
PEG35 and Glutathione Improve Mitochondrial Function and Reduce Oxidative Stress in Cold Fatty Liver Graft Preservation

 ,
, 


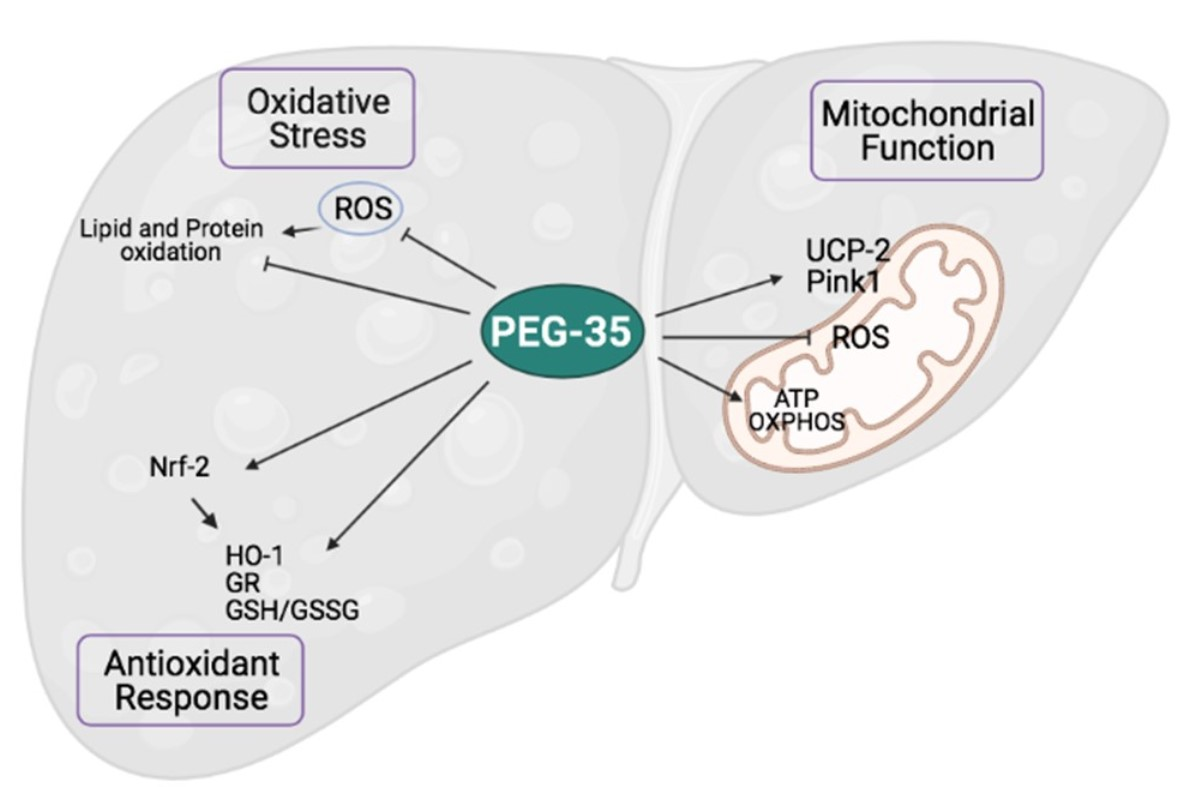
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Liver Isolation and Perfusion
2.2. ATP and Succinate Determination
2.3. Oxidant and Antioxidant Assays in Liver
2.3.1. Lipid and Protein Oxidation
2.3.2. Antioxidant Enzymes Activity
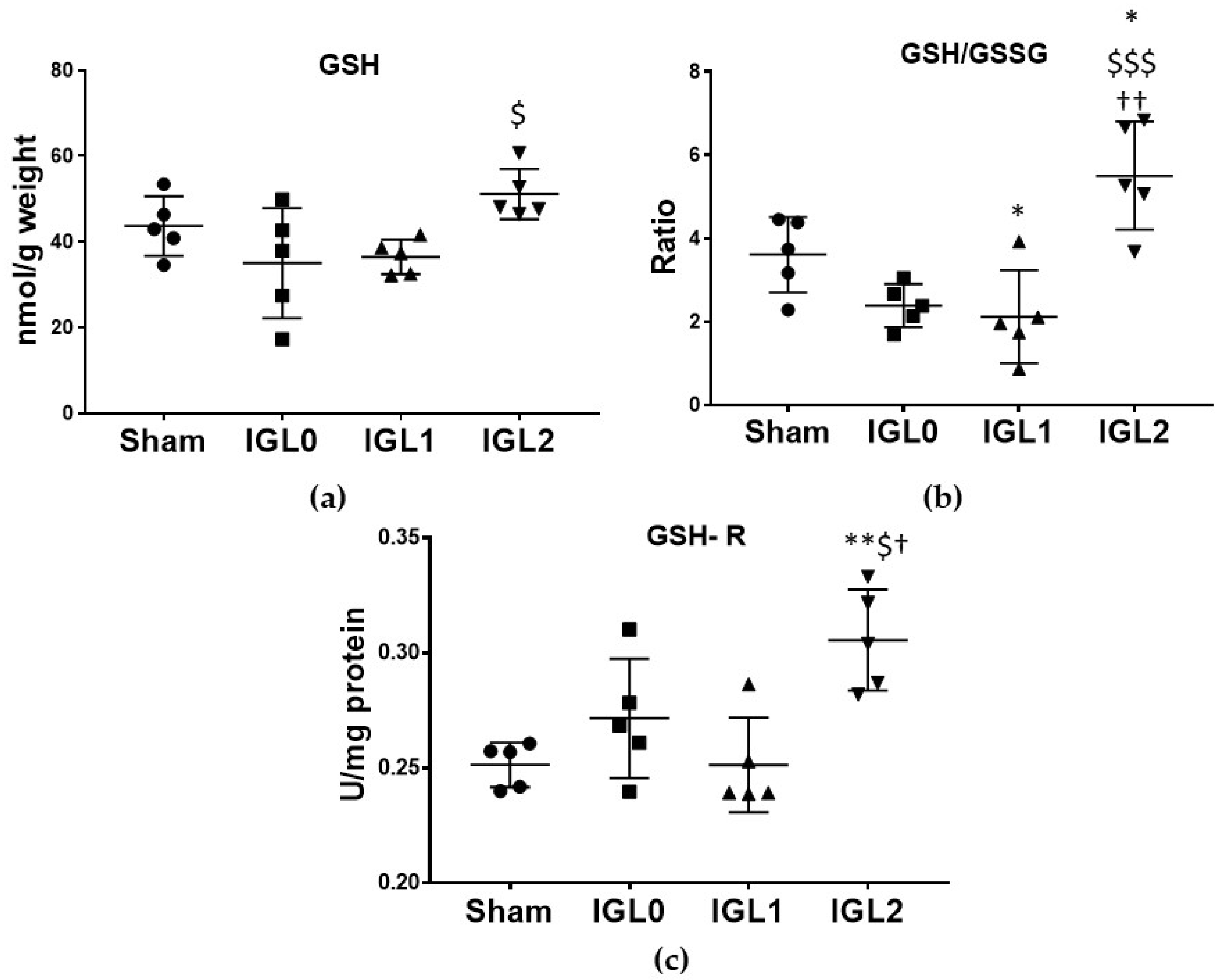
2.3.3. GSH
2.4. Western Blotting Analysis and Quantification
2.5. Statistical Analysis
3. Results
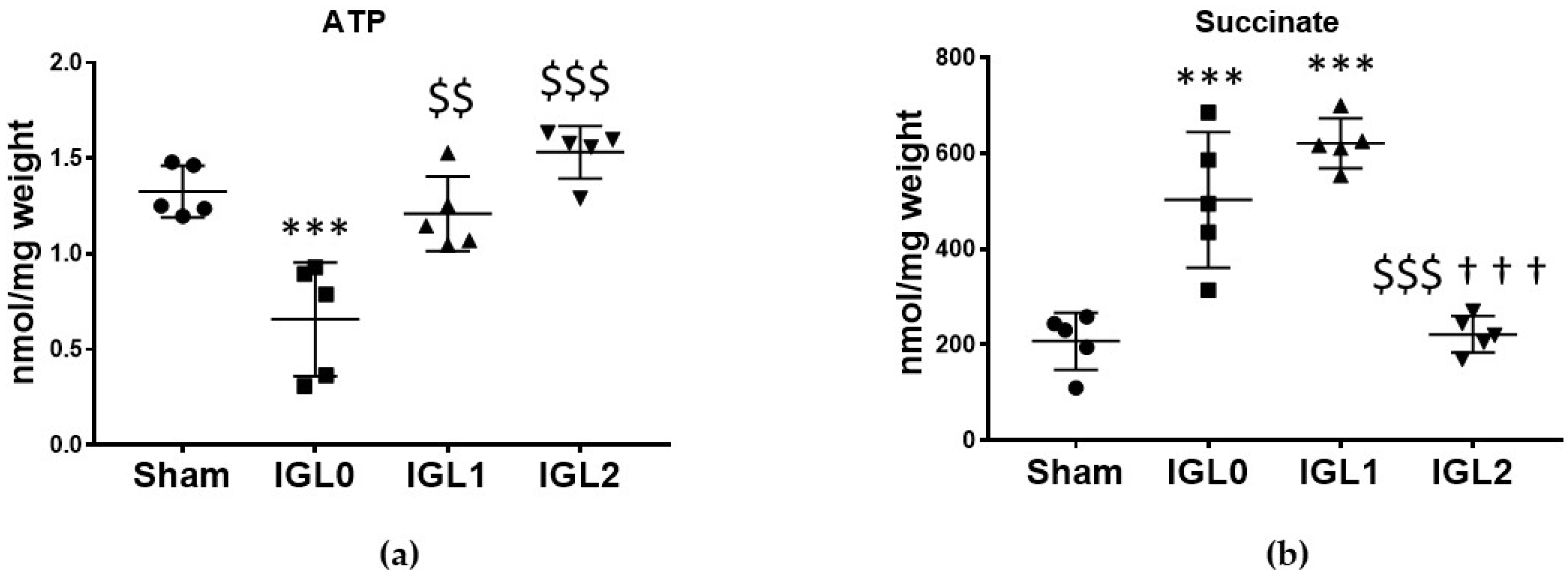
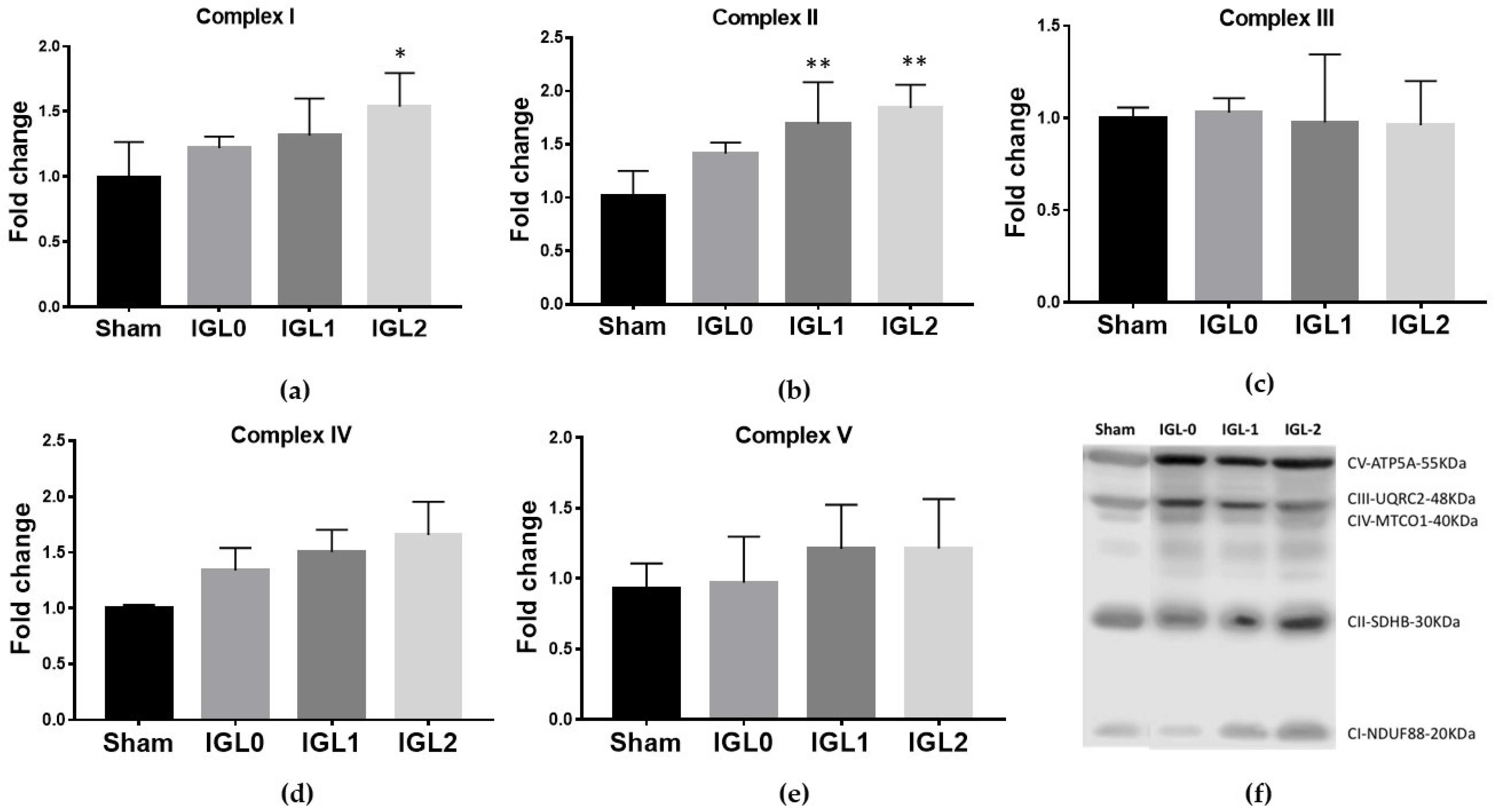
3.1. IGL-2 Prevents ATP Breakdown and Succinate Accumulation and Affects Mitochondrial OXPHOS Complex Expression
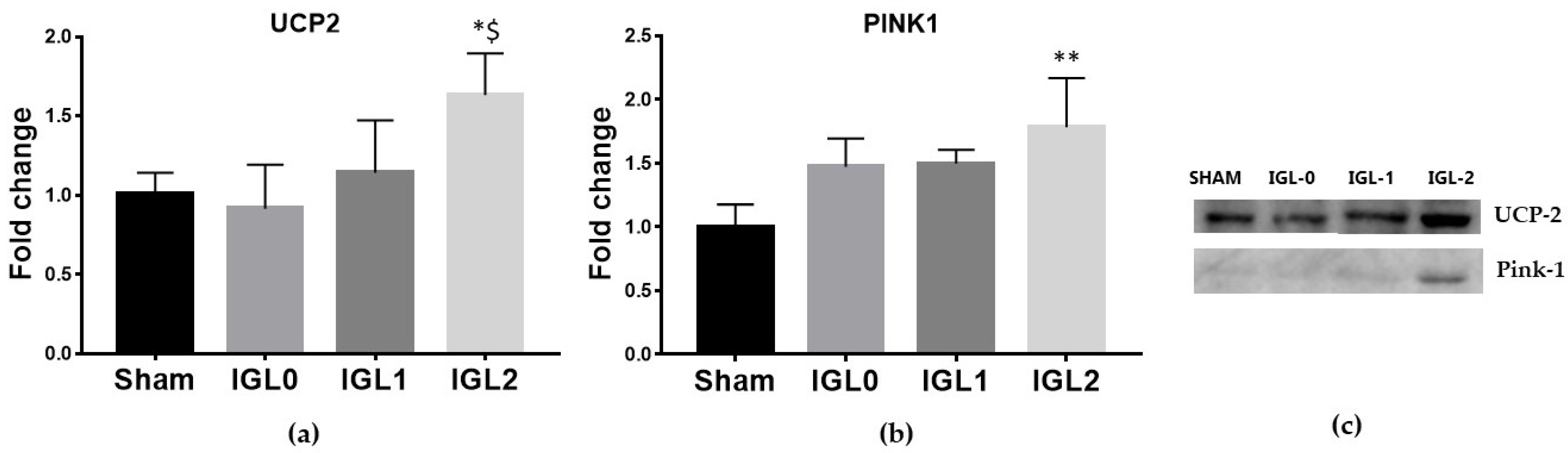
3.2. UCP2 Expression and Mitophagy Increases in IGL-2 Preservation
3.3. Nrf2 Transcription Factor Increases in IGL-2 Solution Preservation
3.4. IGL-2 Preservation Solution Better Protects Steatotic Livers from Oxidative Stress
3.5. Antioxidant Enzymatic Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cholankeril, G.; Wong, R.J.; Hu, M.; Perumpail, R.B.; Yoo, E.R.; Puri, P.; Younossi, Z.M.; Harrison, S.A.; Ahmed, A. Liver Transplantation for Nonalcoholic Steatohepatitis in the US: Temporal Trends and Outcomes. Dig. Dis. Sci. 2017, 62, 2915–2922. [Google Scholar] [CrossRef]
- Peralta, C.; Roselló-Catafau, J. The Future of Fatty Livers. J. Hepatol. 2004, 41, 149–151. [Google Scholar] [CrossRef]
- Tashiro, H.; Kuroda, S.; Mikuriya, Y.; Ohdan, H. Ischemia-Reperfusion Injury in Patients with Fatty Liver and the Clinical Impact of Steatotic Liver on Hepatic Surgery. Surg. Today 2014, 44, 1611–1625. [Google Scholar] [CrossRef]
- Bejaoui, M.; Pantazi, E.; Folch-Puy, E.; Baptista, P.M.; García-Gil, A.; Adam, R.; Roselló-Catafau, J. Emerging Concepts in Liver Graft Preservation. World J. Gastroenterol. 2015, 21, 396–407. [Google Scholar] [CrossRef] [Green Version]
- Zaouali, M.A.; Ben Abdennebi, H.; Padrissa-Altés, S.; Mahfoudh-Boussaid, A.; Roselló-Catafau, J. Pharmacological Strategies against Cold Ischemia Reperfusion Injury. Expert Opin. Pharmacother. 2010, 11, 537–555. [Google Scholar] [CrossRef] [PubMed]
- Guibert, E.E.; Petrenko, A.Y.; Balaban, C.L.; Somov, A.Y.; Rodriguez, J.V.; Fuller, B.J. Organ Preservation: Current Concepts and New Strategies for the next Decade. Transfus. Med. Hemotherapy 2011, 38, 125–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadzuka, Y. Biological and Pharmaceutical Bulletin: Foreword. Biol. Pharm. Bull. 2013, 36, 877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puts, C.F.; Berendsen, T.A.; Bruinsma, B.G.; Ozer, S.; Luitje, M.; Usta, O.B.; Yarmush, M.L.; Uygun, K. Polyethylene Glycol Protects Primary Hepatocytes during Supercooling Preservation. Cryobiology 2015, 71, 125–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch-Puy, E.; Panisello, A.; Oliva, J.; Lopez, A.; Benítez, C.C.; Adam, R.; Roselló-Catafau, J. Relevance of Endoplasmic Reticulum Stress Cell Signaling in Liver Cold Ischemia Reperfusion Injury. Int. J. Mol. Sci. 2016, 17, 807. [Google Scholar] [CrossRef] [Green Version]
- Zaouali, M.A.; Bejaoui, M.; Calvo, M.; Folch-Puy, E.; Pantazi, E.; Pasut, G.; Rimola, A.; Ben Abdennebi, H.; Adam, R.; Roselló-Catafau, J. Polyethylene Glycol Rinse Solution: An Effective Way to Prevent Ischemia-Reperfusion Injury. World J. Gastroenterol. 2014, 20, 16203–16214. [Google Scholar] [CrossRef]
- Panisello-rosell, A.; Verde, E.; Zaouali, M.A.M.A.; Flores, M.; Alva, N.; Lopez, A.; Folch-puy, E.; Id, T.C.; Hotter, G.; Adam, R.; et al. The Relevance of the UPS in Fatty Liver Graft Preservation: A New Approach for IGL-1 and HTK Solutions. Int. J. Mol. Sci. 2017, 18, 2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaouali, M.A.; Ben Abdennebi, H.; Padrissa-Altés, S.; Alfany-Fernandez, I.; Rimola, A.; Roselló-Catafau, J. How Institut Georges Lopez Preservation Solution Protects Nonsteatotic and Steatotic Livers against Ischemia-Reperfusion Injury. Transplant Proc. 2011, 43, 77–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.Y.; Wang, J.Y.; Kausik, R.; Lee, K.Y.C.; Han, S. Nature of Interactions between PEO-PPO-PEO Triblock Copolymers and Lipid Membranes: (II) Role of Hydration Dynamics Revealed by Dynamic Nuclear Polarization. Biomacromolecules 2012, 13, 2624–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutheil, D.; Rioja-Pastor, I.; Tallineau, C.; Goujon, J.-M.; Hauet, T.; Mauco, G.; Petit-Paris, I. Protective Effect of PEG 35 000 Da on Renal Cells: Paradoxical Activation of JNK Signaling Pathway During Cold Storage. Am. J. Transplant. 2006, 6, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Bardallo, R.G.; da Silva, R.T.; Carbonell, T.; Folch-Puy, E.; Palmeira, C.; Roselló-Catafau, J.; Pirenne, J.; Adam, R.; Panisello-Roselló, A. Role of PEG35, Mitochondrial ALDH2, and Glutathione in Cold Fatty Liver Graft Preservation: An IGL-2 Approach. Int. J. Mol. Sci. 2021, 22, 5332. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, J.; Otarashvili, G.; Meszaros, A.; Ebner, S.; Weissenbacher, A.; Cardini, B.; Oberhuber, R.; Resch, T.; Öfner, D.; Schneeberger, S.; et al. Restoring Mitochondrial Function While Avoiding Redox Stress: The Key to Preventing Ischemia/Reperfusion Injury in Machine Perfused Liver Grafts? Int. J. Mol. Sci. 2020, 21, 3132. [Google Scholar] [CrossRef]
- Selzner, M.; RüDiger, H.A.; Sindram, D.; Madden, J.; Clavien, P.A. Mechanisms of Ischemic Injury Are Different in the Steatotic and Normal Rat Liver. Hepatology 2000, 32, 1280–1288. [Google Scholar] [CrossRef]
- Serafín, A.; Roselló-Catafau, J.; Prats, N.; Xaus, C.; Gelpí, E.; Peralta, C. Ischemic Preconditioning Increases the Tolerance of Fatty Liver to Hepatic Ischemia-Reperfusion Injury in the Rat. Am. J. Pathol. 2002, 161, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Witko-Sarsat, V.; Friedlander, M.; Nguyen Khoa, T.; Capeillère-Blandin, C.; Nguyen, A.T.; Canteloup, S.; Dayer, J.M.; Jungers, P.; Drüeke, T.; Descamps-Latscha, B. Advanced Oxidation Protein Products as Novel Mediators of Inflammation and Monocyte Activation in Chronic Renal Failure. J. Immunol. 1998, 161, 2524–2532. [Google Scholar]
- Nakamura, A.; Goto, S. Analysis of Protein Carbonyls with 2,4-Dinitrophenyl Hydrazine and Its Antibodies by Immunoblot in Two-Dimensional Gel Electrophoresis. J. Biochem. 1996, 119, 768–774. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Hissin, P.J.; Hilf, R. A Fluorometric Method for Determination of Oxidized and Reduced Glutathione in Tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef]
- Coimbra-Costa, D.; Alva, N.; Duran, M.; Carbonell, T.; Rama, R. Oxidative Stress and Apoptosis after Acute Respiratory Hypoxia and Reoxygenation in Rat Brain. Redox Biol. 2017, 12, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Kirshner, Z.Z.; Gibbs, R.B. Use of the REVERT® Total Protein Stain as a Loading Control Demonstrates Significant Benefits over the Use of Housekeeping Proteins When Analyzing Brain Homogenates by Western Blot: An Analysis of Samples Representing Different Gonadal Hormone States. Mol. Cell. Endocrinol. 2018, 473, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Forte, M.; Bianchi, F.; Cotugno, M.; Marchitti, S.; Stanzione, R.; Maglione, V.; Sciarretta, S.; Valenti, V.; Carnevale, R.; Versaci, F.; et al. An Interplay between UCP2 and ROS Protects Cells from High-Salt-Induced Injury through Autophagy Stimulation. Cell Death Dis. 2021, 12, 919. [Google Scholar] [CrossRef]
- Wang, H.; Ni, H.M.; Chao, X.; Ma, X.; Rodriguez, Y.A.; Chavan, H.; Wang, S.; Krishnamurthy, P.; Dobrowsky, R.; Xu, D.X.; et al. Double Deletion of PINK1 and Parkin Impairs Hepatic Mitophagy and Exacerbates Acetaminophen-Induced Liver Injury in Mice. Redox Biol. 2019, 22, 101148. [Google Scholar] [CrossRef]
- Pan, C.W.; Pan, Z.Z.; Hu, J.J.; Chen, W.L.; Zhou, G.Y.; Lin, W.; Jin, L.X.; Xu, C.L. Mangiferin Alleviates Lipopolysaccharide and D-Galactosamine-Induced Acute Liver Injury by Activating the Nrf2 Pathway and Inhibiting NLRP3 Inflammasome Activation. Eur. J. Pharmacol. 2016, 770, 85–91. [Google Scholar] [CrossRef]
- Hennig, P.; Garstkiewicz, M.; Grossi, S.; Di Filippo, M.; French, L.E.; Beer, H.D. The Crosstalk between Nrf2 and Inflammasomes. Int. J. Mol. Sci. 2018, 19, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witko-Sarsat, V.; Gausson, V.; Nguyen, A.T.; Touam, M.; Drüeke, T.; Santangelo, F.; Descamps-Latscha, B. AOPP-Induced Activation of Human Neutrophil and Monocyte Oxidative Metabolism: A Potential Target for N-Acetylcysteine Treatment in Dialysis Patients. Kidney Int. 2003, 64, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Caraceni, P.; Bianchi, C.; Domenicali, M.; Maria Pertosa, A.; Maiolini, E.; Parenti Castelli, G.; Nardo, B.; Trevisani, F.; Lenaz, G.; Bernardi, M. Impairment of Mitochondrial Oxidative Phosphorylation in Rat Fatty Liver Exposed to Preservation-Reperfusion Injury. J. Hepatol. 2004, 41, 82–88. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijević, D.; Aksentijević, A.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; et al. Ischaemic Accumulation of Succinate Controls Reperfusion Injury through Mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, D.; He, Z.; Fedorova, J.; Zhang, J.; Wood, E.; Zhang, X.; Kang, D.E.; Li, J. Sestrin2 Maintains OXPHOS Integrity to Modulate Cardiac Substrate Metabolism during Ischemia and Reperfusion. Redox Biol. 2021, 38, 101824. [Google Scholar] [CrossRef] [PubMed]
- Boss, O.; Hagen, T.; Lowell, B.B. Uncoupling Proteins 2 and 3. Perspect. Diabetes 2000, 49, 143–156. [Google Scholar]
- López-Bernardo, E.; Anedda, A.; Sánchez-Pérez, P.; Acosta-Iborra, B.; Cadenas, S. 4-Hydroxynonenal Induces Nrf2-Mediated UCP3 Upregulation in Mouse Cardiomyocytes. Free Radic. Biol. Med. 2015, 88, 427–438. [Google Scholar] [CrossRef]
- Ayyasamy, V.; Owens, K.M.; Desouki, M.M.; Liang, P.; Bakin, A.; Thangaraj, K.; Buchsbaum, D.J.; LoBuglio, A.F.; Singh, K.K. Cellular Model of Warburg Effect Identifies Tumor Promoting Function of UCP2 in Breast Cancer and Its Suppression by Genipin. PLoS ONE 2011, 6, e24792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echtay, K.S.; Esteves, T.C.; Pakay, J.L.; Jekabsons, M.B.; Lambert, A.J.; Otín, M.P.; Pamplona, R.; Puig, A.J.V.; Wang, S.; Roebuck, S.J.; et al. A Signalling Role for 4-Hydroxy-2-Nonenal in Regulation of Mitochondrial Uncoupling. EMBO J. 2003, 22, 4103–4110. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC175801 (accessed on 30 May 2020). [CrossRef] [Green Version]
- Wu, H.; Ye, M.; Liu, D.I.; Yang, J.; Ding, J.W.; Zhang, J.; Wang, X.; Dong, W.; Fan, Z.; Yang, J. UCP2 Protect the Heart from Myocardial Ischemia/Reperfusion Injury via Induction of Mitochondrial Autophagy. J. Cell. Biochem. 2019, 120, 15455–15466. [Google Scholar] [CrossRef]
- Ke, B.; Shen, X.-D.; Zhang, Y.; Ji, H.; Gao, F.; Yue, S.; Kamo, N.; Zhai, Y.; Yamamoto, M.; Busuttil, R.W.; et al. KEAP1-NRF2 Complex in Ischemia-Induced Hepatocellular Damage of Mouse Liver Transplants. J. Hepatol. 2013, 59, 1200–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudoh, K.; Uchinami, H.; Yoshioka, M.; Seki, E.; Yamamoto, Y. Nrf2 Activation Protects the Liver from Ischemia/Reperfusion Injury in Mice. Ann. Surg. 2014, 260, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhu, Z.; Xue, M.; Pan, X.; Tong, G.; Yi, X.; Fan, J.; Li, Y.; Li, W.; Dong, Y.; et al. The Protective Effects of Fibroblast Growth Factor 10 against Hepatic Ischemia-Reperfusion Injury in Mice. Redox Biol. 2021, 40, 101859. [Google Scholar] [CrossRef]
- Ehren, J.L.; Maher, P. Concurrent Regulation of the Transcription Factors Nrf2 and ATF4 Mediates the Enhancement of Glutathione Levels by the Flavonoid Fisetin. Biochem. Pharmacol. 2013, 85, 1816–1826. [Google Scholar] [CrossRef]
- Murata, H.; Takamatsu, H.; Liu, S.; Kataoka, K.; Huh, N.H.; Sakaguchi, M. NRF2 Regulates PINK1 Expression under Oxidative Stress Conditions. PLoS ONE 2015, 10, e0142438. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.; Xu, M.; Zhou, F.; Wein, A.N.; Upadhya, G.A.; Ye, L.; Wong, B.W.; Lin, Y.; O’Farrelly, C.; Chapman, W.C. NRF2 Assessment in Discarded Liver Allografts: A Role in Allograft Function and Salvage. Am. J. Transplant. 2022, 22, 58–70. [Google Scholar] [CrossRef]
- Arduini, A.; Mezzetti, A.; Porreca, E.; Lapenna, D.; DeJulia, J.; Marzio, L.; Polidoro, G.; Cuccurullo, F. Effect of Ischemia and Reperfusion on Antioxidant Enzymes and Mitochondrial Inner Membrane Proteins in Perfused Rat Heart. Biochim. Biophys. Acta Mol. Cell Res. 1988, 970, 113–121. [Google Scholar] [CrossRef]
- Dhalla, N.S.; Elmoselhi, A.B.; Hata, T.; Makino, N. Status of Myocardial Antioxidants in Ischemia-Reperfusion Injury. Cardiovasc. Res. 2000, 47, 446–456. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bardallo, R.G.; Company-Marin, I.; Folch-Puy, E.; Roselló-Catafau, J.; Panisello-Rosello, A.; Carbonell, T. PEG35 and Glutathione Improve Mitochondrial Function and Reduce Oxidative Stress in Cold Fatty Liver Graft Preservation. Antioxidants 2022, 11, 158. https://doi.org/10.3390/antiox11010158
Bardallo RG, Company-Marin I, Folch-Puy E, Roselló-Catafau J, Panisello-Rosello A, Carbonell T. PEG35 and Glutathione Improve Mitochondrial Function and Reduce Oxidative Stress in Cold Fatty Liver Graft Preservation. Antioxidants. 2022; 11(1):158. https://doi.org/10.3390/antiox11010158
Chicago/Turabian StyleBardallo, Raquel G., Idoia Company-Marin, Emma Folch-Puy, Joan Roselló-Catafau, Arnau Panisello-Rosello, and Teresa Carbonell. 2022. "PEG35 and Glutathione Improve Mitochondrial Function and Reduce Oxidative Stress in Cold Fatty Liver Graft Preservation" Antioxidants 11, no. 1: 158. https://doi.org/10.3390/antiox11010158
APA StyleBardallo, R. G., Company-Marin, I., Folch-Puy, E., Roselló-Catafau, J., Panisello-Rosello, A., & Carbonell, T. (2022). PEG35 and Glutathione Improve Mitochondrial Function and Reduce Oxidative Stress in Cold Fatty Liver Graft Preservation. Antioxidants, 11(1), 158. https://doi.org/10.3390/antiox11010158