
Clinical Significance of Heme Oxygenase 1 in Tumor Progression


Abstract
1. Introduction
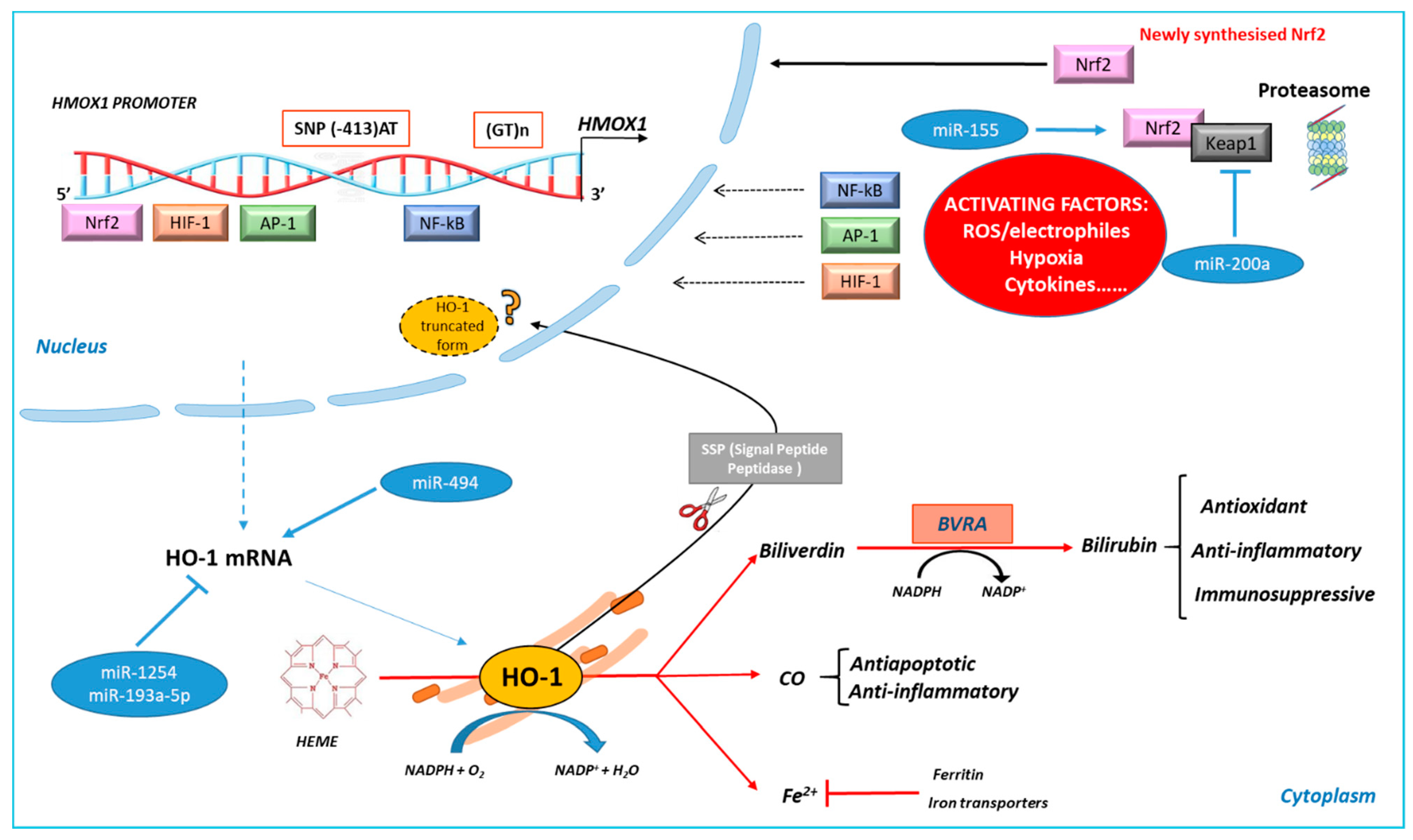
2. HO-1 Gene Transcription and Protein Localization
2.1. HO-1 Transcriptional and Post-Transcriptional Regulation
2.2. HO-1 Sub-Cellular and Extra-Cellular Localization
3. Role of HO-1 in Cancer Progression
3.1. HO-1 in Cancer Cell Growth, Metastasis, and Angiogenesis
3.2. HO-1 in Cancer Immune Escape
3.3. HO-1 in the Resistance to Therapy
4. HO-1 Promoter Polymorphisms and Cancer Risk
5. HO-1 Expression, Tumor Aggressiveness, and Disease Outcome. Evidence from Immunohistochemistry
5.1. HO-1 Expression and Disease Outcome
5.2. HO-1 Expression and Tumor Grade/Stage
5.3. Correlation between HO-1 Expression and Tumor Markers
5.4. Contrasting Evidence

{kind=link}
{kind=link}
| Tumor | HO-1 | Nrf2 | Grade and Stage | Additional Markers | Metastasis, Lymph Node, Angiogenesis | Clinical and Pathological Features | Disease Outcome/Prognosis | Ref. |
|---|---|---|---|---|---|---|---|---|
| Positive correlation among HO-1 expression and tumor aggressiveness/poor prognosis | ||||||||
| -Solid tumors | ||||||||
| Astrocytoma | High level | n.e. | Grade II and III | n.e. | n.e. | n.e. | Poor OS | [163] |
| Clear cell renal cell carcinoma | High level | High level | No correlation with ISUP grade and T stage | n.e. | No correlation with lymph node metastasis | No significant correlation with age, gender | Poor prognosis Low MST Low post operative OS | [165] |
| Colangiocarcinoma | High level | n.e. | n.e. | n.e. | No association with metastasis | No significant association with age, gender, histological type | Poor OS | [166] |
| Gastric cancer | High level | High level | Poor differentiated tumors | Positive correlation with VEGF | Positive correlation with MVD | n.e. | n.e. | [174] |
| Gallbladder cancer | High level | High level | Moderately differentiated and poorly differentiated tumors (G2-G3) Correlation with Nevin classification (III-IV-V) | Positive correlation with MRP3 | Metastasis | No significant correlation with gender, age, and histology type (SCC and AD) | Poor OS | [164] |
| Hepatocellular carcinoma | High level | n.e. | Poor differentiated tumors Edmondson-Steiner grade 2–4 | n.e. | Microvascular and capsular invasion | High levels of preoperative AFP | No significant correlation with OS and recurrence | [175] |
| Hormone refractory prostate cancer | High level | n.e. | n.e. | n.e. | n.e. | Cancer progression | n.e. | [188] |
| Laryngeal cancer | High level | High level | No correlation with tumor stage (clinical stage III and IV), size tumor | High level Keap1 and NQO1 | No correlation with lymph node metastasis | No correlation with age | n.e. | [189] |
| Melanoma | High level | n.e. | n.e. | Positive correlation with B-Raf and ERK | n.e. | n.e. | n.e. | [176] |
| Neuroblastoma | High level | n.e. | n.e. | n.e. | n.e. | n.e. | Poor OS | [168] |
| Non-muscle-invasive bladder cancer | High level | n.e. | Tumor grade G3 tumor stage pT1 | Ki-67 and p53 | n.e. | No significant correlation with age and gender | Poor prognosis No correlation with RFS and PFS | [161] |
| High level | n.e. | Tumor grade G3 Tumor stage T1 | Positive correlation with S100A4 | Lymph vascular invasion | n.e. | Low RFS Low PFS | [162] | |
| High level | n.e. | n.e. | Positive correlation with HIF-1α | High MVD | n.e. | n.e. | [123] | |
| High level | High level | n.e. | Correlation with HIF-1α, HIF-2α, VEGF | n.e. | Increased serum/plasma level of IL-6, IL-8, VEGF | n.e. | [124] | |
| Non-small cell lung cancer | High level | n.e. | Stage III-IV | Positive correlation with MMP-9 | High metastatic rate | No correlation with age and gender | Poor prognosis Low OS High mortality risk | [118] |
| High level | n.e. | Stage III-IV T status (T3-T4) | n.e. | Lymph node metastasis | No correlation with gender | No significant difference in patient survival between high and low staining group | [98] | |
| Ovarian cancer | High level | n.e. | Serous undifferentiated tumors Correlation with FIGO stage (III-IV) | n.e. | Lymph node metastasis | Non optimal-debulking | Poor OS | [160] |
| Prostate cancer | High level | n.e. | Localized tumor | PTEN deletion | n.e. | n.e. | Relapse after radical prostatectomy | [173] |
| Thyroid cancer | High level | n.e. | Positive correlation with TNM (1,2,3,4) and with MACIS score | BRAFV600E mutation | No significant association with lymph node metastasis | Correlation with age and tumor aggressiveness | n.e. | [172] |
| -Hematopoietic tumors | ||||||||
| Acute myeloid leukemia | High level | n.e. | n.e. | Positive correlation with HIF-1α and GLUT-1 | n.e. | n.e. | Correlation with relapse and refractory | [170] |
| High level | n.e. | Correleation with M5 patients | Correlation with RET gene | n.e. | Correlation with leukocytosis at diagnosis | n.e. | [167] | |
| Chronic myeloid leukemia | Higher level in peripheral blood cells | n.e. | n.e. | n.e. | n.e. | Tumor progression | Correlation with relapse | [169] |
| Myelodysplastic Syndrome | High level | n.e. | Correlation with high-risk and very high-risk patients | Positive correlation with EZH2 | n.e. | Progression to AML and decreased response to decitabine | n.e. | [171] |
| Positive correlation among HO-1 expression in tumor-associated cells and tumor aggressiveness/poor prognosis | ||||||||
| Colorectal cancer | High level in cancer cells and in macrophages | n.e. | Stage III | n.e. | Lymph node metastasis | No significant difference between the HO-1-positive and negative with gender, age, tumor size, histological type, and depth of tumor invasion | Poor prognosis Short DSF | [179] |
| Glioblastoma | High level in infiltrating macrophages | n.e. | Grade IV | n.e. | Positive correlation with vascular density | n.e. | n.e. | [178] |
| Glioma | HO-1 positive Treg | n.e. | Correlation with grade glioma (II-III-IV) | n.e. | n.e. | n.e. | n.e. | [137] |
| Non-muscle-invasive bladder cancer | High level in cancer cells and fibroblast-like, tumor- infiltrating, and endothelial cells | n.e. | Correlation with high grade tumors and with stage (T1) | COX-1 | MVD, LVD, PI, increased risk of metastasis | No association with age and gender | No association with recurrence | [177] |
| Prostate cancer | HO-1 positive macrophages infiltrate and in bone metastasis | n.e. | High-grade tumors Gleason score 7–10 | n.e. | Bone metastasis | n.e. | n.e. | [117] |
| Negative correlation among HO-1 expression and tumor aggressiveness/poor prognosis | ||||||||
| Colorectal cancer | High level | n.e. | Invasive CRC | Significant correlation with K-ras | n.e. | Significant correlation with normal CEA level | Better prognosis, increased MTS | [181] |
| High level | n.e. | n.e. | n.e. | Low vascular invasion and lymph node metastasis | n.e. | Better survival rate | [180] | |
| Gastric cancer | High level | n.e. | Well and moderate differentiated | n.e. | Negative lymph node metastasis | n.e. | Better prognosis | [182] |
| Oral squamous cell carcinoma | High level | n.e. | Well-differentiated Grade G1 No association with T stage | n.e. | Low lymph node metastasis | No association with age and sex No association with clinical stage | n.e. | [184] |
| Small intestinal adenocarcinoma | High level | n.e. | Low T stage (T1, T2, T3) | n.e. | Low pancreatic invasion | n.e. | Tend to have longer OS (difference not significative) | [183] |
| Different correlation among HO-1 expression and tumor aggressiveness/poor prognosis depending on HO-1 subcellular localization | ||||||||
| Breast cancer | High level in malignant epithelial cells | n.e. | Grade I-II (>80%) | Positive correlation with E-cadherin | Negative correlation with lymph node metastasis | Reduced tumor size | Longer OS with increased MST | [186] |
| Colorectal cancer | High level in cancer cells and in stromal cells (fibroblasts, neutrophils, and macrophages) | n.e. | Well-differentiated adenocarcinoma Nuclear HO-1 localization in moderate and poor differentiated No association with TNM | n.e. | No correlation with lymph node and liver metastasis | n.e. | n.e. | [187] |
| Head and neck squamous cell carcinoma | High level | n.e. | High rate of HO-1 positivity in well-differentiated and moderately differentiated (<90%) Poor-differentiated high rate of nuclear HO-1 | n.e. | n.e. | No association with age, gender, tumor location | n.e. | [185] |
6. HO-1 and Tumor Therapies
6.1. Inhibition of HO-1 by Pharmacological Compounds
6.2. Inhibition of HO-1 by RNA Interference and CRIPR/Cas9 Technology
| Pharmacological Inhibitors | Benefits | Drawbacks | Ref. | |
|---|---|---|---|---|
| Porphyrin-Based Compounds | ||||
Metalloporphyrins
|
|
| [196,201,202] | |
Modified protoporphyrins
|
| [206,207,208,209,210] | ||
| Imidazole-based compounds | ||||
|
|
| [211,212] | |
| Genetic tools | ||||
| Small interfering RNA and short hairpin RNA |
|
| [195] | |
| CRISPR/Cas9 |
|
| [195] |
6.3. Induction of HO-1 as a Therapeutic Strategy
7. Future Perspectives and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALL | acute lymphoblastic leukemia |
| AML | acute myeloid leukemia |
| BR | bilirubin |
| BVRA | biliverdin reductase A |
| ccRCC | clear cell Renal cell carcinoma |
| CML | chronic myeloid leukemia |
| DC | dendritic cells |
| EV | extracellular vescicles |
| HO-1 | heme oxygenase 1 |
| MAPK | mitogen-activated protein kinase pathway |
| NK | natural killer cells |
| NMIBC | non-muscle-invasive bladder cancer |
| NOS | nitric oxide synthase |
| PMBC | peripheral blood mononuclear cells |
| ROS | reactive oxygen species |
| sGC | soluble guanylyl cyclase |
| SnMPPIX | tin mesoporphyrin IX |
| SnPPIX | tin protoporphyrin IX |
| SSP | signal peptide peptidase |
| VSMC | vascular smooth muscle cells |
| ZnPPIX | zinc(II) protoporphyrin IX |
References
- Maines, M.D. Heme Oxygenase: Function, Multiplicity, Regulatory Mechanisms, and Clinical Applications. FASEB J. 1988, 2, 2557–2568. [Google Scholar] [CrossRef] [PubMed]
- Waza, A.A.; Hamid, Z.; Ali, S.; Bhat, S.A.; Bhat, M.A. A Review on Heme Oxygenase-1 Induction: Is It a Necessary Evil. Inflamm. Res. 2018, 67, 579–588. [Google Scholar] [CrossRef]
- Kutty, R.K.; Nagineni, C.N.; Kutty, G.; Hooks, J.J.; Chader, G.J.; Wiggert, B. Increased Expression of Heme Oxygenase-1 in Human Retinal Pigment Epithelial Cells by Transforming Growth Factor-Beta. J. Cell Physiol. 1994, 159, 371–378. [Google Scholar] [CrossRef]
- Bian, C.; Zhong, M.; Nisar, M.F.; Wu, Y.; Ouyang, M.; Bartsch, J.W.; Zhong, J.L. A Novel Heme Oxygenase-1 Splice Variant, 14kDa HO-1, Promotes Cell Proliferation and Increases Relative Telomere Length. Biochem. Biophys Res. Commun. 2018, 500, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Furfaro, A.L.; Traverso, N.; Domenicotti, C.; Piras, S.; Moretta, L.; Marinari, U.M.; Pronzato, M.A.; Nitti, M. The Nrf2/HO-1 Axis in Cancer Cell Growth and Chemoresistance. Oxid. Med. Cell. Longev. 2016, 2016, 1958174. [Google Scholar] [CrossRef]
- Keyse, S.M.; Tyrrell, R.M. Heme Oxygenase Is the Major 32-KDa Stress Protein Induced in Human Skin Fibroblasts by UVA Radiation, Hydrogen Peroxide, and Sodium Arsenite. Proc. Natl. Acad. Sci. USA 1989, 86, 99–103. [Google Scholar] [CrossRef]
- Alam, J.; Shibahara, S.; Smith, A. Transcriptional Activation of the Heme Oxygenase Gene by Heme and Cadmium in Mouse Hepatoma Cells. J. Biol. Chem. 1989, 264, 6371–6375. [Google Scholar] [CrossRef]
- Foresti, R.; Clark, J.E.; Green, C.J.; Motterlini, R. Thiol Compounds Interact with Nitric Oxide in Regulating Heme Oxygenase-1 Induction in Endothelial Cells. Involvement of Superoxide and Peroxynitrite Anions. J. Biol. Chem. 1997, 272, 18411–18417. [Google Scholar] [CrossRef]
- Ayer, A.; Zarjou, A.; Agarwal, A.; Stocker, R. Heme Oxygenases in Cardiovascular Health and Disease. Physiol. Rev. 2016, 96, 1449–1508. [Google Scholar] [CrossRef]
- Nitti, M.; Piras, S.; Marinari, U.M.; Moretta, L.; Pronzato, M.A.; Furfaro, A.L. HO-1 Induction in Cancer Progression: A Matter of Cell Adaptation. Antioxidants 2017, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Jozkowicz, A.; Dulak, J. HO-1/CO System in Tumor Growth, Angiogenesis and Metabolism-Targeting HO-1 as an Anti-Tumor Therapy. Vasc. Pharmacol. 2015, 74, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, S.A.; Carreño, L.J.; Espinoza, J.A.; Mackern-Oberti, J.P.; Alvarez-Lobos, M.M.; Riedel, C.A.; Bueno, S.M.; Kalergis, A.M. Modulation of Antigen Processing by Haem-Oxygenase 1. Implications on Inflammation and Tolerance. Immunology 2016, 149, 1–12. [Google Scholar] [CrossRef]
- Vijayan, V.; Wagener, F.A.D.T.G.; Immenschuh, S. The Macrophage Heme-Heme Oxygenase-1 System and Its Role in Inflammation. Biochem. Pharmacol. 2018, 153, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Siow, R.C.; Sato, H.; Mann, G.E. Heme Oxygenase-Carbon Monoxide Signalling Pathway in Atherosclerosis: Anti-Atherogenic Actions of Bilirubin and Carbon Monoxide? Cardiovasc. Res. 1999, 41, 385–394. [Google Scholar] [CrossRef]
- Otterbein, L.E.; Foresti, R.; Motterlini, R. Heme Oxygenase-1 and Carbon Monoxide in the Heart: The Balancing Act Between Danger Signaling and Pro-Survival. Circ. Res. 2016, 118, 1940–1959. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Kondo, K.; Momiyama, Y. The Protective Role of Heme Oxygenase-1 in Atherosclerotic Diseases. Int. J. Mol. Sci. 2019, 20, 3628. [Google Scholar] [CrossRef]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin Is an Antioxidant of Possible Physiological Importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R. Antioxidant Activities of Bile Pigments. Antioxid. Redox Signal. 2004, 6, 841–849. [Google Scholar] [CrossRef]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The Enzymatic Conversion of Heme to Bilirubin by Microsomal Heme Oxygenase. Proc. Natl. Acad. Sci. USA 1968, 61, 748–755. [Google Scholar] [CrossRef]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The Enzymatic Conversion of Hemoglobin to Bilirubin. Trans. Assoc. Am. Physicians 1969, 82, 363–371. [Google Scholar]
- Neuzil, J.; Stocker, R. Bilirubin Attenuates Radical-Mediated Damage to Serum Albumin. FEBS Lett. 1993, 331, 281–284. [Google Scholar] [CrossRef]
- Sedlak, T.W.; Saleh, M.; Higginson, D.S.; Paul, B.D.; Juluri, K.R.; Snyder, S.H. Bilirubin and Glutathione Have Complementary Antioxidant and Cytoprotective Roles. Proc. Natl. Acad. Sci. USA 2009, 106, 5171–5176. [Google Scholar] [CrossRef]
- Wu, T.W.; Fung, K.P.; Yang, C.C. Unconjugated Bilirubin Inhibits the Oxidation of Human Low Density Lipoprotein Better than Trolox. Life Sci. 1994, 54, P477–P481. [Google Scholar] [CrossRef]
- He, M.; Nitti, M.; Piras, S.; Furfaro, A.L.; Traverso, N.; Pronzato, M.A.; Mann, G.E. Heme Oxygenase-1-Derived Bilirubin Protects Endothelial Cells against High Glucose-Induced Damage. Free Radic. Biol. Med. 2015, 89, 91–98. [Google Scholar] [CrossRef]
- Wu, B.; Wu, Y.; Tang, W. Heme Catabolic Pathway in Inflammation and Immune Disorders. Front. Pharmacol. 2019, 10, 825. [Google Scholar] [CrossRef] [PubMed]
- Canesin, G.; Hejazi, S.M.; Swanson, K.D.; Wegiel, B. Heme-Derived Metabolic Signals Dictate Immune Responses. Front. Immunol. 2020, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W. Heme Oxygenase-1/Carbon Monoxide as Modulators of Autophagy and Inflammation. Arch. Biochem. Biophys. 2019, 678, 108186. [Google Scholar] [CrossRef]
- Keshavan, P.; Deem, T.L.; Schwemberger, S.J.; Babcock, G.F.; Cook-Mills, J.M.; Zucker, S.D. Unconjugated Bilirubin Inhibits VCAM-1-Mediated Transendothelial Leukocyte Migration. J. Immunol. 2005, 174, 3709–3718. [Google Scholar] [CrossRef] [PubMed]
- Grochot-Przeczek, A.; Dulak, J.; Jozkowicz, A. Haem Oxygenase-1: Non-Canonical Roles in Physiology and Pathology. Clin. Sci. 2012, 122, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, G.L.; Rigato, I.; Ostrow, J.D.; Bossi, F.; Bortoluzzi, A.; Sukowati, C.H.C.; Tedesco, F.; Tiribelli, C. Bilirubin Inhibits the TNFalpha-Related Induction of Three Endothelial Adhesion Molecules. Biochem. Biophys. Res. Commun. 2009, 386, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Nitti, M.; Furfaro, A.L.; Mann, G.E. Heme Oxygenase Dependent Bilirubin Generation in Vascular Cells: A Role in Preventing Endothelial Dysfunction in Local Tissue Microenvironment? Front. Physiol. 2020, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Petrache, I.; Otterbein, L.E.; Alam, J.; Wiegand, G.W.; Choi, A.M. Heme Oxygenase-1 Inhibits TNF-Alpha-Induced Apoptosis in Cultured Fibroblasts. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 278, L312–L319. [Google Scholar] [CrossRef]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme Oxygenase-1/Carbon Monoxide: From Basic Science to Therapeutic Applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Ma, K.C.; Choi, A.M.K. Carbon Monoxide in Lung Cell Physiology and Disease. Am. J. Physiol. Cell Physiol. 2018, 314, C211–C227. [Google Scholar] [CrossRef]
- Motterlini, R.; Foresti, R.; Bassi, R.; Green, C.J. Curcumin, an Antioxidant and Anti-Inflammatory Agent, Induces Heme Oxygenase-1 and Protects Endothelial Cells against Oxidative Stress. Free Radic. Biol. Med. 2000, 28, 1303–1312. [Google Scholar] [CrossRef]
- Dennery, P.A. Heme Oxygenase in Neonatal Lung Injury and Repair. Antioxid. Redox Signal. 2014, 21, 1881–1892. [Google Scholar] [CrossRef] [PubMed]
- Dulak, J.; Jozkowicz, A.; Foresti, R.; Kasza, A.; Frick, M.; Huk, I.; Green, C.J.; Pachinger, O.; Weidinger, F.; Motterlini, R. Heme Oxygenase Activity Modulates Vascular Endothelial Growth Factor Synthesis in Vascular Smooth Muscle Cells. Antioxid. Redox Signal. 2002, 4, 229–240. [Google Scholar] [CrossRef]
- Jozkowicz, A.; Huk, I.; Nigisch, A.; Weigel, G.; Dietrich, W.; Motterlini, R.; Dulak, J. Heme Oxygenase and Angiogenic Activity of Endothelial Cells: Stimulation by Carbon Monoxide and Inhibition by Tin Protoporphyrin-IX. Antioxid. Redox Signal. 2003, 5, 155–162. [Google Scholar] [CrossRef]
- Loboda, A.; Jazwa, A.; Grochot-Przeczek, A.; Rutkowski, A.J.; Cisowski, J.; Agarwal, A.; Jozkowicz, A.; Dulak, J. Heme Oxygenase-1 and the Vascular Bed: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox Signal. 2008, 10, 1767–1812. [Google Scholar] [CrossRef]
- Rochette, L.; Cottin, Y.; Zeller, M.; Vergely, C. Carbon Monoxide: Mechanisms of Action and Potential Clinical Implications. Pharmacol. Ther. 2013, 137, 133–152. [Google Scholar] [CrossRef]
- Ryter, S.W.; Choi, A.M.K. Targeting Heme Oxygenase-1 and Carbon Monoxide for Therapeutic Modulation of Inflammation. Transl. Res. 2016, 167, 7–34. [Google Scholar] [CrossRef]
- Pae, H.O.; Choi, B.M.; Oh, G.S.; Lee, M.S.; Ryu, D.G.; Rhew, H.Y.; Kim, Y.M.; Chung, H.T. Roles of Heme Oxygenase-1 in the Antiproliferative and Antiapoptotic Effects of Nitric Oxide on Jurkat T Cells. Mol. Pharmacol. 2004, 66, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.E.; Bach, F.H.; Alam, J.; Soares, M.; Tao Lu, H.; Wysk, M.; Davis, R.J.; Flavell, R.A.; Choi, A.M. Carbon Monoxide Has Anti-Inflammatory Effects Involving the Mitogen-Activated Protein Kinase Pathway. Nat. Med. 2000, 6, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Balla, G.; Jacob, H.S.; Balla, J.; Rosenberg, M.; Nath, K.; Apple, F.; Eaton, J.W.; Vercellotti, G.M. Ferritin: A Cytoprotective Antioxidant Strategem of Endothelium. J. Biol. Chem. 1992, 267, 18148–18153. [Google Scholar] [CrossRef]
- Baker, H.M.; Anderson, B.F.; Baker, E.N. Dealing with Iron: Common Structural Principles in Proteins That Transport Iron and Heme. Proc. Natl. Acad. Sci. USA 2003, 100, 3579–3583. [Google Scholar] [CrossRef]
- Chiang, S.-K.; Chen, S.-E.; Chang, L.-C. A Dual Role of Heme Oxygenase-1 in Cancer Cells. Int. J. Mol. Sci. 2018, 20, 39. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Long, D. Nrf2 and Ferroptosis: A New Research Direction for Neurodegenerative Diseases. Front. Neurosci. 2020, 14, 267. [Google Scholar] [CrossRef]
- Di Biase, S.; Longo, V.D. Fasting-Induced Differential Stress Sensitization in Cancer Treatment. Mol. Cell. Oncol. 2016, 3, e1117701. [Google Scholar] [CrossRef]
- Lavrovsky, Y.; Schwartzman, M.L.; Levere, R.D.; Kappas, A.; Abraham, N.G. Identification of Binding Sites for Transcription Factors NF-Kappa B and AP-2 in the Promoter Region of the Human Heme Oxygenase 1 Gene. Proc. Natl. Acad. Sci. USA 1994, 91, 5987–5991. [Google Scholar] [CrossRef] [PubMed]
- Lavrovsky, Y.; Schwartzman, M.L.; Abraham, N.G. Novel Regulatory Sites of the Human Heme Oxygenase-1 Promoter Region. Biochem. Biophys. Res. Commun. 1993, 196, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.V.; Sapochnik, D.; Garcia Solá, M.; Coso, O. Regulation of the Expression of Heme Oxygenase-1: Signal Transduction, Gene Promoter Activation, and Beyond. Antioxid. Redox Signal. 2020, 32, 1033–1044. [Google Scholar] [CrossRef]
- Was, H.; Dulak, J.; Jozkowicz, A. Heme Oxygenase-1 in Tumor Biology and Therapy. Curr. Drug Targets 2010, 11, 1551–1570. [Google Scholar] [CrossRef]
- Exner, M.; Minar, E.; Wagner, O.; Schillinger, M. The Role of Heme Oxygenase-1 Promoter Polymorphisms in Human Disease. Free Radic. Biol. Med. 2004, 37, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-M.; Zheng, Y.-Y.; Gao, Y.; Zhang, J.-Z.; Liu, F.; Yang, Y.-N.; Li, X.-M.; Ma, Y.-T.; Xie, X. Heme Oxygenase-1 Gene Promoter Polymorphisms Are Associated with Coronary Heart Disease and Restenosis after Percutaneous Coronary Intervention: A Meta-Analysis. Oncotarget 2016, 7, 83437–83450. [Google Scholar] [CrossRef]
- Daenen, K.E.L.; Martens, P.; Bammens, B. Association of HO-1 (GT)n Promoter Polymorphism and Cardiovascular Disease: A Reanalysis of the Literature. Can. J. Cardiol. 2016, 32, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, F.-F.; Huang, Y.-B.; Zheng, H.; Song, F.-J.; Chen, K.-X. Association between the (GT)n Polymorphism of the HO-1 Gene Promoter Region and Cancer Risk: A Meta-Analysis. Asian Pac. J. Cancer Prev. 2014, 15, 4617–4622. [Google Scholar] [CrossRef] [PubMed]
- Horio, T.; Morishita, E.; Mizuno, S.; Uchino, K.; Hanamura, I.; Espinoza, J.L.; Morishima, Y.; Kodera, Y.; Onizuka, M.; Kashiwase, K.; et al. Donor Heme Oxygenase-1 Promoter Gene Polymorphism Predicts Survival after Unrelated Bone Marrow Transplantation for High-Risk Patients. Cancers 2020, 12, 424. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Goto, Y.; Takagi, S.; Baba, S.; Tago, N.; Nonogi, H.; Iwai, N. A Promoter Variant of the Heme Oxygenase-1 Gene May Reduce the Incidence of Ischemic Heart Disease in Japanese. Atherosclerosis 2004, 173, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Buis, C.I.; van der Steege, G.; Visser, D.S.; Nolte, I.M.; Hepkema, B.G.; Nijsten, M.; Slooff, M.J.H.; Porte, R.J. Heme Oxygenase-1 Genotype of the Donor Is Associated with Graft Survival after Liver Transplantation. Am. J. Transplant. 2008, 8, 377–385. [Google Scholar] [CrossRef]
- Bukowska-Strakova, K.; Włodek, J.; Pitera, E.; Kozakowska, M.; Konturek-Cieśla, A.; Cieśla, M.; Gońka, M.; Nowak, W.; Wieczorek, A.; Pawińska-Wąsikowska, K.; et al. Role of HMOX1 Promoter Genetic Variants in Chemoresistance and Chemotherapy Induced Neutropenia in Children with Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2021, 22, 988. [Google Scholar] [CrossRef]
- Paladino, S.; Conte, A.; Caggiano, R.; Pierantoni, G.M.; Faraonio, R. Nrf2 Pathway in Age-Related Neurological Disorders: Insights into MicroRNAs. Cell Physiol. Biochem. 2018, 47, 1951–1976. [Google Scholar] [CrossRef]
- Mitsuishi, Y.; Motohashi, H.; Yamamoto, M. The Keap1-Nrf2 System in Cancers: Stress Response and Anabolic Metabolism. Front. Oncol. 2012, 2, 200. [Google Scholar] [CrossRef]
- Shibata, T.; Kokubu, A.; Gotoh, M.; Ojima, H.; Ohta, T.; Yamamoto, M.; Hirohashi, S. Genetic Alteration of Keap1 Confers Constitutive Nrf2 Activation and Resistance to Chemotherapy in Gallbladder Cancer. Gastroenterology 2008, 135, 1358–1368.e1-4. [Google Scholar] [CrossRef]
- Na, H.K.; Surh, Y.J. Oncogenic Potential of Nrf2 and Its Principal Target Protein Heme Oxygenase-1. Free Radic. Biol. Med. 2014, 67, 353–365. [Google Scholar] [CrossRef]
- Muscarella, L.A.; Parrella, P.; D’Alessandro, V.; la Torre, A.; Barbano, R.; Fontana, A.; Tancredi, A.; Guarnieri, V.; Balsamo, T.; Coco, M.; et al. Frequent Epigenetics Inactivation of KEAP1 Gene in Non-Small Cell Lung Cancer. Epigenetics 2011, 6, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Hanada, N.; Takahata, T.; Zhou, Q.; Ye, X.; Sun, R.; Itoh, J.; Ishiguro, A.; Kijima, H.; Mimura, J.; Itoh, K.; et al. Methylation of the KEAP1 Gene Promoter Region in Human Colorectal Cancer. BMC Cancer 2012, 12, 66. [Google Scholar] [CrossRef]
- Zhao, X.-Q.; Zhang, Y.-F.; Xia, Y.-F.; Zhou, Z.-M.; Cao, Y.-Q. Promoter Demethylation of Nuclear Factor-Erythroid 2-Related Factor 2 Gene in Drug-Resistant Colon Cancer Cells. Oncol. Lett. 2015, 10, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- van der Wijst, M.G.; Brown, R.; Rots, M.G. Nrf2, the Master Redox Switch: The Achilles’ Heel of Ovarian Cancer? Biochim. Biophys. Acta 2014, 1846, 494–509. [Google Scholar] [CrossRef]
- Ogawa, K.; Sun, J.; Taketani, S.; Nakajima, O.; Nishitani, C.; Sassa, S.; Hayashi, N.; Yamamoto, M.; Shibahara, S.; Fujita, H.; et al. Heme Mediates Derepression of Maf Recognition Element through Direct Binding to Transcription Repressor Bach1. EMBO J. 2001, 20, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Davudian, S.; Mansoori, B.; Shajari, N.; Mohammadi, A.; Baradaran, B. BACH1, the Master Regulator Gene: A Novel Candidate Target for Cancer Therapy. Gene 2016, 588, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Lignitto, L.; LeBoeuf, S.E.; Homer, H.; Jiang, S.; Askenazi, M.; Karakousi, T.R.; Pass, H.I.; Bhutkar, A.J.; Tsirigos, A.; Ueberheide, B.; et al. Nrf2 Activation Promotes Lung Cancer Metastasis by Inhibiting the Degradation of Bach1. Cell 2019, 178, 316–329.e18. [Google Scholar] [CrossRef]
- Wiel, C.; Le Gal, K.; Ibrahim, M.X.; Jahangir, C.A.; Kashif, M.; Yao, H.; Ziegler, D.V.; Xu, X.; Ghosh, T.; Mondal, T.; et al. BACH1 Stabilization by Antioxidants Stimulates Lung Cancer Metastasis. Cell 2019, 178, 330–345.e22. [Google Scholar] [CrossRef]
- Alam, J.; Wicks, C.; Stewart, D.; Gong, P.; Touchard, C.; Otterbein, S.; Choi, A.M.; Burow, M.E.; Tou, J. Mechanism of Heme Oxygenase-1 Gene Activation by Cadmium in MCF-7 Mammary Epithelial Cells. Role of P38 Kinase and Nrf2 Transcription Factor. J. Biol. Chem. 2000, 275, 27694–27702. [Google Scholar] [CrossRef]
- Liu, Z.-M.; Chen, G.G.; Ng, E.K.W.; Leung, W.-K.; Sung, J.J.Y.; Chung, S.C.S. Upregulation of Heme Oxygenase-1 and P21 Confers Resistance to Apoptosis in Human Gastric Cancer Cells. Oncogene 2004, 23, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Dal-Cim, T.; Molz, S.; Egea, J.; Parada, E.; Romero, A.; Budni, J.; Martín de Saavedra, M.D.; del Barrio, L.; Tasca, C.I.; López, M.G. Guanosine Protects Human Neuroblastoma SH-SY5Y Cells against Mitochondrial Oxidative Stress by Inducing Heme Oxigenase-1 via PI3K/Akt/GSK-3β Pathway. Neurochem. Int. 2012, 61, 397–404. [Google Scholar] [CrossRef]
- Talabnin, C.; Talabnin, K.; Wongkham, S. Enhancement of Piperlongumine Chemosensitivity by Silencing Heme Oxygenase-1 Expression in Cholangiocarcinoma Cell Lines. Oncol. Lett. 2020, 20, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ku, C.H.; Siow, R.C. Regulation of the Nrf2 Antioxidant Pathway by MicroRNAs: New Players in Micromanaging Redox Homeostasis. Free Radic. Biol. Med. 2013, 64, 4–11. [Google Scholar] [CrossRef]
- Gu, S.; Lai, Y.; Chen, H.; Liu, Y.; Zhang, Z. MiR-155 Mediates Arsenic Trioxide Resistance by Activating Nrf2 and Suppressing Apoptosis in Lung Cancer Cells. Sci. Rep. 2017, 7, 12155. [Google Scholar] [CrossRef] [PubMed]
- Eades, G.; Yang, M.; Yao, Y.; Zhang, Y.; Zhou, Q. MiR-200a Regulates Nrf2 Activation by Targeting Keap1 MRNA in Breast Cancer Cells. J. Biol. Chem. 2011, 286, 40725–40733. [Google Scholar] [CrossRef] [PubMed]
- Pu, M.; Li, C.; Qi, X.; Chen, J.; Wang, Y.; Gao, L.; Miao, L.; Ren, J. MiR-1254 Suppresses HO-1 Expression through Seed Region-Dependent Silencing and Non-Seed Interaction with TFAP2A Transcript to Attenuate NSCLC Growth. PLoS Genet. 2017, 13, e1006896. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, J.-S.; Wen, J.-K.; Gao, H.-T.; Zheng, B.; Qu, C.-B.; Liu, K.-L.; Zhang, M.-L.; Gu, J.-F.; Li, J.-D.; et al. Silencing of MiR-193a-5p Increases the Chemosensitivity of Prostate Cancer Cells to Docetaxel. J. Exp. Clin. Cancer Res. 2017, 36, 178. [Google Scholar] [CrossRef]
- Piras, S.; Furfaro, A.L.; Caggiano, R.; Brondolo, L.; Garibaldi, S.; Ivaldo, C.; Marinari, U.M.; Pronzato, M.A.; Faraonio, R.; Nitti, M. MicroRNA-494 Favors HO-1 Expression in Neuroblastoma Cells Exposed to Oxidative Stress in a Bach1-Independent Way. Front. Oncol. 2018, 8, 199. [Google Scholar] [CrossRef]
- Durante, W. Targeting Heme Oxygenase-1 in the Arterial Response to Injury and Disease. Antioxidants 2020, 9, 829. [Google Scholar] [CrossRef]
- Huber, W.J.; Backes, W.L. Expression and Characterization of Full-Length Human Heme Oxygenase-1: The Presence of Intact Membrane-Binding Region Leads to Increased Binding Affinity for NADPH Cytochrome P450 Reductase. Biochemistry 2007, 46, 12212–12219. [Google Scholar] [CrossRef]
- Jung, N.-H.; Kim, H.P.; Kim, B.-R.; Cha, S.H.; Kim, G.A.; Ha, H.; Na, Y.E.; Cha, Y.-N. Evidence for Heme Oxygenase-1 Association with Caveolin-1 and -2 in Mouse Mesangial Cells. IUBMB Life 2003, 55, 525–532. [Google Scholar] [CrossRef]
- Slebos, D.J.; Ryter, S.W.; van der Toorn, M.; Liu, F.; Guo, F.; Baty, C.J.; Karlsson, J.M.; Watkins, S.C.; Kim, H.P.; Wang, X.; et al. Mitochondrial Localization and Function of Heme Oxygenase-1 in Cigarette Smoke-Induced Cell Death. Am. J. Respir. Cell Mol. Biol. 2007, 36, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Weis, S.; Yang, G.; Weng, Y.H.; Helston, R.; Rish, K.; Smith, A.; Bordner, J.; Polte, T.; Gaunitz, F.; et al. Heme Oxygenase-1 Protein Localizes to the Nucleus and Activates Transcription Factors Important in Oxidative Stress. J. Biol. Chem. 2007, 282, 20621–20633. [Google Scholar] [CrossRef]
- Lin, Q.S.; Weis, S.; Yang, G.; Zhuang, T.; Abate, A.; Dennery, P.A. Catalytic Inactive Heme Oxygenase-1 Protein Regulates Its Own Expression in Oxidative Stress. Free Radic. Biol. Med. 2008, 44, 847–855. [Google Scholar] [CrossRef]
- Hsu, F.F.; Yeh, C.T.; Sun, Y.J.; Chiang, M.T.; Lan, W.M.; Li, F.A.; Lee, W.H.; Chau, L.Y. Signal Peptide Peptidase-Mediated Nuclear Localization of Heme Oxygenase-1 Promotes Cancer Cell Proliferation and Invasion Independent of Its Enzymatic Activity. Oncogene 2015, 34, 2410–2411. [Google Scholar] [CrossRef]
- Biswas, C.; Shah, N.; Muthu, M.; La, P.; Fernando, A.P.; Sengupta, S.; Yang, G.; Dennery, P.A. Nuclear Heme Oxygenase-1 (HO-1) Modulates Subcellular Distribution and Activation of Nrf2, Impacting Metabolic and Anti-Oxidant Defenses. J. Biol. Chem. 2014, 289, 26882–26894. [Google Scholar] [CrossRef]
- Hsu, F.-F.; Chiang, M.-T.; Li, F.-A.; Yeh, C.-T.; Lee, W.-H.; Chau, L.-Y. Acetylation Is Essential for Nuclear Heme Oxygenase-1-Enhanced Tumor Growth and Invasiveness. Oncogene 2017, 36, 6805–6814. [Google Scholar] [CrossRef]
- Mascaró, M.; Alonso, E.N.; Alonso, E.G.; Lacunza, E.; Curino, A.C.; Facchinetti, M.M. Nuclear Localization of Heme Oxygenase-1 in Pathophysiological Conditions: Does It Explain the Dual Role in Cancer? Antioxidants 2021, 10, 87. [Google Scholar] [CrossRef]
- Dennery, P.A. Signaling Function of Heme Oxygenase Proteins. Antioxid. Redox Signal. 2014, 20, 1743–1753. [Google Scholar] [CrossRef]
- Tibullo, D.; Barbagallo, I.; Giallongo, C.; La Cava, P.; Parrinello, N.; Vanella, L.; Stagno, F.; Palumbo, G.A.; Li Volti, G.; Di Raimondo, F. Nuclear Translocation of Heme Oxygenase-1 Confers Resistance to Imatinib in Chronic Myeloid Leukemia Cells. Curr. Pharm. Des. 2013, 19, 2765–2770. [Google Scholar] [CrossRef]
- Elguero, B.; Gueron, G.; Giudice, J.; Toscani, M.A.; De Luca, P.; Zalazar, F.; Coluccio-Leskow, F.; Meiss, R.; Navone, N.; De Siervi, A.; et al. Unveiling the Association of STAT3 and HO-1 in Prostate Cancer: Role beyond Heme Degradation. Neoplasia 2012, 14, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Gueron, G.; De Siervi, A.; Ferrando, M.; Salierno, M.; De Luca, P.; Elguero, B.; Meiss, R.; Navone, N.; Vazquez, E.S. Critical Role of Endogenous Heme Oxygenase 1 as a Tuner of the Invasive Potential of Prostate Cancer Cells. Mol. Cancer Res. MCR 2009, 7, 1745–1755. [Google Scholar] [CrossRef]
- Ferrando, M.; Gueron, G.; Elguero, B.; Giudice, J.; Salles, A.; Leskow, F.C.; Jares-Erijman, E.A.; Colombo, L.; Meiss, R.; Navone, N.; et al. Heme Oxygenase 1 (HO-1) Challenges the Angiogenic Switch in Prostate Cancer. Angiogenesis 2011, 14, 467–479. [Google Scholar] [CrossRef]
- Degese, M.S.; Mendizabal, J.E.; Gandini, N.A.; Gutkind, J.S.; Molinolo, A.; Hewitt, S.M.; Curino, A.C.; Coso, O.A.; Facchinetti, M.M. Expression of Heme Oxygenase-1 in Non-Small Cell Lung Cancer (NSCLC) and Its Correlation with Clinical Data. Lung Cancer 2012, 77, 168–175. [Google Scholar] [CrossRef]
- Vanella, L.; Barbagallo, I.; Tibullo, D.; Forte, S.; Zappalà, A.; Li Volti, G. The Non-Canonical Functions of the Heme Oxygenases. Oncotarget 2016, 7, 69075–69086. [Google Scholar] [CrossRef]
- Signorelli, S.S.; Li Volsi, G.; Fiore, V.; Mangiafico, M.; Barbagallo, I.; Parenti, R.; Rizzo, M.; Li Volti, G. Plasma Heme Oxygenase-1 Is Decreased in Peripheral Artery Disease Patients. Mol. Med. Rep. 2016, 14, 3459–3463. [Google Scholar] [CrossRef] [PubMed]
- Serpero, L.D.; Frigiola, A.; Gazzolo, D. Human Milk and Formulae: Neurotrophic and New Biological Factors. Early Hum. Dev. 2012, 88 (Suppl. 1), S9–S12. [Google Scholar] [CrossRef]
- Novo, G.; Cappello, F.; Rizzo, M.; Fazio, G.; Zambuto, S.; Tortorici, E.; Marino Gammazza, A.; Gammazza, A.M.; Corrao, S.; Zummo, G.; et al. Hsp60 and Heme Oxygenase-1 (Hsp32) in Acute Myocardial Infarction. Transl. Res. 2011, 157, 285–292. [Google Scholar] [CrossRef]
- Zager, R.A.; Johnson, A.C.M.; Becker, K. Plasma and Urinary Heme Oxygenase-1 in AKI. J. Am. Soc. Nephrol. 2012, 23, 1048–1057. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Margolis, L.; Sadovsky, Y. The Biology of Extracellular Vesicles: The Known Unknowns. PLoS Biol. 2019, 17, e3000363. [Google Scholar] [CrossRef]
- El-Rifaie, A.-A.A.; Sabry, D.; Doss, R.W.; Kamal, M.A.; Abd El Hassib, D.M. Heme Oxygenase and Iron Status in Exosomes of Psoriasis Patients. Arch. Dermatol. Res. 2018, 310, 651–656. [Google Scholar] [CrossRef]
- Cressatti, M.; Galindez, J.M.; Juwara, L.; Orlovetskie, N.; Velly, A.M.; Eintracht, S.; Liberman, A.; Gornitsky, M.; Schipper, H.M. Characterization and Heme Oxygenase-1 Content of Extracellular Vesicles in Human Biofluids. J. Neurochem. 2020. [Google Scholar] [CrossRef]
- Hurwitz, S.N.; Rider, M.A.; Bundy, J.L.; Liu, X.; Singh, R.K.; Meckes, D.G. Proteomic Profiling of NCI-60 Extracellular Vesicles Uncovers Common Protein Cargo and Cancer Type-Specific Biomarkers. Oncotarget 2016, 7, 86999–87015. [Google Scholar] [CrossRef]
- Goodman, A.I.; Choudhury, M.; da Silva, J.L.; Schwartzman, M.L.; Abraham, N.G. Overexpression of the Heme Oxygenase Gene in Renal Cell Carcinoma. Proc. Soc. Exp. Biol. Med. 1997, 214, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Was, H.; Cichon, T.; Smolarczyk, R.; Rudnicka, D.; Stopa, M.; Chevalier, C.; Leger, J.J.; Lackowska, B.; Grochot, A.; Bojkowska, K.; et al. Overexpression of Heme Oxygenase-1 in Murine Melanoma: Increased Proliferation and Viability of Tumor Cells, Decreased Survival of Mice. Am. J. Pathol. 2006, 169, 2181–2198. [Google Scholar] [CrossRef]
- Maines, M.D.; Abrahamsson, P.A. Expression of Heme Oxygenase-1 (HSP32) in Human Prostate: Normal, Hyperplastic, and Tumor Tissue Distribution. Urology 1996, 47, 727–733. [Google Scholar] [CrossRef]
- Schacter, B.A.; Kurz, P. Alterations in Microsomal Drug Metabolism and Heme Oxygenase Activity in Isolated Hepatic Parenchymal and Sinusoidal Cells in Murphy-Sturm Lymphosarcoma-Bearing Rats. Clin. Investig. Med. Med. Clin. Exp. 1986, 9, 150–155. [Google Scholar]
- Sferrazzo, G.; Di Rosa, M.; Barone, E.; Li Volti, G.; Musso, N.; Tibullo, D.; Barbagallo, I. Heme Oxygenase-1 in Central Nervous System Malignancies. J. Clin. Med. 2020, 9, 1562. [Google Scholar] [CrossRef]
- Li Volti, G.; Tibullo, D.; Vanella, L.; Giallongo, C.; Di Raimondo, F.; Forte, S.; Di Rosa, M.; Signorelli, S.S.; Barbagallo, I. The Heme Oxygenase System in Hematological Malignancies. Antioxid. Redox Signal. 2017, 27, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Sunamura, M.; Duda, D.G.; Ghattas, M.H.; Lozonschi, L.; Motoi, F.; Yamauchi, J.; Matsuno, S.; Shibahara, S.; Abraham, N.G. Heme Oxygenase-1 Accelerates Tumor Angiogenesis of Human Pancreatic Cancer. Angiogenesis 2003, 6, 15–24. [Google Scholar] [CrossRef]
- Lee, S.S.; Yang, S.F.; Tsai, C.H.; Chou, M.C.; Chou, M.Y.; Chang, Y.C. Upregulation of Heme Oxygenase-1 Expression in Areca-Quid-Chewing-Associated Oral Squamous Cell Carcinoma. J. Formos. Med. Assoc. 2008, 107, 355–363. [Google Scholar] [CrossRef][Green Version]
- Halin Bergström, S.; Nilsson, M.; Adamo, H.; Thysell, E.; Jernberg, E.; Stattin, P.; Widmark, A.; Wikström, P.; Bergh, A. Extratumoral Heme Oxygenase-1 (HO-1) Expressing Macrophages Likely Promote Primary and Metastatic Prostate Tumor Growth. PLoS ONE 2016, 11, e0157280. [Google Scholar] [CrossRef]
- Tsai, J.R.; Wang, H.M.; Liu, P.L.; Chen, Y.H.; Yang, M.C.; Chou, S.H.; Cheng, Y.J.; Yin, W.H.; Hwang, J.J.; Chong, I.W. High Expression of Heme Oxygenase-1 Is Associated with Tumor Invasiveness and Poor Clinical Outcome in Non-Small Cell Lung Cancer Patients. Cell Oncol. 2012, 35, 461–471. [Google Scholar] [CrossRef]
- Wang, X.; Ye, T.; Xue, B.; Yang, M.; Li, R.; Xu, X.; Zeng, X.; Tian, N.; Bao, L.; Huang, Y. Mitochondrial GRIM-19 Deficiency Facilitates Gastric Cancer Metastasis through Oncogenic ROS-NRF2-HO-1 Axis via a NRF2-HO-1 Loop. Gastric. Cancer 2020. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhao, J.; Xue, J.; Zhao, X.; Liu, P. Autophagy Inhibition Promotes Epithelial-Mesenchymal Transition through ROS/HO-1 Pathway in Ovarian Cancer Cells. Am. J. Cancer Res. 2016, 6, 2162–2177. [Google Scholar]
- Chang, Y.J.; Chen, W.Y.; Huang, C.Y.; Liu, H.H.; Wei, P.L. Glucose-Regulated Protein 78 (GRP78) Regulates Colon Cancer Metastasis through EMT Biomarkers and the NRF-2/HO-1 Pathway. Tumour Biol. 2015, 36, 1859–1869. [Google Scholar] [CrossRef]
- Jozkowicz, A.; Huk, I.; Nigisch, A.; Weigel, G.; Weidinger, F.; Dulak, J. Effect of Prostaglandin-J(2) on VEGF Synthesis Depends on the Induction of Heme Oxygenase-1. Antioxid. Redox Signal. 2002, 4, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Fujimoto, K.; Anai, S.; Ohnishi, S.; Kuwada, M.; Nakai, Y.; Inoue, T.; Matsumura, Y.; Tomioka, A.; Ikeda, T.; et al. Heme Oxygenase-1 Promotes Angiogenesis in Urothelial Carcinoma of the Urinary Bladder. Oncol. Rep. 2011, 25, 653–660. [Google Scholar] [CrossRef]
- Kozakowska, M.; Dobrowolska-Glazar, B.; Okoń, K.; Józkowicz, A.; Dobrowolski, Z.; Dulak, J. Preliminary Analysis of the Expression of Selected Proangiogenic and Antioxidant Genes and MicroRNAs in Patients with Non-Muscle-Invasive Bladder Cancer. J. Clin. Med. 2016, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.T.; Hui, L.L.; An, X.S.; Zhang, X.C.; Guo, S.G.; Kui, Z. ZnPPIX Inhibits Peritoneal Metastasis of Gastric Cancer via Its Antiangiogenic Activity. Biomed. Pharmacother. 2015, 71, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.C.; Guan, S.S.; Yang, H.J.; Chang, C.C.; Luo, T.Y.; Chang, J.; Ho, A.S. Blocking Heme Oxygenase-1 by Zinc Protoporphyrin Reduces Tumor Hypoxia-Mediated VEGF Release and Inhibits Tumor Angiogenesis as a Potential Therapeutic Agent against Colorectal Cancer. J. Biomed. Sci. 2016, 23, 18. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Peng, J.; Wang, H.; Li, L.; Wen, X.; Tan, Y.; Zhang, L.; Wan, H.; Chen, F.; Nie, X. Oxysophocarpine Retards the Growth and Metastasis of Oral Squamous Cell Carcinoma by Targeting the Nrf2/HO-1 Axis. Cell Physiol. Biochem. 2018, 49, 1717–1733. [Google Scholar] [CrossRef]
- Hao, N.B.; Lu, M.H.; Fan, Y.H.; Cao, Y.L.; Zhang, Z.R.; Yang, S.M. Macrophages in Tumor Microenvironments and the Progression of Tumors. Clin. Dev. Immunol. 2012, 2012, 948098. [Google Scholar] [CrossRef]
- Blancou, P.; Anegon, I. Editorial: Heme Oxygenase-1 and Dendritic Cells: What Else? J. Leukoc. Biol. 2010, 87, 185–187. [Google Scholar] [CrossRef]
- Alaluf, E.; Vokaer, B.; Detavernier, A.; Azouz, A.; Splittgerber, M.; Carrette, A.; Boon, L.; Libert, F.; Soares, M.; Le Moine, A.; et al. Heme Oxygenase-1 Orchestrates the Immunosuppressive Program of Tumor-Associated Macrophages. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Nemeth, Z.; Li, M.; Csizmadia, E.; Döme, B.; Johansson, M.; Persson, J.L.; Seth, P.; Otterbein, L.; Wegiel, B. Heme Oxygenase-1 in Macrophages Controls Prostate Cancer Progression. Oncotarget 2015, 6, 33675–33688. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Wang, S.M.; Yin, T.; Ye, T.H.; Shen, G.B.; Li, L.; Zhao, J.Y.; Sang, Y.X.; Duan, X.G.; Wei, Y.Q. Inhibition of Tumor Growth and Alteration of Associated Macrophage Cell Type by an HO-1 Inhibitor in Breast Carcinoma-Bearing Mice. Oncol. Res. 2013, 20, 473–482. [Google Scholar] [CrossRef]
- Beyer, M.; Schultze, J.L. Regulatory T Cells in Cancer. Blood 2006, 108, 804–811. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Chakraborty, N.G.; Mukherji, B. Regulatory T Cells and Tumor Immunity. Cancer Immunol. Immunother. CII 2005, 54, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Kohno, T.; Yamada, Y.; Akamatsu, N.; Kamihira, S.; Imaizumi, Y.; Tomonaga, M.; Matsuyama, T. Possible Origin of Adult T-Cell Leukemia/Lymphoma Cells from Human T Lymphotropic Virus Type-1-Infected Regulatory T Cells. Cancer Sci. 2005, 96, 527–533. [Google Scholar] [CrossRef]
- Di Biase, S.; Lee, C.; Brandhorst, S.; Manes, B.; Buono, R.; Cheng, C.W.; Cacciottolo, M.; Martin-Montalvo, A.; de Cabo, R.; Wei, M.; et al. Fasting-Mimicking Diet Reduces HO-1 to Promote T Cell-Mediated Tumor Cytotoxicity. Cancer Cell 2016, 30, 136–146. [Google Scholar] [CrossRef]
- El Andaloussi, A.; Lesniak, M.S. CD4+ CD25+ FoxP3+ T-Cell Infiltration and Heme Oxygenase-1 Expression Correlate with Tumor Grade in Human Gliomas. J. Neuro-Oncol. 2007, 83, 145–152. [Google Scholar] [CrossRef]
- Orange, J.S. Formation and Function of the Lytic NK-Cell Immunological Synapse. Nat. Rev. Immunol. 2008, 8, 713–725. [Google Scholar] [CrossRef]
- Gomez-Lomeli, P.; Bravo-Cuellar, A.; Hernandez-Flores, G.; Jave-Suarez, L.F.; Aguilar-Lemarroy, A.; Lerma-Diaz, J.M.; Dominguez-Rodriguez, J.R.; Sanchez-Reyes, K.; Ortiz-Lazareno, P.C. Increase of IFN-Gamma and TNF-Alpha Production in CD107a + NK-92 Cells Co-Cultured with Cervical Cancer Cell Lines Pre-Treated with the HO-1 Inhibitor. Cancer Cell Int. 2014, 14, 100. [Google Scholar] [CrossRef] [PubMed]
- Furfaro, A.L.; Ottonello, S.; Loi, G.; Cossu, I.; Piras, S.; Spagnolo, F.; Queirolo, P.; Marinari, U.M.; Moretta, L.; Pronzato, M.A.; et al. HO-1 Downregulation Favors BRAFV600 Melanoma Cell Death Induced by Vemurafenib/PLX4032 and Increases NK Recognition. Int. J. Cancer 2020, 146, 1950–1962. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, H.; Liu, Y.; Duan, C.; Liu, X.; Xia, T.; Chen, D.; Piao, H.-L.; Liu, H.-X. The Double-Edged Roles of ROS in Cancer Prevention and Therapy. Theranostics 2021, 11, 4839–4857. [Google Scholar] [CrossRef]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef]
- Liu, Y.-S.; Li, H.-S.; Qi, D.-F.; Zhang, J.; Jiang, X.-C.; Shi, K.; Zhang, X.-J.; Zhang, X.-H. Zinc Protoporphyrin IX Enhances Chemotherapeutic Response of Hepatoma Cells to Cisplatin. World J. Gastroenterol. 2014, 20, 8572–8582. [Google Scholar] [CrossRef]
- Sun, X.; Wang, S.; Gai, J.; Guan, J.; Li, J.; Li, Y.; Zhao, J.; Zhao, C.; Fu, L.; Li, Q. SIRT5 Promotes Cisplatin Resistance in Ovarian Cancer by Suppressing DNA Damage in a ROS-Dependent Manner via Regulation of the Nrf2/HO-1 Pathway. Front. Oncol. 2019, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Furfaro, A.L.; Macay, J.R.; Marengo, B.; Nitti, M.; Parodi, A.; Fenoglio, D.; Marinari, U.M.; Pronzato, M.A.; Domenicotti, C.; Traverso, N. Resistance of Neuroblastoma GI-ME-N Cell Line to Glutathione Depletion Involves Nrf2 and Heme Oxygenase-1. Free Radic. Biol. Med. 2012, 52, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Wang, H.; Hu, Y.; Hu, M.; Li, X.; Aodengqimuge; Ma, Y.; Wei, C.; Song, L. Src/STAT3-Dependent Heme Oxygenase-1 Induction Mediates Chemoresistance of Breast Cancer Cells to Doxorubicin by Promoting Autophagy. Cancer Sci. 2015, 106, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.; Kong, Y.; Shao, C.; Yue, X.; Wang, Z.; Zhang, N. Heme Oxygenase-1 Induction Mediates Chemoresistance of Breast Cancer Cells to Pharmorubicin by Promoting Autophagy via PI3K/Akt Pathway. J. Cell Mol. Med. 2018, 22, 5311–5321. [Google Scholar] [CrossRef]
- Furfaro, A.L.; Piras, S.; Passalacqua, M.; Domenicotti, C.; Parodi, A.; Fenoglio, D.; Pronzato, M.A.; Marinari, U.M.; Moretta, L.; Traverso, N.; et al. HO-1 up-Regulation: A Key Point in High-Risk Neuroblastoma Resistance to Bortezomib. Biochim. Biophys. Acta 2014, 1842, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Furfaro, A.L.; Piras, S.; Domenicotti, C.; Fenoglio, D.; De Luigi, A.; Salmona, M.; Moretta, L.; Marinari, U.M.; Pronzato, M.A.; Traverso, N.; et al. Role of Nrf2, HO-1 and GSH in Neuroblastoma Cell Resistance to Bortezomib. PLoS ONE 2016, 11, e0152465. [Google Scholar] [CrossRef]
- Barbagallo, I.; Giallongo, C.; Volti, G.L.; Distefano, A.; Camiolo, G.; Raffaele, M.; Salerno, L.; Pittalà, V.; Sorrenti, V.; Avola, R.; et al. Heme Oxygenase Inhibition Sensitizes Neuroblastoma Cells to Carfilzomib. Mol. Neurobiol. 2019, 56, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.Y.; Ahmad, I.M.; Rachagani, S.; Banerjee, K.; Thompson, C.M.; Maurer, H.C.; Olive, K.P.; Bailey, K.L.; Britigan, B.E.; Kumar, S. Enhancing Responsiveness of Pancreatic Cancer Cells to Gemcitabine Treatment under Hypoxia by Heme Oxygenase-1 Inhibition. Transl. Res. 2019, 207, 56–69. [Google Scholar] [CrossRef]
- Ma, J.; Yu, K.N.; Cheng, C.; Ni, G.; Shen, J.; Han, W. Targeting Nrf2-Mediated Heme Oxygenase-1 Enhances Non-Thermal Plasma-Induced Cell Death in Non-Small-Cell Lung Cancer A549 Cells. Arch. Biochem. Biophys. 2018, 658, 54–65. [Google Scholar] [CrossRef]
- Zhang, W.; Qiao, T.; Zha, L. Inhibition of Heme Oxygenase-1 Enhances the Radiosensitivity in Human Nonsmall Cell Lung Cancer A549 Cells. Cancer Biother. Radiopharm. 2011, 26, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Wu, L.; Yuan, H.; Wang, J. ROS/Autophagy/Nrf2 Pathway Mediated Low-Dose Radiation Induced Radio-Resistance in Human Lung Adenocarcinoma A549 Cell. Int. J. Biol. Sci. 2015, 11, 833–844. [Google Scholar] [CrossRef]
- Wang, R.; Shen, J.; Yang, R.; Wang, W.-G.; Yuan, Y.; Guo, Z.-H. Association between Heme Oxygenase-1 Gene Promoter Polymorphisms and Cancer Susceptibility: A Meta-Analysis. Biomed. Rep. 2018, 8, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Li, Z.Y.; Liu, W.; Zhang, R.G.; Li, G.L.; Wang, T.; Ren, J.H.; Wu, G. Polymorphism in Heme Oxygenase-1 (HO-1) Promoter and Alcohol Are Related to the Risk of Esophageal Squamous Cell Carcinoma on Chinese Males. Neoplasma 2010, 57, 86–92. [Google Scholar] [CrossRef][Green Version]
- Kikuchi, A.; Yamaya, M.; Suzuki, S.; Yasuda, H.; Kubo, H.; Nakayama, K.; Handa, M.; Sasaki, T.; Shibahara, S.; Sekizawa, K.; et al. Association of Susceptibility to the Development of Lung Adenocarcinoma with the Heme Oxygenase-1 Gene Promoter Polymorphism. Hum. Genet. 2005, 116, 354–360. [Google Scholar] [CrossRef]
- Murakami, A.; Fujimori, Y.; Yoshikawa, Y.; Yamada, S.; Tamura, K.; Hirayama, N.; Terada, T.; Kuribayashi, K.; Tabata, C.; Fukuoka, K.; et al. Heme Oxygenase-1 Promoter Polymorphism Is Associated with Risk of Malignant Mesothelioma. Lung 2012, 190, 333–337. [Google Scholar] [CrossRef]
- Wu, M.-M.; Lee, C.-H.; Hsu, L.-I.; Cheng, W.-F.; Lee, T.-C.; Wang, Y.-H.; Chiou, H.-Y.; Chen, C.-J. Effect of Heme Oxygenase-1 Gene Promoter Polymorphism on Cancer Risk by Histological Subtype: A Prospective Study in Arseniasis-Endemic Areas in Taiwan. Int. J. Cancer 2016, 138, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xu, Y.; Lu, J.; Xue, J.; Liu, P. High Expression of HO-1 Predicts Poor Prognosis of Ovarian Cancer Patients and Promotes Proliferation and Aggressiveness of Ovarian Cancer Cells. Clin. Transl. Oncol. 2018, 20, 491–499. [Google Scholar] [CrossRef]
- Miyake, M.; Fujimoto, K.; Anai, S.; Ohnishi, S.; Nakai, Y.; Inoue, T.; Matsumura, Y.; Tomioka, A.; Ikeda, T.; Okajima, E.; et al. Inhibition of Heme Oxygenase-1 Enhances the Cytotoxic Effect of Gemcitabine in Urothelial Cancer Cells. Anticancer Res. 2010, 30, 2145–2152. [Google Scholar]
- Kim, J.H.; Park, J. Prognostic Significance of Heme Oxygenase-1, S100 Calcium-Binding Protein A4, and Syndecan-1 Expression in Primary Non-Muscle-Invasive Bladder Cancer. Hum. Pathol. 2014, 45, 1830–1838. [Google Scholar] [CrossRef]
- Gandini, N.A.; Fermento, M.E.; Salomon, D.G.; Obiol, D.J.; Andres, N.C.; Zenklusen, J.C.; Arevalo, J.; Blasco, J.; Lopez Romero, A.; Facchinetti, M.M.; et al. Heme Oxygenase-1 Expression in Human Gliomas and Its Correlation with Poor Prognosis in Patients with Astrocytoma. Tumour Biol. 2014, 35, 2803–2815. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, M.; Zhang, L.; Cai, H.; Zhou, S.; Zhang, J.; Wang, Y. Correlation of Nrf2, HO-1, and MRP3 in Gallbladder Cancer and Their Relationships to Clinicopathologic Features and Survival. J. Surg. Res. 2010, 164, e99–e105. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, Y.; Zhao, P.; Weng, W.; Ye, M.; Sun, H.; Xu, M.; Wang, C. The Nrf2/HO-1 Axis Can Be a Prognostic Factor in Clear Cell Renal Cell Carcinoma. Cancer Manag. Res. 2019, 11, 1221–1230. [Google Scholar] [CrossRef]
- Kongpetch, S.; Puapairoj, A.; Ong, C.K.; Senggunprai, L.; Prawan, A.; Kukongviriyapan, U.; Chan-On, W.; Siew, E.Y.; Khuntikeo, N.; Teh, B.T.; et al. Haem Oxygenase 1 Expression Is Associated with Prognosis in Cholangiocarcinoma Patients and with Drug Sensitivity in Xenografted Mice. Cell Prolif. 2016, 49, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, J.; Ma, D.; Chen, S.; Lin, X.; Fang, Q.; Zhe, N. HO-1, RET and PML as Possible Markers for Risk Stratification of Acute Myelocytic Leukemia and Prognostic Evaluation. Oncol. Lett. 2015, 10, 3137–3144. [Google Scholar] [CrossRef] [PubMed]
- Fest, S.; Soldati, R.; Christiansen, N.M.; Zenclussen, M.L.; Kilz, J.; Berger, E.; Starke, S.; Lode, H.N.; Engel, C.; Zenclussen, A.C.; et al. Targeting of Heme Oxygenase-1 as a Novel Immune Regulator of Neuroblastoma. Int. J. Cancer 2016, 138, 2030–2042. [Google Scholar] [CrossRef]
- Wei, S.; Wang, Y.; Chai, Q.; Fang, Q.; Zhang, Y.; Lu, Y.; Wang, J. Over-Expression of Heme Oxygenase-1 in Peripheral Blood Predicts the Progression and Relapse Risk of Chronic Myeloid Leukemia. Chin. Med. J. 2014, 127, 2795–2801. [Google Scholar] [PubMed]
- Zhe, N.; Wang, J.; Chen, S.; Lin, X.; Chai, Q.; Zhang, Y.; Zhao, J.; Fang, Q. Heme Oxygenase-1 Plays a Crucial Role in Chemoresistance in Acute Myeloid Leukemia. Hematology 2015, 20, 384–391. [Google Scholar] [CrossRef]
- He, Z.; Zhang, S.; Ma, D.; Fang, Q.; Yang, L.; Shen, S.; Chen, Y.; Ren, L.; Wang, J. HO-1 Promotes Resistance to an EZH2 Inhibitor through the PRB-E2F Pathway: Correlation with the Progression of Myelodysplastic Syndrome into Acute Myeloid Leukemia. J. Transl. Med. 2019, 17, 366. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-Y.; Liu, C.-L.; Chen, M.-J.; Lee, J.-J.; Pun, P.C.; Cheng, S.-P. Expression of Haem Oxygenase-1 Correlates with Tumour Aggressiveness and BRAF V600E Expression in Thyroid Cancer. Histopathology 2015, 66, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Su, J.; DingZhang, X.; Zhang, J.; Yoshimoto, M.; Liu, S.; Bijian, K.; Gupta, A.; Squire, J.A.; Alaoui Jamali, M.A.; et al. PTEN Deletion and Heme Oxygenase-1 Overexpression Cooperate in Prostate Cancer Progression and Are Associated with Adverse Clinical Outcome. J. Pathol. 2011, 224, 90–100. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, Y.; Huang, Y.; Ma, Q.; Shang, J.; Guo, J.; Cao, X.; Wang, X.; Li, M. Inhibition of Nrf2/HO-1 Signaling Pathway by Dextran Sulfate Suppresses Angiogenesis of Gastric Cancer. J. Cancer 2021, 12, 1042–1060. [Google Scholar] [CrossRef] [PubMed]
- Park, C.-S.; Eom, D.-W.; Ahn, Y.; Jang, H.J.; Hwang, S.; Lee, S.-G. Can Heme Oxygenase-1 Be a Prognostic Factor in Patients with Hepatocellular Carcinoma? Medicine 2019, 98, e16084. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wu, Y.; Bian, C.; Nisar, M.F.; Wang, M.; Hu, X.; Diao, Q.; Nian, W.; Wang, E.; Xu, W.; et al. Heme Oxygenase 1 Facilitates Cell Proliferation via the B-Raf-ERK Signaling Pathway in Melanoma. Cell Commun. Signal. 2019, 17, 3. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Miyata, Y.; Mitsunari, K.; Yasuda, T.; Ohba, K.; Sakai, H. Pathological Significance and Prognostic Implications of Heme Oxygenase 1 Expression in Non-Muscle-Invasive Bladder Cancer: Correlation with Cell Proliferation, Angiogenesis, Lymphangiogenesis and Expression of VEGFs and COX-2. Oncol. Lett. 2017, 13, 275–280. [Google Scholar] [CrossRef]
- Nishie, A.; Ono, M.; Shono, T.; Fukushi, J.; Otsubo, M.; Onoue, H.; Ito, Y.; Inamura, T.; Ikezaki, K.; Fukui, M.; et al. Macrophage Infiltration and Heme Oxygenase-1 Expression Correlate with Angiogenesis in Human Gliomas. Clin. Cancer Res. 1999, 5, 1107–1113. [Google Scholar]
- Kimura, S.; Aung, N.Y.; Ohe, R.; Yano, M.; Hashimoto, T.; Fujishima, T.; Kimura, W.; Yamakawa, M. Increasing Heme Oxygenase-1-Expressing Macrophages Indicates a Tendency of Poor Prognosis in Advanced Colorectal Cancer. Digestion 2020, 101, 401–410. [Google Scholar] [CrossRef]
- Becker, J.C.; Fukui, H.; Imai, Y.; Sekikawa, A.; Kimura, T.; Yamagishi, H.; Yoshitake, N.; Pohle, T.; Domschke, W.; Fujimori, T. Colonic Expression of Heme Oxygenase-1 Is Associated with a Better Long-Term Survival in Patients with Colorectal Cancer. Scand. J. Gastroenterol. 2007, 42, 852–858. [Google Scholar] [CrossRef]
- Andrés, N.C.; Fermento, M.E.; Gandini, N.A.; Romero, A.L.; Ferro, A.; Donna, L.G.; Curino, A.C.; Facchinetti, M.M. Heme Oxygenase-1 Has Antitumoral Effects in Colorectal Cancer: Involvement of P53. Exp. Mol. Pathol. 2014, 97, 321–331. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, Q.; Wang, B.; Chen, G.; Xu, L.; Zhou, H. Expression and Function of Heme Oxygenase-1 in Human Gastric Cancer. Exp Biol. Med. 2012, 237, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.-Y.; Hong, S.-M.; Bae, Y.K.; Kim, H.K.; Jang, K.Y.; Eom, D.W. Clinicopathological and Prognostic Significance of Heme Oxygenase-1 Expression in Small Intestinal Adenocarcinomas. Pathol. Int. 2018, 68, 294–300. [Google Scholar] [CrossRef]
- Tsuji, M.H.; Yanagawa, T.; Iwasa, S.; Tabuchi, K.; Onizawa, K.; Bannai, S.; Toyooka, H.; Yoshida, H. Heme Oxygenase-1 Expression in Oral Squamous Cell Carcinoma as Involved in Lymph Node Metastasis. Cancer Lett. 1999, 138, 53–59. [Google Scholar] [CrossRef]
- Gandini, N.A.; Fermento, M.E.; Salomon, D.G.; Blasco, J.; Patel, V.; Gutkind, J.S.; Molinolo, A.A.; Facchinetti, M.M.; Curino, A.C. Nuclear Localization of Heme Oxygenase-1 Is Associated with Tumor Progression of Head and Neck Squamous Cell Carcinomas. Exp. Mol. Pathol. 2012, 93, 237–245. [Google Scholar] [CrossRef]
- Gandini, N.A.; Alonso, E.N.; Fermento, M.E.; Mascaró, M.; Abba, M.C.; Coló, G.P.; Arévalo, J.; Ferronato, M.J.; Guevara, J.A.; Núñez, M.; et al. Heme Oxygenase-1 Has an Antitumor Role in Breast Cancer. Antioxid. Redox Signal. 2019, 30, 2030–2049. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Fang, J.; Liao, L.; Maeda, H.; Su, Q. Upregulation of Heme Oxygenase-1 in Colorectal Cancer Patients with Increased Circulation Carbon Monoxide Levels, Potentially Affects Chemotherapeutic Sensitivity. BMC Cancer 2014, 14, 436. [Google Scholar] [CrossRef]
- Alaoui-Jamali, M.A.; Bismar, T.A.; Gupta, A.; Szarek, W.A.; Su, J.; Song, W.; Xu, Y.; Xu, B.; Liu, G.; Vlahakis, J.Z.; et al. A Novel Experimental Heme Oxygenase-1-Targeted Therapy for Hormone-Refractory Prostate Cancer. Cancer Res. 2009, 69, 8017–8024. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, H.; Wang, S.; Zhu, J. Expression and Correlation of NRF2, KEAP1, NQO-1 and HO-1 in Advanced Squamous Cell Carcinoma of the Larynx and Their Association with Clinicopathologic Features. Mol. Med. Rep. 2016, 14, 5171–5179. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Lu, T.; Ma, D.; Yu, K.; Li, X.; Chen, B.; Xiong, J.; Zhang, T.; Wang, J. Heme Oxygenase-1 Reduces the Sensitivity to Imatinib through Nonselective Activation of Histone Deacetylases in Chronic Myeloid Leukemia. J. Cell Physiol. 2019, 234, 5252–5263. [Google Scholar] [CrossRef]
- Cheng, B.; Tang, S.; Zhe, N.; Ma, D.; Yu, K.; Wei, D.; Zhou, Z.; Lu, T.; Wang, J.; Fang, Q. Low Expression of GFI-1 Gene Is Associated with Panobinostat-Resistance in Acute Myeloid Leukemia through Influencing the Level of HO-1. Biomed. Pharmacother. 2018, 100, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Fang, Q.; Chen, S.; Zhe, N.; Chai, Q.; Yu, M.; Zhang, Y.; Wang, Z.; Wang, J. Heme Oxygenase-1 Suppresses the Apoptosis of Acute Myeloid Leukemia Cells via the JNK/c-JUN Signaling Pathway. Leuk. Res. 2015, 39, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Fang, Q.; Wang, P.; Gao, R.; Sun, J.; Li, Y.; Hu, X.Y.; Wang, J.S. Downregulation of HO-1 Promoted Apoptosis Induced by Decitabine via Increasing P15INK4B Promoter Demethylation in Myelodysplastic Syndrome. Gene Ther. 2015, 22, 287–296. [Google Scholar] [CrossRef]
- Barrera, L.N.; Rushworth, S.A.; Bowles, K.M.; MacEwan, D.J. Bortezomib Induces Heme Oxygenase-1 Expression in Multiple Myeloma. Cell Cycle 2012, 11, 2248–2252. [Google Scholar] [CrossRef]
- Podkalicka, P.; Mucha, O.; Józkowicz, A.; Dulak, J.; Łoboda, A. Heme Oxygenase Inhibition in Cancers: Possible Tools and Targets. Contemp. Oncol. 2018, 22, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Vreman, H.J.; Ekstrand, B.C.; Stevenson, D.K. Selection of Metalloporphyrin Heme Oxygenase Inhibitors Based on Potency and Photoreactivity. Pediatr. Res. 1993, 33, 195–200. [Google Scholar] [CrossRef]
- Schulz, S.; Wong, R.J.; Vreman, H.J.; Stevenson, D.K. Metalloporphyrins—An Update. Front. Pharmacol. 2012, 3, 68. [Google Scholar] [CrossRef]
- Shi, L.; Fang, J. Implication of Heme Oxygenase-1 in the Sensitivity of Nasopharyngeal Carcinomas to Radiotherapy. J. Exp. Clin. Cancer Res. 2008, 27, 13. [Google Scholar] [CrossRef] [PubMed]
- Nowis, D.; Legat, M.; Grzela, T.; Niderla, J.; Wilczek, E.; Wilczynski, G.M.; Glodkowska, E.; Mrowka, P.; Issat, T.; Dulak, J.; et al. Heme Oxygenase-1 Protects Tumor Cells against Photodynamic Therapy-Mediated Cytotoxicity. Oncogene 2006, 25, 3365–3374. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.; Lornejad-Schäfer, M.R.; Schöffl, H.; Flaccus, A.; Lambert, C.; Biesalski, H.K. Inhibition of Heme Oxygenase-1 Increases Responsiveness of Melanoma Cells to ALA-Based Photodynamic Therapy. Int. J. Oncol. 2007, 31, 1539–1545. [Google Scholar] [CrossRef]
- Appleton, S.D.; Chretien, M.L.; McLaughlin, B.E.; Vreman, H.J.; Stevenson, D.K.; Brien, J.F.; Nakatsu, K.; Maurice, D.H.; Marks, G.S. Selective Inhibition of Heme Oxygenase, without Inhibition of Nitric Oxide Synthase or Soluble Guanylyl Cyclase, by Metalloporphyrins at Low Concentrations. Drug Metab. Dispos. 1999, 27, 1214–1219. [Google Scholar]
- Kinobe, R.T.; Dercho, R.A.; Nakatsu, K. Inhibitors of the Heme Oxygenase-Carbon Monoxide System: On the Doorstep of the Clinic? Can. J. Physiol. Pharmacol. 2008, 86, 577–599. [Google Scholar] [CrossRef] [PubMed]
- Sardana, M.K.; Kappas, A. Dual Control Mechanism for Heme Oxygenase: Tin(IV)-Protoporphyrin Potently Inhibits Enzyme Activity While Markedly Increasing Content of Enzyme Protein in Liver. Proc. Natl. Acad. Sci. USA 1987, 84, 2464–2468. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Nguyen, X.; Ou, J.; Rekulapelli, P.; Stevenson, D.K.; Dennery, P.A. Unique Effects of Zinc Protoporphyrin on HO-1 Induction and Apoptosis. Blood 2001, 97, 1306–1313. [Google Scholar] [CrossRef]
- Kwok, S.C.M. Zinc Protoporphyrin Upregulates Heme Oxygenase-1 in PC-3 Cells via the Stress Response Pathway. Int. J. Cell Biol. 2013, 2013, 162094. [Google Scholar] [CrossRef]
- Sahoo, S.K.; Sawa, T.; Fang, J.; Tanaka, S.; Miyamoto, Y.; Akaike, T.; Maeda, H. Pegylated Zinc Protoporphyrin: A Water-Soluble Heme Oxygenase Inhibitor with Tumor-Targeting Capacity. Bioconjugate Chem. 2002, 13, 1031–1038. [Google Scholar] [CrossRef]
- Iyer, A.K.; Greish, K.; Seki, T.; Okazaki, S.; Fang, J.; Takeshita, K.; Maeda, H. Polymeric Micelles of Zinc Protoporphyrin for Tumor Targeted Delivery Based on EPR Effect and Singlet Oxygen Generation. J. Drug Target. 2007, 15, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.K.; Greish, K.; Fang, J.; Murakami, R.; Maeda, H. High-Loading Nanosized Micelles of Copoly(Styrene-Maleic Acid)-Zinc Protoporphyrin for Targeted Delivery of a Potent Heme Oxygenase Inhibitor. Biomaterials 2007, 28, 1871–1881. [Google Scholar] [CrossRef]
- Fang, J.; Sawa, T.; Akaike, T.; Akuta, T.; Sahoo, S.K.; Khaled, G.; Hamada, A.; Maeda, H. In Vivo Antitumor Activity of Pegylated Zinc Protoporphyrin: Targeted Inhibition of Heme Oxygenase in Solid Tumor. Cancer Res. 2003, 63, 3567–3574. [Google Scholar] [PubMed]
- Herrmann, H.; Kneidinger, M.; Cerny-Reiterer, S.; Rülicke, T.; Willmann, M.; Gleixner, K.V.; Blatt, K.; Hörmann, G.; Peter, B.; Samorapoompichit, P.; et al. The Hsp32 Inhibitors SMA-ZnPP and PEG-ZnPP Exert Major Growth-Inhibitory Effects on D34+/CD38+ and CD34+/CD38- AML Progenitor Cells. Curr. Cancer Drug Targets 2012, 12, 51–63. [Google Scholar] [CrossRef]
- Kinobe, R.T.; Vlahakis, J.Z.; Vreman, H.J.; Stevenson, D.K.; Brien, J.F.; Szarek, W.A.; Nakatsu, K. Selectivity of Imidazole-Dioxolane Compounds for in Vitro Inhibition of Microsomal Haem Oxygenase Isoforms. Br. J. Pharmacol. 2006, 147, 307–315. [Google Scholar] [CrossRef]
- Pittala, V.; Salerno, L.; Romeo, G.; Modica, M.N.; Siracusa, M.A. A Focus on Heme Oxygenase-1 (HO-1) Inhibitors. Curr. Med. Chem. 2013, 20, 3711–3732. [Google Scholar] [CrossRef] [PubMed]
- Vlahakis, J.Z.; Kinobe, R.T.; Bowers, R.J.; Brien, J.F.; Nakatsu, K.; Szarek, W.A. Synthesis and Evaluation of Azalanstat Analogues as Heme Oxygenase Inhibitors. Bioorg. Med. Chem. Lett. 2005, 15, 1457–1461. [Google Scholar] [CrossRef]
- Salerno, L.; Floresta, G.; Ciaffaglione, V.; Gentile, D.; Margani, F.; Turnaturi, R.; Rescifina, A.; Pittalà, V. Progress in the Development of Selective Heme Oxygenase-1 Inhibitors and Their Potential Therapeutic Application. Eur. J. Med. Chem. 2019, 167, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Ciaffaglione, V.; Intagliata, S.; Pittalà, V.; Marrazzo, A.; Sorrenti, V.; Vanella, L.; Rescifina, A.; Floresta, G.; Sultan, A.; Greish, K.; et al. New Arylethanolimidazole Derivatives as HO-1 Inhibitors with Cytotoxicity against MCF-7 Breast Cancer Cells. Int. J. Mol. Sci. 2020, 21, 1923. [Google Scholar] [CrossRef]
- Salerno, L.; Pittalà, V.; Romeo, G.; Modica, M.N.; Marrazzo, A.; Siracusa, M.A.; Sorrenti, V.; Di Giacomo, C.; Vanella, L.; Parayath, N.N.; et al. Novel Imidazole Derivatives as Heme Oxygenase-1 (HO-1) and Heme Oxygenase-2 (HO-2) Inhibitors and Their Cytotoxic Activity in Human-Derived Cancer Cell Lines. Eur. J. Med. Chem. 2015, 96, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Berberat, P.O.; Dambrauskas, Z.; Gulbinas, A.; Giese, T.; Giese, N.; Kunzli, B.; Autschbach, F.; Meuer, S.; Buchler, M.W.; Friess, H. Inhibition of Heme Oxygenase-1 Increases Responsiveness of Pancreatic Cancer Cells to Anticancer Treatment. Clin. Cancer Res. 2005, 11, 3790–3798. [Google Scholar] [CrossRef]
- Sass, G.; Leukel, P.; Schmitz, V.; Raskopf, E.; Ocker, M.; Neureiter, D.; Meissnitzer, M.; Tasika, E.; Tannapfel, A.; Tiegs, G. Inhibition of Heme Oxygenase 1 Expression by Small Interfering RNA Decreases Orthotopic Tumor Growth in Livers of Mice. Int. J. Cancer 2008, 123, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Cerny-Reiterer, S.; Meyer, R.A.; Herrmann, H.; Peter, B.; Gleixner, K.V.; Stefanzl, G.; Hadzijusufovic, E.; Pickl, W.F.; Sperr, W.R.; Melo, J.V.; et al. Identification of Heat Shock Protein 32 (Hsp32) as a Novel Target in Acute Lymphoblastic Leukemia. Oncotarget 2014, 5, 1198–1211. [Google Scholar] [CrossRef]
- Zhou, Z.; Fang, Q.; Li, P.; Ma, D.; Zhe, N.; Ren, M.; Chen, B.; He, Z.; Wang, J.; Zhong, Q.; et al. Entinostat Combined with Fludarabine Synergistically Enhances the Induction of Apoptosis in TP53 Mutated CLL Cells via the HDAC1/HO-1 Pathway. Life Sci. 2019, 232, 116583. [Google Scholar] [CrossRef]
- Zhou, Z.; Ma, D.; Liu, P.; Wang, P.; Wei, D.; Yu, K.; Li, P.; Fang, Q.; Wang, J. Deletion of HO-1 Blocks Development of B Lymphocytes in Mice. Cell Signal. 2019, 63, 109378. [Google Scholar] [CrossRef] [PubMed]
- Skrzypek, K.; Tertil, M.; Golda, S.; Ciesla, M.; Weglarczyk, K.; Collet, G.; Guichard, A.; Kozakowska, M.; Boczkowski, J.; Was, H.; et al. Interplay between Heme Oxygenase-1 and MiR-378 Affects Non-Small Cell Lung Carcinoma Growth, Vascularization, and Metastasis. Antioxid. Redox Signal. 2013, 19, 644–660. [Google Scholar] [CrossRef] [PubMed]
- Tertil, M.; Golda, S.; Skrzypek, K.; Florczyk, U.; Weglarczyk, K.; Kotlinowski, J.; Maleszewska, M.; Czauderna, S.; Pichon, C.; Kieda, C.; et al. Nrf2-Heme Oxygenase-1 Axis in Mucoepidermoid Carcinoma of the Lung: Antitumoral Effects Associated with down-Regulation of Matrix Metalloproteinases. Free Radic. Biol. Med. 2015, 89, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Cheng, W.; Li, Q.; Han, Z.; Wang, X.; Jin, J.; Zou, J.; Liu, Z.; Zhou, Z.; Zhao, W.; et al. Heme Oxygenase-1 Retards Hepatocellular Carcinoma Progression through the MicroRNA Pathway. Oncol. Rep. 2016, 36, 2715–2722. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Pereira, V.; Chauveau, C.; Zagani, R.; Remy, S.; Tesson, L.; Mazal, D.; Ubillos, L.; Brion, R.; Asghar, K.; et al. Heme Oxygenase-1 Inhibits Rat and Human Breast Cancer Cell Proliferation: Mutual Cross Inhibition with Indoleamine 2,3-Dioxygenase. FASEB J. 2005, 19, 1957–1968. [Google Scholar] [CrossRef]
- Gueron, G.; Giudice, J.; Valacco, P.; Paez, A.; Elguero, B.; Toscani, M.; Jaworski, F.; Leskow, F.C.; Cotignola, J.; Marti, M.; et al. Heme-Oxygenase-1 Implications in Cell Morphology and the Adhesive Behavior of Prostate Cancer Cells. Oncotarget 2014, 5, 4087–4102. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.N.; Park, G.H.; Park, S.B.; Kim, J.D.; Eo, H.J.; Son, H.-J.; Song, J.H.; Jeong, J.B. Extracts from Sageretia Thea Reduce Cell Viability through Inducing Cyclin D1 Proteasomal Degradation and HO-1 Expression in Human Colorectal Cancer Cells. BMC Complement. Altern. Med. 2019, 19, 43. [Google Scholar] [CrossRef]
- Bi, W.; He, C.-N.; Li, X.-X.; Zhou, L.-Y.; Liu, R.-J.; Zhang, S.; Li, G.-Q.; Chen, Z.-C.; Zhang, P.-F. Ginnalin A from Kujin Tea (Acer tataricum subsp. ginnala) Exhibits a Colorectal Cancer Chemoprevention Effect via Activation of the Nrf2/HO-1 Signaling Pathway. Food Funct. 2018, 9, 2809–2819. [Google Scholar] [CrossRef]
- Tsai, C.-F.; Chen, J.-H.; Chang, C.-N.; Lu, D.-Y.; Chang, P.-C.; Wang, S.-L.; Yeh, W.-L. Fisetin Inhibits Cell Migration via Inducing HO-1 and Reducing MMPs Expression in Breast Cancer Cell Lines. Food Chem. Toxicol. 2018, 120, 528–535. [Google Scholar] [CrossRef]
- Kim, M.J.; Yun, G.J.; Kim, S.E. Metabolic Regulation of Ferroptosis in Cancer. Biology 2021, 10, 83. [Google Scholar] [CrossRef]
- Greco, G.; Catanzaro, E.; Fimognari, C. Natural Products as Inducers of Non-Canonical Cell Death: A Weapon against Cancer. Cancers 2021, 13, 304. [Google Scholar] [CrossRef] [PubMed]
- Hassannia, B.; Wiernicki, B.; Ingold, I.; Qu, F.; Van Herck, S.; Tyurina, Y.Y.; Bayır, H.; Abhari, B.A.; Angeli, J.P.F.; Choi, S.M.; et al. Nano-Targeted Induction of Dual Ferroptotic Mechanisms Eradicates High-Risk Neuroblastoma. J. Clin. Investig. 2018, 128, 3341–3355. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; Genovese, C.; La Mantia, A.; Cammarata, F.P.; Ragusa, M.; Renis, M.; Di Giacomo, C. Betula Etnensis Raf. (Betulaceae) Extract Induced HO-1 Expression and Ferroptosis Cell Death in Human Colon Cancer Cells. Int. J. Mol. Sci. 2019, 20, 2723. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, J.; Zhou, Y.; Gao, Q.; Wang, R.; Fu, Y.; Zheng, L.; Yu, H. Transcriptome Investigation and In Vitro Verification of Curcumin-Induced HO-1 as a Feature of Ferroptosis in Breast Cancer Cells. Oxid. Med. Cell. Longev. 2020, 2020, 3469840. [Google Scholar] [CrossRef]
- Chen, P.; Li, X.; Zhang, R.; Liu, S.; Xiang, Y.; Zhang, M.; Chen, X.; Pan, T.; Yan, L.; Feng, J.; et al. Combinative Treatment of β-Elemene and Cetuximab Is Sensitive to KRAS Mutant Colorectal Cancer Cells by Inducing Ferroptosis and Inhibiting Epithelial-Mesenchymal Transformation. Theranostics 2020, 10, 5107–5119. [Google Scholar] [CrossRef] [PubMed]
- Zager, R.A.; Johnson, A.C.M.; Guillem, A.; Keyser, J.; Singh, B. A Pharmacologic “Stress Test” for Assessing Select Antioxidant Defenses in Patients with CKD. Clin. J. Am. Soc. Nephrol. 2020, 15, 633–642. [Google Scholar] [CrossRef]
- Bhutani, V.K.; Poland, R.; Meloy, L.D.; Hegyi, T.; Fanaroff, A.A.; Maisels, M.J. Clinical Trial of Tin Mesoporphyrin to Prevent Neonatal Hyperbilirubinemia. J. Perinatol. 2016, 36, 533–539. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nitti, M.; Ivaldo, C.; Traverso, N.; Furfaro, A.L. Clinical Significance of Heme Oxygenase 1 in Tumor Progression. Antioxidants 2021, 10, 789. https://doi.org/10.3390/antiox10050789
Nitti M, Ivaldo C, Traverso N, Furfaro AL. Clinical Significance of Heme Oxygenase 1 in Tumor Progression. Antioxidants. 2021; 10(5):789. https://doi.org/10.3390/antiox10050789
Chicago/Turabian StyleNitti, Mariapaola, Caterina Ivaldo, Nicola Traverso, and Anna Lisa Furfaro. 2021. "Clinical Significance of Heme Oxygenase 1 in Tumor Progression" Antioxidants 10, no. 5: 789. https://doi.org/10.3390/antiox10050789
APA StyleNitti, M., Ivaldo, C., Traverso, N., & Furfaro, A. L. (2021). Clinical Significance of Heme Oxygenase 1 in Tumor Progression. Antioxidants, 10(5), 789. https://doi.org/10.3390/antiox10050789