]
- Nury, T.; Zarrouk, A.; Vejux, A.; Doria, M.; Riedinger, J.M.; Delage-Mourroux, R.; Lizard, G. Induction of oxiapoptophagy, a mixed mode of cell death associated with oxidative stress, apoptosis and autophagy, on 7-ketocholesterol-treated 158N murine oligodendrocytes: Impairment by α-tocopherol. Biochem. Biophys. Res. Commun. 2014, 446, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Nury, T.; Zarrouk, A.; Mackrill, J.J.; Samadi, M.; Durand, P.; Riedinger, J.M.; Doria, M.; Vejux, A.; Limagne, E.; Delmas, D.; et al. Induction of oxiapoptophagy on 158N murine oligodendrocytes treated by 7-ketocholesterol-, 7β-hydroxycholesterol-, or 24 (S)-hydroxycholesterol: Protective effects of α-tocopherol and docosahexaenoic acid (DHA; C22: 6 n-3). Steroids 2015, 99, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Arcbdelmohsen, K.; Arobe, A.; Arbediasn, E.M.J.; Arbelimotovich, H.; Arioszena, A.R.A.; Armstrongdachi, J.L.H.; Arnouldams, T.C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Accad, M.; Farese, R.V. Cholesterol homeostasis: A role for oxysterols. Curr. Biol. 1998, 8, R601–R604. [Google Scholar] [CrossRef] [Green Version]
- Samadi, A.; Gurlek, A.; Sendur, S.N.; Karahan, S.; Akbiyik, F.; Lay, I. Oxysterol species: Reliable markers of oxidative stress in diabetes mellitus. J. Endocrinol. Investig. 2019, 42, 7–17. [Google Scholar] [CrossRef]
- Liu, Y.; Hulten, L.M.; Wiklund, O. Macrophages isolated from human atherosclerotic plaques produce IL-8, and oxysterols may have a regulatory function for IL-8 production. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 317–323. [Google Scholar] [CrossRef]
- Bourdon, E.; Loreau, N.; Davignon, J.; Bernier, L.; Blache, D. Involvement of oxysterols and lysophosphatidylcholine in the oxidized LDL-induced impairment of serum albumin synthesis by HEPG2 cells. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2643–2650. [Google Scholar] [CrossRef] [Green Version]
- Hanley, K.; Ng, D.C.; He, S.S.; Lau, P.; Min, K.; Elias, P.M.; Bikle, D.D.; Mangelsdorf, D.J.; Williams, M.L.; Feingold, K.R. Oxysterols induce differentiation in human keratinocytes and increase Ap-1-dependent involucrin transcription. J. Investig. Dermatol. 2000, 114, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Lemaire-Ewing, S.; Prunet, C.; Montange, T.; Vejux, A.; Berthier, A.; Besseède, G.; Corcos, L.; Gambert, P.; Neel, D.; Lizalrd, G. Comparison of the cytotoxic, pro-oxidant and pro-inflammatory characteristics of different oxysterols. Cell Biol. Toxicol. 2005, 21, 97–114. [Google Scholar] [CrossRef]
- Vergallo, C. Nutraceutical Vegetable Oil Nanoformulations for Prevention and Management of Diseases. Nanomaterials 2020, 10, 1232. [Google Scholar] [CrossRef]
- Kang, P.; Wang, Y.; Li, X.G.; Wan, Z.C.; Wang, X.Y.; Zhu, H.L.; Wang, C.W.; Zhao, S.J.; Chen, H.F.; Liu, Y.L. Effect of flaxseed oil on muscle protein loss and carbohydrate oxidation impairment in a pig model after lipopolysaccharide challenge. Br. J. Nutr. 2020, 123, 859–869. [Google Scholar] [CrossRef]
- Daoued, K.B.; Chouaibi, M.; Gaout, N.; Haj, O.B.; Hamdi, S. Chemical composition and antioxidant activities of cold pressed lentisc (Pistacialentiscus L.) seed oil. Riv. Grasse 2016, 93, 31–38. [Google Scholar]
- Le Floc’h, E. Contribution à une étude Ethnobotanique de la Flore Tunisienne; Imprimerie Officielle de la République Tunisienne: Radès, Tunisia, 1983. [Google Scholar]
- Ben Khedir, S.; Bardaa, S.; Chabchoub, N.; Moalla, D.; Sahnoun, Z.; Rebai, T. The healing effect of Pistacialentiscus fruit oil on laser burn. Pharm. Biol. 2017, 55, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Saidi, S.A.; Ncir, M.; Chaaben, R.; Jamoussi, K.; van Pelt, J.; Elfeki, A. Liver injury following small intestinal ischemia reperfusion in rats is attenuated by Pistacia lentiscus oil: Antioxidant and anti-inflammatory effects. Arch. Physiol. Biochem. 2017, 123, 199–205. [Google Scholar] [CrossRef]
- Milia, E.; Bullitta, S.M.; Mastandrea, G.; Szotáková, B.; Schoubben, A.; Langhansová, L.; Quartu, M.; Bortonet, A.; Eick, S. Leaves and Fruits Preparations of Pistacialentiscus L.: A Review on the Ethnopharmacological Uses and Implications in Inflammation and Infection. Antibiotics 2021, 10, 425. [Google Scholar]
- Mezni, F.; Shili, S.; Ben Ali, N.; Khouja, M.L.; Khaldi, A.; Maaroufi, A. Evaluation of Pistacialentiscus seed oil and phenolic compounds for in vitro antiproliferative effects against BHK21 cells. Pharm. Biol. 2016, 54, 747–751. [Google Scholar] [CrossRef] [Green Version]
- Ammari, M.; Othman, H.; Hajri, A.; Sakly, M.; Abdelmelek, H. Pistacia lentiscus oil attenuates memory dysfunction and decreases levels of biomarkers of oxidative stress induced by lipopolysaccharide in rats. Brain Res. Bull. 2018, 140, 140–147. [Google Scholar] [CrossRef]
- Trabelsi, H.; Sakouhi, F.; Renaud, J.; Villeneuve, P.; Khouja, M.L.; Mayer, P.; Boukhchina, S. Fatty acids, 4-desmethylsterols, and triterpene alcohols from Tunisian lentisc (Pistacialentiscus) fruits. Eur. J. Lipid Sci. Technol. 2012, 114, 968–973. [Google Scholar] [CrossRef]
- Gimeno, E.; Castellote, A.I.; Lamuela-Raventós, R.M.; De la Torre, M.C.; López-Sabater, M.C. The effects of harvest and extraction methods on the antioxidant content (phenolics, α-tocopherol, and β-carotene) in virgin olive oil. Food Chem. 2002, 78, 207–211. [Google Scholar] [CrossRef]
- Matos, L.C.; Pereira, J.A.; Andrade, P.B.; Seabra, R.M.; Oliveira, M.B.P. Evaluation of a numerical method to predict the polyphenols content in monovarietal olive oils. Food Chem. 2007, 102, 976–983. [Google Scholar] [CrossRef]
- Debbabi, M.; Zarrouk, A.; Bezine, M.; Meddeb, W.; Nury, T.; Badreddine, A.; Sghaier, R.; Bretillon, L.; Guyot, S.; Samadi, M.; et al. Comparison of the effects of major fatty acids present in the Mediterranean diet (oleic acid, docosahexaenoic acid) and in hydrogenated oils (elaidic acid) on 7-ketocholesterol-induced oxiapoptophagy in microglial BV-2 cells. Chem. Phys. Lipids 2017, 207, 151–170. [Google Scholar] [CrossRef]
- Yammine, A.; Nury, T.; Vejux, A.; Latruffe, N.; Vervandier-Fasseur, D.; Samadi, M.; Gretige-Gerges, H.; Auezova, L.; Lizard, G. Prevention of 7-Ketocholesterol-Induced Overproduction of Reactive Oxygen Species, Mitochondrial Dysfunction and Cell Death with Major Nutrients (Polyphenols, ω3 and ω9 Unsaturated Fatty Acids) of the Mediterranean Diet on N2a Neuronal Cells. Molecules 2020, 25, 2296. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Kim, D.O.; Chun, O.K.; Kim, Y.J.; Moon, H.Y.; Lee, C.Y. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef]
- Dhibi, M.; Issaoui, M.; Brahmi, F.; Mechri, B.; Mnari, A.; Cheraif, I.; etSkhiri, F.; Galzzah, N.; Hammami, M. Nutritional quality of fresh and heated Aleppo pine (Pinus halepensis Mill.) seed oil: Trans-fatty acid isomers profiles and antioxidant properties. J. Food Sci. Technol. 2014, 51, 1442–1452. [Google Scholar] [CrossRef] [Green Version]
- Moilanen, T.; Nikkari, T. The effect of storage on the fatty acid composition of human serum. Clin. Chim. Acta 1981, 114, 111–116. [Google Scholar] [CrossRef]
- Morrison, W.R.; Smith, L.M. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride–methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [CrossRef]
- Zarrouk, A.; Martine, L.; Grégoire, S.; Nury, T.; Meddeb, W.; Camus, E.; Badreddine, A.; Durand, P.; Namsi, A.; Yammine, A.; et al. Profile of fatty acids, tocopherols, phytosterols and polyphenols in mediterranean oils (argan oils, olive oils, milk thistle seed oils and nigella seed oil) and evaluation of their antioxidant and cytoprotective activities. Curr. Pharm. Des. 2019, 25, 1791–1805. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin. J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Bassene, E. Initiation à la Recherche sur les Substances Naturelles: Extraction-Analyse-Essais Biologiques; Presses Universitaires de Dakar: Dakar, Sénégal, 2012. [Google Scholar]
- Dinis, T.C.; Madeira, V.M.; Almeida, L.M. Action of phenolic derivatives (acetaminophen, salicylate, and 5-aminosalicylate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Rossi, R.; Pastorelli, G.; Corino, C. Application of KRL test to assess total antioxidant activity in pigs: Sensitivity to dietary antioxidants. Res. Vet. Sci. 2013, 94, 372–377. [Google Scholar] [CrossRef]
- Ragot, K.; Mackrill, J.J.; Zarrouk, A.; Nury, T.; Aires, V.; Jacquin, A.; Athias, A.; de Barros, J.-P.P.; Vejux, A.; Riedinger, J.-M.; et al. Absence of correlation between oxysterol accumulation in lipid raft microdomains, calcium increase, and apoptosis induction on 158N murine oligodendrocytes. Biochem. Pharm. 2013, 86, 67–79. [Google Scholar] [CrossRef]
- Jones, K.H.; Senft, J.A. An improved method to determine cell viability by simultaneous staining with fluorescein diacetate-propidium iodide. J. Histochem. Cytochem. 1985, 33, 77–79. [Google Scholar] [CrossRef] [Green Version]
- Patková, J.; Anděl, M.; Trnka, J. Palmitate-induced cell death and mitochondrial respiratory dysfunction in myoblasts are not prevented by mitochondria-targeted antioxidants. Cell Physiol. Biochem. 2014, 33, 1439–1451. [Google Scholar] [CrossRef]
- Cui, L.; Zhou, Q.; Zheng, X.; Sun, B.; Zhao, S. Mitoquinone attenuates vascular calcification by suppressing oxidative stress and reducing apoptosis of vascular smooth muscle cells via the Keap1/Nrf2 pathway. Free Radic. Biol. Med. 2020, 161, 23–31. [Google Scholar] [CrossRef]
- Yeh, C.J.; His, B.L.; Faulk, W.P. Propidium iodide as a nuclear marker in immunofluorescence. II. Use with cellular identification and viability studies. J. Immunol Methods 1981, 43, 269–275. [Google Scholar] [CrossRef]
- Rothe, G.; Valet, G. Flow cytometric analysis of respiratory burst activity in phagocytes with hydroethidine and 2′, 7′-dichlorofluorescin. J. Leukoc. Biol. 1990, 47, 440–448. [Google Scholar] [CrossRef]
- Flohe, L.; Günzler, W.A. Assays of glutathione peroxidase. Methods Enzym. 1984, 105, 114–120. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Yoshioka, T.; Kawada, K.; Shimada, T.; Mori, M. Lipid peroxidation in maternal and cord blood and protective mechanism against activated-oxygen toxicity in the blood. Am. J. Obstet. Gynecol. 1979, 135, 372–376. [Google Scholar] [CrossRef]
- Esterbauer, H.; Striegl, G.; Puhl, H.; Rotheneder, M. Continuous monitoring of in vitro oxidation of human low density lipoprotein. Free Radic. Res. Commun. 1989, 6, 67–75. [Google Scholar] [CrossRef]
- Oliver, C.N.; Ahn, B.W.; Moerman, E.J.; Goldstein, S.; Stadtman, E.R. Age-related changes in oxidized proteins. J. Biol. Chem. 1987, 262, 5488–5491. [Google Scholar] [CrossRef]
- Zarrouk, A.; Vejux, A.; Nury, T.; El Hajj, H.I.; Haddad, M.; Cherkaoui-Malki, M.; Rietdinger, J.-M.; Hammami, M.; Lizard, G. Induction of mitochondrial changes associated with oxidative stress on very long chain fatty acids (C22: 0, C24: 0, or C26: 0)-treated human neuronal cells (SK-NB-E). Oxidative Med. Cell Longev. 2012, 2012, 623257. [Google Scholar] [CrossRef] [Green Version]
- Gray, E.; Rice, C.; Hares, K.; Redondo, J.; Kemp, K.; Williams, M.; etBrown, A.; Scolding, N.; Wilkins, A. Reductions in neuronal peroxisomes in multiple sclerosis grey matter. Mult. Scler. J. 2014, 20, 651–659. [Google Scholar] [CrossRef]
- Nury, T.; Sghaier, R.; Zarrouk, A.; Ménétrier, F.; Uzun, T.; Leoni, V.; Caccia, C.; Meddeb, W.; Namsi, A.; Sassi, K.; et al. Induction of peroxisomal changes in oligodendrocytes treated with 7-ketocholesterol: Attenuation by α-tocopherol. Biochimie 2018, 153, 181–202. [Google Scholar] [CrossRef]
- Savary, S.; Trompier, D.; Andréoletti, P.; Le Borgne, F.; Demarquoy, J.; Lizard, G. Fatty acids-induced lipotoxicity and inflammation. Curr. Drug Metab. 2012, 13, 1358–1370. [Google Scholar] [CrossRef]
- Blondelle, J.; de Barros, J.P.P.; Pilot-Storck, F.; Tiret, L. Targeted lipidomic analysis of myoblasts by GC-MS and LC-MS/MS. In Skeletal Muscle Development; Humana Press: New York, NY, USA, 2017; pp. 39–60. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Lizard, G.; Fournel, S.; Genestier, L.; Dhedin, N.; Chaput, C.; Flacher, M.; Mutin, M.; Panayet, G.; Revillalrd, J.-P. Kinetics of plasma membrane and mitochondrial alterations in cells undergoing apoptosis. Cytom. J. Int. Soc. Anal. Cytol. 1995, 21, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Baarine, M.; Ragot, K.; Genin, E.C.; El Hajj, H.; Trompier, D.; Andreoletti, P.; Ghandour, S.; Menetrier, F.; Cherkaoui-Malki, M.; Savary, S.; et al. Peroxisomal and mitochondrial status of two murine oligodendrocytic cell lines (158N, 158JP): Potential models for the study of peroxisomal disorders associated with dysmyelination processes. J. Neurochem. 2009, 111, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badreddine, A.; Zarrouk, A.; Karym, E.M.; Debbabi, M.; Nury, T.; Meddeb, W.; Sghaier, R.; Bezine, M.; Vejux, A.; Martine, L.; et al. Argan oil-mediated attenuation of organelle dysfunction, oxidative stress and cell death induced by 7-ketocholesterol in murine oligodendrocytes 158N. Int. J. Mol. Sci. 2017, 18, 2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sottero, B.; Rossin, D.; Staurenghi, E.; Gamba, P.; Poli, G.; Testa, G. Omics analysis of oxysterols to better understand their pathophysiological role. Free Radic. Biol. Med. 2019, 144, 55–71. [Google Scholar] [CrossRef]
- Fujita, T.; Adachi, J.; Ueno, Y.; Peters, T.J.; Preedy, V.R. Chronic ethanol feeding increases 7-hydroperoxycholesterol and oxysterols in rat skeletal muscle. Metab. -Clin. Exp. 2002, 51, 737–742. [Google Scholar] [CrossRef]
- Jamadagni, P.; Patten, S.A. 25-hydroxycholesterol impairs neuronal and muscular development in zebrafish. Neurotoxicology 2019, 75, 14–23. [Google Scholar] [CrossRef]
- Nishi, H.; Higashihara, T.; Inagi, R. Lipotoxicity in kidney, heart, and skeletal muscle dysfunction. Nutrients 2019, 11, 1664. [Google Scholar] [CrossRef] [Green Version]
- Kemp, S.; Theodoulou, F.L.; Wanders, R.J. Mammalian peroxisomal ABC transporters: From endogenous substrates to pathology and clinical significance. Br. J. Pharmacol. 2011, 164, 1753–1766. [Google Scholar] [CrossRef] [Green Version]
- Tawbeh, A.; Gondcaille, C.; Trompier, D.; Savary, S. Peroxisomal ABC Transporters: An Update. Int. J. Mol. Sci. 2021, 22, 6093. [Google Scholar] [CrossRef]
- Trompier, D.; Vejux, A.; Zarrouk, A.; Gondcaille, C.; Geillon, F.; Nury, T.; etSavary, S.; Lizalrd, G. Brain peroxisomes. Biochimie 2014, 98, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef]
- Kihara, A. Very long-chain fatty acids: Elongation, physiology and related disorders. J. Biochem. 2021, 152, 387–395. [Google Scholar] [CrossRef]
- Musaro, A.; Fulle, S.; Fano, G. Oxidative stress and muscle homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 236–242. [Google Scholar] [CrossRef]
- Powers, S.K.; Talbert, E.E.; Adhihetty, P.J. Reactive oxygen and nitrogen species as intracellular signals in skeletal muscle. J. Physiol. 2011, 589, 2129–2138. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Pistollato, F.; Giampieri, F.; Battino, M. The use of plant-derived bioactive compounds to target cancer stem cells and modulate tumor microenvironment. Food Chem. Toxicol. 2015, 75, 58–70. [Google Scholar] [CrossRef]
- Gaussen, H.; Leroy, J.F.; Ozenda, P. Précis de Botanique. 2- Végétaux Supérieurs, 2nd ed.; Editions Masson: Paris, France, 1982; pp. 307–308. [Google Scholar]
- Fazeli-nasab, B.; Fooladvand, Z. Classification and Evaluation of medicinal plant and medicinal properties of mastic. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 2155–2161. [Google Scholar]
- Brahmi, F.; Haddad, S.; Bouamara, K.; Yalaoui-Guellal, D.; Prost-Camus, E.; De Barros, J.P.P.; Prost, M.; Atanasov, A.G.; Madani, K.; Boulekbache-Makhlouf, L.; et al. Comparison of chemical composition and biological activities of Algerian seed oils of Pistacialentiscus L., Opuntia ficusindica (L.) mill. and Argania spinosa L. Skeels. Ind. Crop. Products 2020, 151, 112456. [Google Scholar] [CrossRef]
- Bouyahya, A.; Dakka, N.; Talbaoui, A.; El Moussaoui, N.; Abrini, J.; Bakri, Y. Phenolic contents and antiradical capacity of vegetable oil from Pistacialentiscus (L). J. Mater. Environ. Sci. 2018, 9, 1518–1524. [Google Scholar]
- Maria, A.G.; Graziano, R.; Nicolantonio, D.O. Carotenoids: Potential allies of cardiovascular health? Food Nutr. Res. 2015, 59, 26762. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.A. Carotenoids and cancer: Prevention and potential therapy. Br. J. Biomed. Sci. 1998, 55, 268. [Google Scholar]
- Kitakaze, T.; Harada, N.; Imagita, H.; Yamaji, R. β-Carotene increases muscle mass and hypertrophy in the soleus muscle in mice. J. Nutr. Sci. Vitaminol. 2015, 61, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Dhifi, W.; Jelali, N.; Chaabani, E.; Beji, M.; Fatnassi, S.; Omri, S.; Mnif, W. Chemical composition of Lentisk (Pistacialentiscus L.) seed oil. Afr. J. Agric. Res. 2013, 8, 1395–1400. [Google Scholar]
- Aguilera, C.M.; Ramírez-Tortosa, M.C.; Mesa, M.D.; Gil, A. Do MUFA and PUFA have beneficial effects on the development of cardiovascular disease? Recent Res. Dev. Lipids 2000, 4, 369–390. [Google Scholar]
- Debbabi, M.; Nury, T.; Zarrouk, A.; Mekahli, N.; Bezine, M.; Sghaier, R.; Grégoire, S.; Martine, L.; Durand, P.; Prost, E.; et al. Protective effects of α-tocopherol, γ-tocopherol and oleic acid, three compounds of olive oils, and no effect of trolox, on 7-ketocholesterol-induced mitochondrial and peroxisomal dysfunction in microglial BV-2 cells. Int. J. Mol. Sci. 2016, 17, 1973. [Google Scholar] [CrossRef]
- Jandacek, R.J. Linoleic Acid: A Nutritional Quandary. Healthcare 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Tachibana, H.; Morinaga, Y.; Fujimura, Y.; Yamada, K. Modulation of proliferation and differentiation of C2C12 skeletal muscle cells by fatty acids. Life Sci. 2009, 84, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Charef, M.; Yousfi, M.; Saidi, M.; Stocker, P. Determination of the fatty acid composition of acorn (Quercus), Pistacialentiscus seeds growing in Algeria. J. Am. Oil Chem. Soc. 2008, 85, 921–924. [Google Scholar] [CrossRef]
- French, M.A.; Sundram, K.; Clandinin, M.T. Cholesterolaemic effect of palmitic acid in relation to other dietary fatty acids. Asia Pac. J. Clin. Nutr. 2002, 11, S401–S407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.-L.; Kim, H.-N.; Jung, H.-H.; Kim, J.-E.; Choi, D.-K.; Hur, J.-M.; Lee, J.-Y.; Song, H.; Song, K.-S.; Huh, T.-L. Beneficial effects of β-sitosterol on glucose and lipid metabolism in L6 myotube cells are mediated by AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2008, 377, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.-C.; Lai, M.-H.; Hsu, K.-P.; Kuo, Y.-H.; Chen, J.; Tsai, M.-C.; Li, C.-X.; Yin, X.-J.; Jetyashoke, N.; Chao, L.K.-P. Identification of β-sitosterol as in vitro anti-inflammatory constituent in Moringa oleifera. J. Agric. Food Chem. 2018, 66, 10748–10759. [Google Scholar] [CrossRef]
- Gupta, R.; Sharma, A.K.; Dobhal, M.P.; Sharma, M.C.; Gupta, R.S. Antidiabetic and antioxidant potential of β-sitosterol in streptozotocin-induced experimental hyperglycemia. J. Diabetes 2011, 3, 29–37. [Google Scholar] [CrossRef]
- Wong, H.S.; Leong, P.K.; Chen, J.; Leung, H.Y.; Chan, W.M.; Ko, K.M. β-Sitosterol increases mitochondrial electron transport by fluidizing mitochondrial membranes and enhances mitochondrial responsiveness to increasing energy demand by the induction of uncoupling in C2C12 myotubes. J. Funct. Foods 2016, 23, 253–260. [Google Scholar] [CrossRef]
- Chen, Y. Endogenous Phenolics from Expeller-Pressed Canola Oil Refining Byproducts: Evaluation of Antioxidant Activities in Cell Culture and Deep-Fat Frying Models. Master’s Thesis, University of Manitoba, Winnipeg, MB, Canada, 2014. [Google Scholar]
- Zingg, J.M.; Meydani, M. Interaction between vitamin E and polyunsaturated fatty acids. In Vitamin E in Human Health; Humana Press: Cham, Switzerland, 2019; pp. 141–159. [Google Scholar]
- Rimbach, G.; Minihane, A.M.; Majewicz, J.; Fischer, A.; Pallauf, J.; Virgli, F.; Weinberg, P.D. Regulation of cell signalling by vitamin E. Proc. Nutr. Soc. 2002, 61, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Testa, G.; Rossin, D.; Poli, G.; Biasi, F.; Leonarduzzi, G. Implication of oxysterols in chronic inflammatory human diseases. Biochimie 2018, 153, 220–231. [Google Scholar] [CrossRef]
- Pedruzzi, E.; Guichard, C.; Ollivier, V.; Driss, F.; Fay, M.; Prunet, C.; Marie, J.-C.; Pouzet, C.; Samadi, M.; Elbim, C.; et al. NAD (P) H oxidase Nox-4 mediates 7-ketocholesterol-induced endoplasmic reticulum stress and apoptosis in human aortic smooth muscle cells. Mol. Cell. Biol. 2004, 24, 10703–10717. [Google Scholar] [CrossRef] [Green Version]
- Szentesi, P.; Csernoch, L.; Dux, L.; Keller-Pintér, A. Changes in redox signaling in the skeletal muscle with aging. Oxidative Med. Cell Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Nury, T.; Zarrouk, A.; Yammine, A.; Mackrill, J.J.; Vejux, A.; Lizard, G. Oxiapoptophagy: A type of cell death induced by some oxysterols. Br. J. Pharmacol. 2021, 178, 3115–3123. [Google Scholar] [CrossRef]
- Sghaier, R.; Nury, T.; Leoni, V.; Caccia, C.; De Barros, J.-P.P.; Cherif, A.; Vejux, A.; Moreau, T.; Limem, K.; Samadi, M.; et al. Dimethyl fumarate and monomethyl fumarate attenuate oxidative stress and mitochondrial alterations leading to oxiapoptophagy in 158N murine oligodendrocytes treated with 7β-hydroxycholesterol. J. Steroidbiochemistry Mol. 2019, 194, 105432. [Google Scholar] [CrossRef]
- Yammine, A.; Zarrouk, A.; Nury, T.; Vejux, A.; Latruffe, N.; Vervandier-Fasseur, D.; Samadi, M.; Mackrill, J.J.; Greige-Gerges, H.; Auezova, L.; et al. Prevention by dietary polyphenols (resveratrol, quercetin, apigenin) against 7-ketocholesterol-induced oxiapoptophagy in neuronal N2a cells: Potential interest for the treatment of neurodegenerative and age-related diseases. Cells 2020, 9, 2346. [Google Scholar] [CrossRef]
- Poli, G.; Biasi, F.; Leonarduzzi, G. Oxysterols in the pathogenesis of major chronic diseases. Redox Biol. 2013, 1, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Kunzelmann, K. Ion channels in regulated cell death. Cell Mol. Life Sci. 2016, 73, 2387–2403. [Google Scholar] [CrossRef]
- Bezine, M.; Namsi, A.; Sghaier, R.; Ben Khalifa, R.B.; Hamdouni, H.; Brahmi, F.; Badreddine, I.; Mihoubi, W.; Nury, T.; Vejux, A.; et al. The effect of oxysterols on nerve impulses. Biochimie 2018, 153, 46–51. [Google Scholar] [CrossRef]
- Zarrouk, A.; Ben Salem, Y.B.; Hafsa, J.; Sghaier, R.; Charfeddine, B.; Limem, K.; etHammami, M.; Maljdoub, H. 7β-hydroxycholesterol-induced cell death, oxidative stress, and fatty acid metabolism dysfunctions attenuated with sea urchin egg oil. Biochimie 2018, 153, 210–219. [Google Scholar] [CrossRef]
- Rufini, A.; Tucci, P.; Celardo, I.; Melino, G. Senescence and aging: The critical roles of p53. Oncogene 2013, 32, 5129–5143. [Google Scholar] [CrossRef]
- Lismont, C.; Nordgren, M.; Van Veldhoven, P.P.; Fransen, M. Redox interplay between mitochondria and peroxisomes. Front. Cell Dev. Biol. 2015, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Pascual-Ahuir, A.; Manzanares-Estreder, S.; Proft, M. Pro-and antioxidant functions of the peroxisome-mitochondria connection and its impact on aging and disease. Oxidative Med. Cell Longev. 2017, 2017, 9860841. [Google Scholar] [CrossRef] [Green Version]
- Baarine, M.; Andreoletti, P.; Athias, A.; Nury, T.; Zarrouk, A.; Ragot, K. Evidence of oxidative stress in very long chain fatty acid–treated oligodendrocytes and potentialization of ROS production using RNA interference-directed knockdown of ABCD1 and ACOX1 peroxisomal proteins. Neuroscience 2012, 213, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wanders, R.J. Metabolic functions of peroxisomes in health and disease. Biochimie 2014, 98, 36–44. [Google Scholar] [CrossRef]
- Titorenko, V.I.; Terlecky, S.R. Peroxisome metabolism and cellular aging. Traffic 2011, 12, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Unger, R.H.; Orci, L. Lipoapoptosis: Its mechanism and its diseases. Biochim. Et Biophys. Acta -Mol. Cell Biol. Lipids 2002, 1585, 202–212. [Google Scholar] [CrossRef]
- Amery, L.; Fransen, M.; De Nys, K.; Mannaerts, G.P.; Van Veldhoven, P.P. Mitochondrial and peroxisomal targeting of 2-methylacyl-CoA racemase in humans. J. Lipid Res. 2000, 41, 1752–1759. [Google Scholar] [CrossRef]
- Cecatto, C.; Amaral, A.U.; Roginski, A.C.; Castilho, R.F.; Wajner, M. Impairment of mitochondrial bioenergetics and permeability transition induction caused by major long-chain fatty acids accumulating in VLCAD deficiency in skeletal muscle as potential pathomechanisms of myopathy. Toxicol. Vitr. 2020, 62, 104665. [Google Scholar] [CrossRef] [PubMed]
- Ben Khedir, S.; Moalla, D.; Jardak, N.; Mzid, M.; Sahnoun, Z.; Rebai, T. Pistacia lentiscus fruit oil reduces oxidative stress in human skin explants caused by hydrogen peroxide. Biotech. Histochem. 2016, 91, 480–491. [Google Scholar] [CrossRef]
- Kang, J.S.; Han, M.H.; Kim, G.-Y.; Kim, C.M.; Chung, H.Y.; Hwang, H.J.; Kim, B.W.; Choi, Y.H. Schisandrae semen essential oil attenuates oxidative stress-induced cell damage in C2C12 murine skeletal muscle cells through Nrf2-mediated upregulation of HO-1. Int. J. Mol. 2015, 35, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H. Protective effects of isorhamnetin against hydrogen peroxide-induced apoptosis in c2c12 murine myoblasts. J. Korean Med. Obes. Res. 2015, 15, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Leporini, L.; Giampietro, L.; Amoroso, R.; Ammazzalorso, A.; Fantacuzzi, M.; Menghini, L.; Maccallini, C.; Ferrante, C.; Brunetti, L.; Orlando, G.; et al. In vitro protective effects of resveratrol and stilbene alkanoic derivatives on induced oxidative stress on C2C12 and MCF7 cells. J. Biol. Regul. Homeost. Agents 2017, 31, 589–601. [Google Scholar]
- Li, J.; Yang, Q.; Han, L.; Pan, C.; Lei, C.; Chen, H.; Lan, X. C2C12 mouse myoblasts damage induced by oxidative stress is alleviated by the antioxidant capacity of the active substance phloretin. Front. Cell Dev. Biol. 2020, 8, 953. [Google Scholar] [CrossRef]
- Badreddine, A.; Zarrouk, A.; Meddeb, W.; Nury, T.; Rezig, L.; Debbabi, M.; Bessam, F.Z.; Brahmi, F.; Vejux, A.; Mejri, M.; et al. Antioxidant and neuroprotective properties of Mediterranean Oils: Argan Oil, Olive Oil, and Milk Thistle Seed Oil. In Oxidative Stress and Dietary Antioxidants in Neurological Diseases; Elsevier: Amsterdam, The Netherlands, 2020; pp. 143–154. [Google Scholar]
- Brahmi, F.; Vejux, A.; Sghaier, R.; Zarrouk, A.; Nury, T.; Meddeb, W.; Rezig, L.; Namsi, A.; Sassi, K.; Yammine, A.; et al. Prevention of 7-ketocholesterol-induced side effects by natural compounds. Crit. Rev. Food Sci. Nutr. 2019, 59, 3179–3198. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Phenols (mg GAE/g of Extract) | Flavonoids (mg CE/g of Extract) | Carotenoids (mg/kg) | |
|---|---|---|---|
| Pistacia lentiscus seed oil | 28.50 ± 0.77 | 51.36 ± 2.30 | 2083.59 ± 55.00 |
| Polyphenols | (mg Equivalents Quercetin/100 g of Oil) |
|---|---|
| Protocatechuic acid | 0.140 ± 0.001 |
| Coumarin | 0.650 ± 0.003 |
| Fatty Acids | g/100 g of Oil (*) | % (**) |
|---|---|---|
| ∑SFA | 32.333 ± 0.566 | 28.77 ± 0.24 |
| Myristic acid (C14:0) | 0.015 ± 0.000 | 0.05 ± 0.01 |
| Palmitic acid (C16:0) | 30.457 ± 0.355 | 27.20 ± 0.22 |
| Margaric acid (C17:0) | 0.022 ± 0.003 | 0.05 ± 0.00 |
| Stearic acid (C18:0) | 1.527 ± 0.124 | 1.26 ± 0.01 |
| Arachidic acid (C20:0) | 0.041 ± 0.012 | 0.12 ± 0.00 |
| Behenic acid (C22:0) | 0.271 ± 0.072 | 0.04 ± 0.00 |
| Lignoceric acid (C24:0) | ND | 0.05 ± 0.00 |
| ∑UFA | 94.706 ± 2.910 | 71.25 ± 0.28 |
| ∑MUFA | 64.736 ± 1.827 | 53.67 ± 0.17 |
| Palmitoleic acid (C16:1 n−7) | 2.623 ± 0.084 | 2.25 ± 0.01 |
| Heptadecenoic acid (C17:1) | 0.179 ± 0.058 | ND |
| Oleic acid (C18:1 n−9) | 60.183 ± 1.556 | 49.77 ± 0.12 |
| Vaccenic acid (C18:1 n−7) | 1.670 ± 0.073 | 1.51 ± 0.03 |
| Gadoleic acid (C20:1 n−9) | 0.081 ± 0.056 | 0.14 ± 0.01 |
| ∑PUFA | 29.970 ± 1.083 | 17.58 ± 0.11 |
| Linoleic acid (C18:2 n−6) | 29.665 ± 0.990 | 17.19 ± 0.10 |
| α-linolenic acid (C18:3 n−3) | 0.305 ± 0.093 | 0.39 ± 0.01 |
| ∑SFA/∑UFA | 0.34 ± 0.19 | 0.41 ± 0.86 |
| Phytosterol | (mg/100 g of Oil) |
|---|---|
| Campesterol | 4.48 ± 0.85 |
| Stigmasterol | 2.69 ± 0.23 |
| β-Sitosterol | 67.25 ± 3.24 |
| ∆5-Avenasterol | 3.10 ± 0.90 |
| β-Amyrine | 1.81 ± 0.19 |
| Cycloartenol | 9.35 ± 1.49 |
| 24-Methylene cycloartenol | 16.10 ± 2.72 |
| α-Epoxysitostanol | 33.36 ± 1.65 |
| Other phytosterols | 16.76 ± 2.83 |
| Total | 154.89 ± 5.40 |
| α-Tocopherol (mg/kg) | |
|---|---|
| Pistacia lentiscusseed oil | 68.10 ± 3.41 |
| IC50 Values (mg/mL) | (Trolox Equivalent) | |||
|---|---|---|---|---|
| Samples | DPPH | FRAP | Iron Chelating (FIC) | KRL |
| PLSO | 5.010 ± 0.095 | 1.15 ± 0.23 | 5.61 ± 0.14 | 4440.00 ± 493.60 |
| AO | − | − | − | 360.80 ± 153.60 |
| EDTA (standard) | − | − | 0.60 ± 0.09 | − |
| AA (standard) | 0.810 ± 0.270 | 0.41 ± 0.16 | − | − |
| α-tocopherol | − | − | − | 0.94 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghzaiel, I.; Zarrouk, A.; Nury, T.; Libergoli, M.; Florio, F.; Hammouda, S.; Ménétrier, F.; Avoscan, L.; Yammine, A.; Samadi, M.; et al. Antioxidant Properties and Cytoprotective Effect of Pistacia lentiscus L. Seed Oil against 7β-Hydroxycholesterol-Induced Toxicity in C2C12 Myoblasts: Reduction in Oxidative Stress, Mitochondrial and Peroxisomal Dysfunctions and Attenuation of Cell Death. Antioxidants 2021, 10, 1772. https://doi.org/10.3390/antiox10111772
Ghzaiel I, Zarrouk A, Nury T, Libergoli M, Florio F, Hammouda S, Ménétrier F, Avoscan L, Yammine A, Samadi M, et al. Antioxidant Properties and Cytoprotective Effect of Pistacia lentiscus L. Seed Oil against 7β-Hydroxycholesterol-Induced Toxicity in C2C12 Myoblasts: Reduction in Oxidative Stress, Mitochondrial and Peroxisomal Dysfunctions and Attenuation of Cell Death. Antioxidants. 2021; 10(11):1772. https://doi.org/10.3390/antiox10111772
Chicago/Turabian StyleGhzaiel, Imen, Amira Zarrouk, Thomas Nury, Michela Libergoli, Francesca Florio, Souha Hammouda, Franck Ménétrier, Laure Avoscan, Aline Yammine, Mohammad Samadi, and et al. 2021. "Antioxidant Properties and Cytoprotective Effect of Pistacia lentiscus L. Seed Oil against 7β-Hydroxycholesterol-Induced Toxicity in C2C12 Myoblasts: Reduction in Oxidative Stress, Mitochondrial and Peroxisomal Dysfunctions and Attenuation of Cell Death" Antioxidants 10, no. 11: 1772. https://doi.org/10.3390/antiox10111772
APA StyleGhzaiel, I., Zarrouk, A., Nury, T., Libergoli, M., Florio, F., Hammouda, S., Ménétrier, F., Avoscan, L., Yammine, A., Samadi, M., Latruffe, N., Biressi, S., Levy, D., Bydlowski, S. P., Hammami, S., Vejux, A., Hammami, M., & Lizard, G. (2021). Antioxidant Properties and Cytoprotective Effect of Pistacia lentiscus L. Seed Oil against 7β-Hydroxycholesterol-Induced Toxicity in C2C12 Myoblasts: Reduction in Oxidative Stress, Mitochondrial and Peroxisomal Dysfunctions and Attenuation of Cell Death. Antioxidants, 10(11), 1772. https://doi.org/10.3390/antiox10111772