

New In Vivo Approach to Broaden the Thioredoxin Family Interactome in Chloroplasts
 , , , ,
, , , ,
Abstract
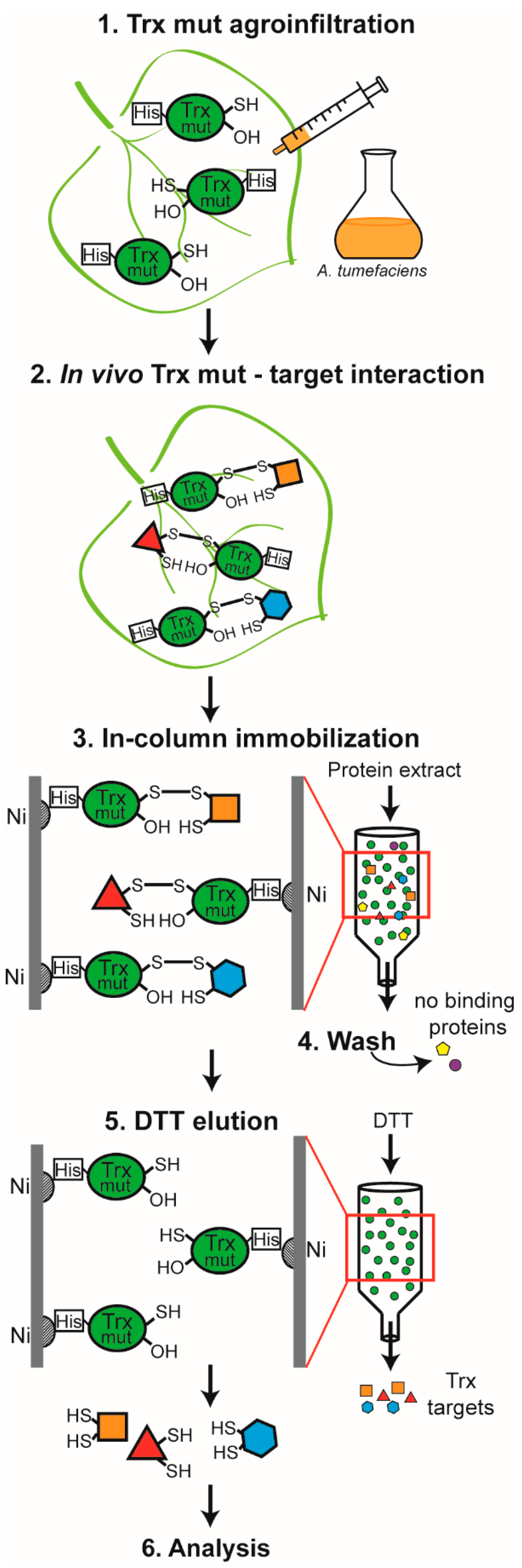
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Construction of Monocysteinic Trx Mutants and Plant Transformation
2.3. Protein Preparation and Purification of Target Proteins
2.4. Proteomic Analysis
2.4.1. Sample Preparation
2.4.2. Mass Spectrometry Analysis
2.4.3. Data Analysis
2.4.4. Peptide Quantification
2.4.5. Bioinformatics and Annotations
3. Results
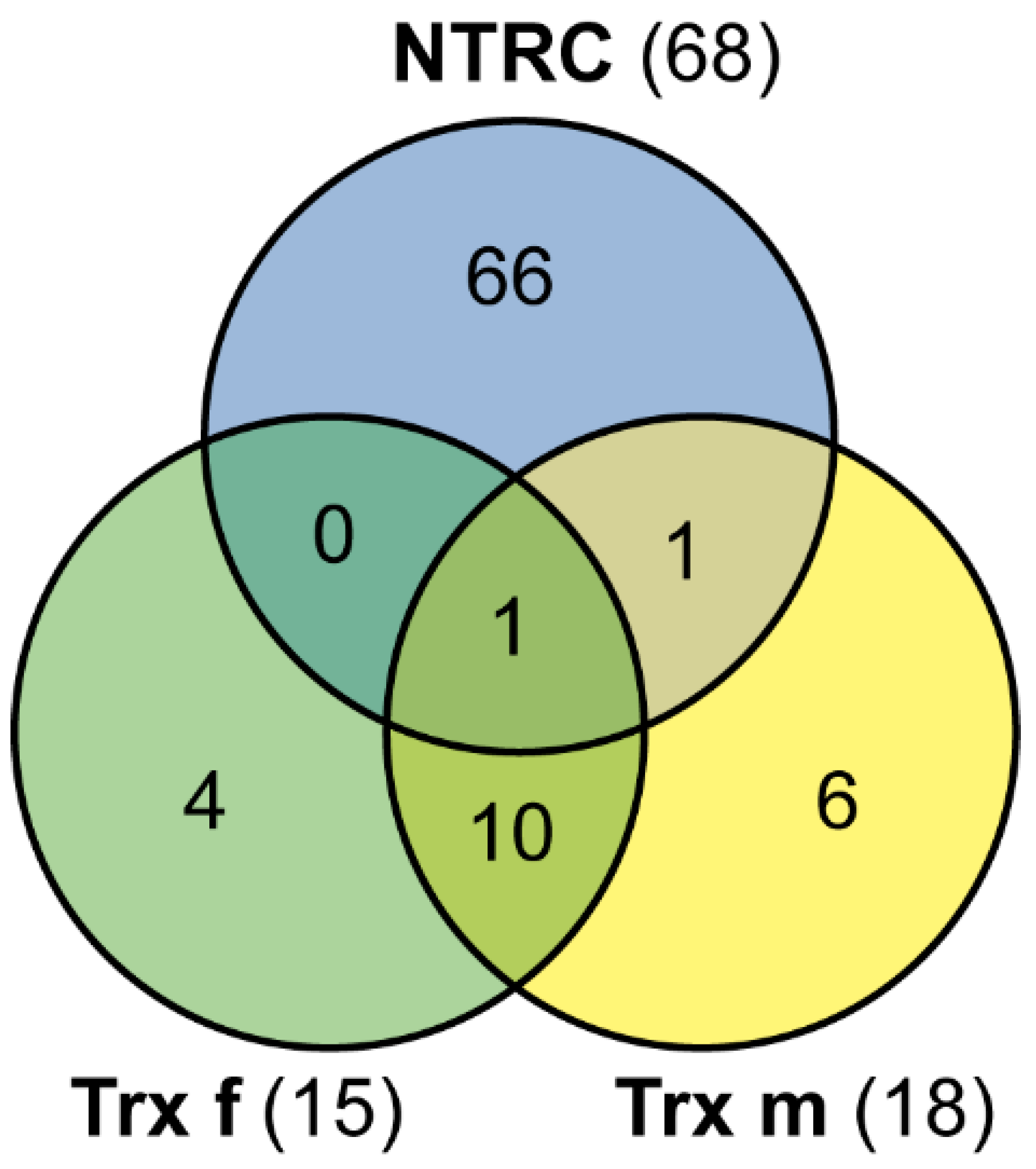
3.1. Identification of Proteins Captured as Redox Trx Interactors
3.2. Analysis of Candidate Trx Partners
4. Discussion
4.1. Novelty and Specificity of the Approach
4.2. New Potential Trx f, Trx m and NTRC Target Proteins
4.2.1. Trx f and Trx m
4.2.2. NTRC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buchanan, B.B. Role of Light in the Regulation of Chloroplast Enzymes. Annu. Rev. Plant Physiol. 1980, 31, 341–374. [Google Scholar] [CrossRef]
- Serrato, A.J.; Fernández-Trijueque, J.; Barajas-López, J.-D.-D.; Chueca, A.; Sahrawy, M. Plastid Thioredoxins: A “One-for-All” Redox-Signaling System in Plants. Front. Plant Sci. 2013, 4, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, B.B.; Schürmann, P.; Wolosiuk, R.A.; Jacquot, J.P. The Ferredoxin/Thioredoxin System: From Discovery to Molecular Structures and Beyond. Photosynth. Res. 2002, 73, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.; Qin, T.; Zhao, Z. Thioredoxins and Thioredoxin Reductase in Chloroplasts: A Review. Gene 2019, 706, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Serrato, A.J.; Pérez-Ruiz, J.M.; Spínola, M.C.; Cejudo, F.J. A Novel NADPH Thioredoxin Reductase, Localised in the Chloroplast, Which Deficiency Causes Hypersensitivity to Abiotic Stress in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 43821–43827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Hisabori, T. Two Distinct Redox Cascades Cooperatively Regulate Chloroplast Functions and Sustain Plant Viability. Proc. Natl. Acad. Sci. USA 2016, 113, E3967–E3976. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.Y.; Ehrlich, M.; Thormählen, I.; Lehmann, M.; Krahnert, I.; Obata, T.; Cejudo, F.J.; Fernie, A.R.; Geigenberger, P. NTRC Plays a Crucial Role in Starch Metabolism, Redox Balance, and Tomato Fruit Growth. Plant Physiol. 2019, 181, 976–992. [Google Scholar] [CrossRef] [Green Version]
- Montrichard, F.; Alkhalfioui, F.; Yano, H.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin Targets in Plants: The First 30 Years. J. Proteom. 2009, 72, 452–474. [Google Scholar] [CrossRef]
- Hägglund, P.; Bunkenborg, J.; Maeda, K.; Svensson, B. Identification of Thioredoxin Disulfide Targets Using a Quantitative Proteomics Approach Based on Isotope-Coded Affinity Tags. J. Proteome Res. 2008, 7, 5270–5276. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, M.; Zhu, N.; Strul, J.M.; Dufresne, C.P.; Schneider, J.D.; Harmon, A.C.; Chen, S. Identification of Thioredoxin Targets in Guard Cell Enriched Epidermal Peels Using CysTMT Proteomics. J. Proteom. 2016, 133, 48–53. [Google Scholar] [CrossRef]
- Kallis, G.B.; Holmgren, A. Differential Reactivity of the Functional Sulfhydryl Groups of Cysteine-32 and Cysteine-32 Present in the Reduced Form of Thioredoxin from Escherichia coli. J. Biol. Chem. 1980, 255, 10261–10265. [Google Scholar] [CrossRef]
- Verdoucq, L.; Vignols, F.; Jacquot, J.P.; Chartier, Y.; Meyer, Y. In Vivo Characterization of a Thioredoxin h Target Protein Defines a New Peroxiredoxin Family. J. Biol. Chem. 1999, 274, 19714–19722. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Noguchi, K.; Motohashi, K.; Hisabori, T. Systematic Exploration of Thioredoxin Target Proteins in Plant Mitochondria. Plant Cell Physiol. 2013, 54, 875–892. [Google Scholar] [CrossRef] [Green Version]
- Balmer, Y.; Koller, A.; del Val, G.; Manieri, W.; Schürmann, P.; Buchanan, B.B. Proteomics Gives Insight into the Regulatory Function of Chloroplast Thioredoxins. Proc. Natl. Acad. Sci. USA 2003, 100, 370–375. [Google Scholar] [CrossRef] [Green Version]
- Marchand, C.; Le Maréchal, P.; Meyer, Y.; Decottignies, P. Comparative Proteomic Approaches for the Isolation of Proteins Interacting with Thioredoxin. Proteomics 2006, 6, 6528–6537. [Google Scholar] [CrossRef] [PubMed]
- Alkhalfioui, F.; Renard, M.; Vensel, W.H.; Wong, J.; Tanaka, C.K.; Hurkman, W.J.; Buchanan, B.B.; Montrichard, F. Thioredoxin-Linked Proteins Are Reduced during Germination of Medicago truncatula Seeds. Plant Physiol. 2007, 144, 1559–1579. [Google Scholar] [CrossRef] [Green Version]
- Rey, P.; Cuiné, S.; Eymery, F.; Garin, J.; Court, M.; Jacquot, J.P.; Rouhier, N.; Broin, M. Analysis of the Proteins Targeted by CDSP32, a Plastidic Thioredoxin Participating in Oxidative Stress Responses. Plant J. 2005, 41, 31–42. [Google Scholar] [CrossRef]
- González, M.; Delgado-Requerey, V.; Ferrández, J.; Serna, A.; Cejudo, F.J. Insights into the Function of NADPH Thioredoxin Reductase C (NTRC) Based on Identification of NTRC-Interacting Proteins in Vivo. J. Exp. Bot. 2019, 70, 5787–5798. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Barrio, R.; Fernández-San Millán, A.; Corral-Martínez, P.; Seguí-Simarro, J.M.; Farran, I. Tobacco Plastidial Thioredoxins as Modulators of Recombinant Protein Production in Transgenic Chloroplasts. Plant Biotechnol. J. 2011, 9, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Hennegan, K.P.; Danna, K.J. PBIN20: An Improved Binary Vector for Agrobacterium-Mediated Transformation. Plant Mol. Biol. Report. 1998, 16, 129–131. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Tang, S.; Zhao, Q.; Zhang, Z.; Zhang, H.; Dong, L.; Guo, H.; Xie, Q. An Efficient System to Detect Protein Ubiquitination by Agroinfiltration in Nicotiana benthamiana. Plant J. 2010, 61, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-Gel Digestion for Mass Spectrometric Characterization of Proteins and Proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Shilov, I.V.; Seymour, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The Paragon Algorithm, a Next Generation Search Engine That Uses Sequence Temperature Values and Feature Probabilities to Identify Peptides from Tandem Mass Spectra. Mol. Cell. Proteom. 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.H.; Shilov, I.V.; Seymour, S.L. Nonlinear Fitting Method for Determining Local False Discovery Rates from Decoy Database Searches. J. Proteome Res. 2008, 7, 3661–3667. [Google Scholar] [CrossRef] [PubMed]
- Brandes, H.K.; Larimer, F.W.; Geck, M.K.; Stringer, C.D.; Schürmann, P.; Hartman, F.C. Direct Identification of the Primary Nucleophile of Thioredoxin f. J. Biol. Chem. 1993, 268, 18411–18414. [Google Scholar] [CrossRef]
- Bolger, M.E.; Arsova, B.; Usadel, B. Plant Genome and Transcriptome Annotations: From Misconceptions to Simple Solutions. Brief. Bioinform. 2018, 19, 437–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schürmann, P.; Buchanan, B.B. The Ferredoxin/Thioredoxin System of Oxygenic Photosynthesis. Antioxid. Redox Signal. 2008, 10, 1235–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsian, J.; Lomonossoff, G.P. Molecular Pharming-VLPs Made in Plants. Curr. Opin. Biotechnol. 2016, 37, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berggård, T.; Linse, S.; James, P. Methods for the Detection and Analysis of Protein-Protein Interactions. Proteomics 2007, 7, 2833–2842. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, K.; Kondoh, A.; Stumpp, M.T.; Hisabori, T. Comprehensive Survey of Proteins Targeted by Chloroplast Thioredoxin. Proc. Natl. Acad. Sci. USA 2001, 98, 11224–11229. [Google Scholar] [CrossRef]
- Motohashi, K.; Hisabori, T. HCF164 Receives Reducing Equivalents from Stromal Thioredoxin across the Thylakoid Membrane and Mediates Reduction of Target Proteins in the Thylakoid Lumen. J. Biol. Chem. 2006, 281, 35039–35047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouhier, N.; Villarejo, A.; Srivastava, M.; Gelhaye, E.; Keech, O.; Droux, M.; Finkemeier, I.; Samuelsson, G.; Dietz, K.J.; Jacquot, J.-P.; et al. Identification of Plant Glutaredoxin Targets. Antioxid. Redox Signal. 2005, 7, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Nikkanen, L.; Toivola, J.; Rintamäki, E. Crosstalk between Chloroplast Thioredoxin Systems in Regulation of Photosynthesis. Plant Cell Environ. 2016, 39, 1691–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojeda, V.; Pérez-Ruiz, J.M.; Cejudo, F.J. The NADPH-Dependent Thioredoxin Reductase C-2-Cys Peroxiredoxin Redox System Modulates the Activity of Thioredoxin x in Arabidopsis Chloroplasts. Plant Cell Physiol. 2018, 59, 2155–2164. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ruiz, J.M.; Naranjo, B.; Ojeda, V.; Guinea, M.; Cejudo, F.J. NTRC-Dependent Redox Balance of 2-Cys Peroxiredoxins Is Needed for Optimal Function of the Photosynthetic Apparatus. Proc. Natl. Acad. Sci. USA 2017, 114, 12069–12074. [Google Scholar] [CrossRef] [Green Version]
- Chibani, K.; Tarrago, L.; Gualberto, J.M.; Wingsle, G.; Rey, P.; Jacquot, J.P.; Rouhier, N. Atypical Thioredoxins in Poplar: The Glutathione-Dependent Thioredoxin-like 2.1 Supports the Activity of Target Enzymes Possessing a Single Redox Active Cysteine. Plant Physiol. 2012, 159, 592–605. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, K.; Romano, P.G.N.; Hisabori, T. Identification of Thioredoxin Targeted Proteins Using Thioredoxin Single-Cysteine Mutant-Immobilized Resin. Methods Mol. Biol. 2009, 479, 117–131. [Google Scholar]
- Bohrer, A.-S.; Massot, V.; Innocenti, G.; Reichheld, J.-P.; Issakidis-Bourguet, E.; Vanacker, H. New Insights into the Reduction Systems of Plastidial Thioredoxins Point out the Unique Properties of Thioredoxin z from Arabidopsis. J. Exp. Bot. 2012, 63, 6315–6323. [Google Scholar] [CrossRef] [Green Version]
- Collin, V.; Issakidis-Bourguet, E.; Marchand, C.; Hirasawa, M.; Lancelin, J.-M.; Knaff, D.B.; Miginiac-Maslow, M. The Arabidopsis Plastidial Thioredoxins. J. Biol. Chem. 2003, 278, 23747–23752. [Google Scholar] [CrossRef] [Green Version]
- Collin, V.; Lamkemeyer, P.; Miginiac-Maslow, M.; Hirasawa, M.; Knaff, D.B.; Dietz, K.-J.; Issakidis-Bourguet, E. Characterization of Plastidial Thioredoxins from Arabidopsis Belonging to the New Y-Type. Plant Physiol. 2004, 136, 4088–4095. [Google Scholar] [CrossRef] [Green Version]
- Gama, F.; Bréhélin, C.; Gelhaye, E.; Meyer, Y.; Jacquot, J.P.; Rey, P.; Rouhier, N. Functional Analysis and Expression Characteristics of Chloroplastic Prx IIE. Physiol. Plant. 2008, 133, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Marchand, C.; Le Maréchal, P.; Meyer, Y.; Miginiac-Maslow, M.; Issakidis-Bourguet, E.; Decottignies, P. New Targets of Arabidopsis Thioredoxins Revealed by Proteomic Analysis. Proteomics 2004, 4, 2696–2706. [Google Scholar] [CrossRef] [PubMed]
- Rey, P.; Sanz-Barrio, R.; Innocenti, G.; Ksas, B.; Courteille, A.; Rumeau, D.; Issakidis-Bourguet, E.; Farran, I. Overexpression of Plastidial Thioredoxins f and m Differentially Alters Photosynthetic Activity and Response to Oxidative Stress in Tobacco Plants. Front. Plant Sci. 2013, 4, 390. [Google Scholar] [CrossRef] [Green Version]
- Chibani, K.; Tarrago, L.; Schürmann, P.; Jacquot, J.P.; Rouhier, N. Biochemical Properties of Poplar Thioredoxin z. FEBS Lett. 2011, 585, 1077–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navrot, N.; Collin, V.; Gualberto, J.; Gelhaye, E.; Hirasawa, M.; Rey, P.; Knaff, D.B.; Issakidis, E.; Jacquot, J.P.; Rouhier, N. Plant Glutathione Peroxidases Are Functional Peroxiredoxins Distributed in Several Subcellular Compartments and Regulated during Biotic and Abiotic Stresses. Plant Physiol. 2006, 142, 1364–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertle, A.P.; Blunder, T.; Wunder, T.; Pesaresi, P.; Pribil, M.; Armbruster, U.; Leister, D. PGRL1 Is the Elusive Ferredoxin-Plastoquinone Reductase in Photosynthetic Cyclic Electron Flow. Mol. Cell 2013, 49, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Courteille, A.; Vesa, S.; Sanz-Barrio, R.; Cazale, A.-C.; Becuwe-Linka, N.; Farran, I.; Havaux, M.; Rey, P.; Rumeau, D. Thioredoxin M4 Controls Photosynthetic Alternative Electron Pathways in Arabidopsis. Plant Physiol. 2013, 161, 508–520. [Google Scholar] [CrossRef] [Green Version]
- Glaring, M.A.; Skryhan, K.; Kötting, O.; Zeeman, S.C.; Blennow, A. Comprehensive Survey of Redox Sensitive Starch Metabolising Enzymes in Arabidopsis thaliana. Plant Physiol. Biochem. 2012, 58, 89–97. [Google Scholar] [CrossRef]
- Skryhan, K.; Cuesta-Seijo, J.A.; Nielsen, M.M.; Marri, L.; Mellor, S.B.; Glaring, M.A.; Jensen, P.E.; Palcic, M.M.; Blennow, A. The Role of Cysteine Residues in Redox Regulation and Protein Stability of Arabidopsis thaliana Starch Synthase 1. PLoS ONE 2015, 10, e0136997. [Google Scholar] [CrossRef]
- Yoshida, K.; Hara, S.; Hisabori, T. Thioredoxin Selectivity for Thiol-Based Redox Regulation of Target Proteins in Chloroplasts. J. Biol. Chem. 2015, 290, 14278–14288. [Google Scholar] [CrossRef] [Green Version]
- Okegawa, Y.; Motohashi, K. Chloroplastic Thioredoxin m Functions as a Major Regulator of Calvin Cycle Enzymes during Photosynthesis in Vivo. Plant J. 2015, 84, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Thormählen, I.; Zupok, A.; Rescher, J.; Leger, J.; Weissenberger, S.; Groysman, J.; Orwat, A.; Chatel-Innocenti, G.; Issakidis-Bourguet, E.; Armbruster, U.; et al. Thioredoxins Play a Crucial Role in Dynamic Acclimation of Photosynthesis in Fluctuating Light. Mol. Plant 2017, 10, 168–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Portis, A.R. Mechanism of Light Regulation of Rubisco: A Specific Role for the Larger Rubisco Activase Isoform Involving Reductive Activation by Thioredoxin-f. Proc. Natl. Acad. Sci. USA 1999, 96, 9438–9443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trebitsh, T.; Levitan, A.; Sofer, A.; Danon, A. Translation of Chloroplast PsbA MRNA Is Modulated in the Light by Counteracting Oxidizing and Reducing Activities. Mol. Cell. Biol. 2000, 20, 1116–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baginsky, S.; Gruissem, W. Endonucleolytic Activation Directs Dark-Induced Chloroplast MRNA Degradation. Nucleic Acids Res. 2002, 30, 4527–4533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Calcagno, P.E.; Howard, T.P.; Raines, C.A. The CP12 Protein Family: A Thioredoxin-Mediated Metabolic Switch? Front. Plant Sci. 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bick, J.A.; Setterdahl, A.; Knaff, D.B.; Chen, Y.; Pitcher, L.H.; Zilinskas, B.; Leustek, T. Regulation of the Plant-Type 5′-Adenylyl Sulfate Reductase by Oxidative Stress. Biochemistry 2001, 40, 9040–9048. [Google Scholar] [CrossRef] [PubMed]
- Pulido, P.; Spínola, M.C.; Kirchsteiger, K.; Guinea, M.; Pascual, M.B.; Sahrawy, M.; Sandalio, L.M.; Dietz, K.-J.; González, M.; Cejudo, F.J. Functional Analysis of the Pathways for 2-Cys Peroxiredoxin Reduction in Arabidopsis thaliana Chloroplasts. J. Exp. Bot. 2010, 61, 4043–4054. [Google Scholar] [CrossRef] [PubMed]
- Yoo, K.S.; Ok, S.H.; Jeong, B.-C.; Jung, K.W.; Cui, M.H.; Hyoung, S.; Lee, M.-R.; Song, H.K.; Shin, J.S. Single Cystathionine β-Synthase Domain-Containing Proteins Modulate Development by Regulating the Thioredoxin System in Arabidopsis. Plant Cell 2011, 23, 3577–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Lee, J.; Kim, Y.; Bae, D.; Kang, K.Y.; Yoon, S.C.; Lim, D. Defining the Plant Disulfide Proteome. Electrophoresis 2004, 25, 532–541. [Google Scholar] [CrossRef]
- Balmer, Y.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin Target Proteins in Chloroplast Thylakoid Membranes. Antioxid. Redox Signal. 2006, 8, 1829–1834. [Google Scholar] [CrossRef] [PubMed]
- Nikkanen, L.; Toivola, J.; Trotta, A.; Diaz, M.G.; Tikkanen, M.; Aro, E.-M.; Rintamaki, E. Regulation of Cyclic Electron Flow by Chloroplast NADPH-Dependent Thioredoxin System. Plant Direct 2018, 2, e00093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Liu, J.; Liu, B.; Feng, D.; Da, Q.; Wang, P.; Shu, S.; Su, J.; Zhang, Y.; Wang, J.; et al. Evidence for a Role of Chloroplastic M-Type Thioredoxins in the Biogenesis of Photosystem II in Arabidopsis. Plant Physiol. 2013, 163, 1710–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalska, J.; Zauber, H.; Buchanan, B.B.; Cejudo, F.J.; Geigenberger, P. NTRC Links Built-in Thioredoxin to Light and Sucrose in Regulating Starch Synthesis in Chloroplasts and Amyloplasts. Proc. Natl. Acad. Sci. USA 2009, 106, 9908–9913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ancín, M.; Larraya, L.; Millán, A.F.-S.; Veramendi, J.; Burch-Smith, T.; Farran, I. NTRC and Thioredoxin f Overexpression Differentially Induces Starch Accumulation in Tobacco Leaves. Plants 2019, 8, 543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepistö, A.; Pakula, E.; Toivola, J.; Krieger-Liszkay, A.; Vignols, F.; Rintamäki, E. Deletion of Chloroplast NADPH-Dependent Thioredoxin Reductase Results in Inability to Regulate Starch Synthesis and Causes Stunted Growth under Short-Day Photoperiods. J. Exp. Bot. 2013, 64, 3843–3854. [Google Scholar] [CrossRef]
- Yamazaki, D.; Motohashi, K.; Kasama, T.; Hara, Y.; Hisabori, T. Target Proteins of the Cytosolic Thioredoxins in Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.H.; Cai, N.; Balmer, Y.; Tanaka, C.K.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin Targets of Developing Wheat Seeds Identified by Complementary Proteomic Approaches. Phytochemistry 2004, 65, 1629–1640. [Google Scholar] [CrossRef] [PubMed]
- Alkhalfioui, F.; Renard, M.; Montrichard, F. Unique Properties of NADP-Thioredoxin Reductase C in Legumes. J. Exp. Bot. 2007, 58, 969–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichter, A.; Häberlein, I. A Light-Dependent Redox Signal Participates in the Regulation of Ammonia Fixation in Chloroplasts of Higher Plants—Ferredoxin: Glutamate Synthase Is a Thioredoxin-Dependent Enzyme. J. Plant Physiol. 1998, 153, 83–90. [Google Scholar] [CrossRef]
- Balmer, Y.; Vensel, W.H.; Cai, N.; Manieri, W.; Schürmann, P.; Hurkman, W.J.; Buchanan, B.B. A Complete Ferredoxin/Thioredoxin System Regulates Fundamental Processes in Amyloplasts. Proc. Natl. Acad. Sci. USA 2006, 103, 2988–2993. [Google Scholar] [CrossRef] [PubMed]
- Rolland, N.; Ruffet, M.L.; Job, D.; Douce, R.; Droux, M. Spinach Chloroplast O-Acetylserine (Thiol)-Lyase Exhibits Two Catalytically Non-Equivalent Pyridoxal-5′-Phosphate-Containing Active Sites. Eur. J. Biochem. 1996, 236, 272–282. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nicotiana tabacum | Arabidopsis thaliana | Trx f | Trx m | Cys | Subcellular Localisation | |||
|---|---|---|---|---|---|---|---|---|
| UniProtKB | Protein Name | UniProtKB | Protein Name | Identity % | ||||
| Cell antioxidant and redox homeostasis | ||||||||
| A0A1S4A3V7 | 2-Cys peroxiredoxin BAS1 | Q9C5R8 | 2-Cys peroxiredoxin BAS1-like(2-Cys Prx B) | 77 | x | x | stroma | |
| A0A1S4A969 | Glutathione peroxidase | P52032 | Phospholipid hydroperoxide glutathione peroxidase 1 (PHGPx) | 72.4 | x | stroma | ||
| A0A1S4C620 | Peptide methionine sulfoxide reductase-like | P54150 | Peptide methionine sulfoxide reductase A4 (MSRA4) | 62 | x | x | stroma | |
| A0A1S4B900 | Peroxiredoxin Q | Q9LU86 | Peroxiredoxin Q (Prx Q) | 76.3 | x | x | lumen | |
| A0A1S4D678 | Peroxiredoxin-2E-2 | Q949U7 | Peroxiredoxin-2E (Prx IIE) | 57.7 | x | stroma | ||
| A0A1S4ASD9 | Thioredoxin-like 2 | Q8LCT3-2 | Thioredoxin-like 2-2 (Lilium 2) | 65.5 | x | 2 | stroma | |
| A0A1S4D2Y9 | Thioredoxin-like 4 | Q9C5C5 | Thioredoxin-like 4 (Lilium 5) | 66.3 | x | x | 2 | stroma |
| Photosynthesis | ||||||||
| A0A1S4CIH3 | PGR5-like protein 1A | Q8H112 | PGR5-like protein 1A (PGRL1) | 72.8 | x | x | thylakoid m | |
| Carbon metabolism | ||||||||
| A0A1S4ATB8 | Glyceraldehyde-3-phosphate dehydrogenase | P25857 | Glyceraldehyde-3-phosphate dehydrogenase GAPB (NADP-GAPDH) | 85.8 | x | stroma | ||
| A0A1S3YQS9 | Malate dehydrogenase (NADP) | Q8H1E2 | Malate dehydrogenase (NADP-MDH) | 79.4 | x | stroma | ||
| A0A1S4CZ71 | Phosphoglucan water dikinase | Q6ZY51 | Phosphoglucan water dikinase (PWD) | 61.2 | x | x | 7 | stroma |
| A0A1S4A3L9 | Ribulose bisphosphate carboxylase/oxygenase activase 1 | P10896 | Ribulose bisphosphate carboxylase/oxygenase activase (RuBisCO activase) | 78.6 | x | x | stroma | |
| A0A1S3YEX0 | Sedoheptulose-1,7-bisphosphatase | P46283 | Sedoheptulose-1,7-bisphosphatase (SBPase) | 81.5 | x | stroma | ||
| A0A1S4DEY3 | Starch synthase | Q9MAQ0 | Granule-bound starch synthase 1 (GBSS1) | 74.1 | x | 6 | stroma | |
| Protein folding | ||||||||
| A0A1S4DGW7 | Peptidylprolyl isomerase | Q9LYR5 | Peptidyl-prolyl cis-trans isomerase FKBP19 (PPIase FKBP19) | 87.7 | x | lumen | ||
| Transcription and translation regulation | ||||||||
| A0A1S4DL03 | 50S ribosomal protein L18 | Q9SX68 | 50S ribosomal protein L18 | 83.8 | x | 1 | stroma | |
| Q6T7F3 | Amidophosphoribosyltransferase | Q9STG9 | Amidophosphoribosyltransferase 2 (ATase2) | 77.6 | x | x | 9 | stroma |
| A0A140G1V5 | Ribosomal protein S3 | P56798 | 30S ribosomal protein S3 | 87.6 | x | x | 3 | stroma |
| A0A1S4CR05 | Uridine kinase | Q9FKS0 | Uridine kinase-like protein 1 (UK) | 87.4 | x | x | # | stroma |
| Secondary metabolism | ||||||||
| A0A1S4B8Q6 | Cis-abienol synthase | G3CCC1 | Cis-abienol synthase (ABS) * | x | x | # | stroma | |
| Amino acid biosynthesis | ||||||||
| A0A1S4DHJ8 | 5′-adenylylsulfate reductase 2 | P92981 | 5′-adenylylsulfate reductase 2 (APR2) | 75.4 | x | x | stroma | |
| Unknown function | ||||||||
| A0A1S4CQQ3 | Thylakoid lumenal 29 kDa protein | P82281 | Thylakoid lumenal 29 kDa protein (TL29) | 69.4 | x | lumen | ||
| Nicotiana tabacum | Arabidopsis thaliana | Cys | Subcellular Localisation | |||
|---|---|---|---|---|---|---|
| UniProtKB | Protein Name | UniProtKB | Protein Name | Identity % | ||
| Cell antioxidant and redox homeostasis | ||||||
| A0A1S4A3V7 | 2-Cys peroxiredoxin BAS1 | Q9C5R8 | 2-Cys peroxiredoxin BAS1-like (2-Cys Prx B) | 77 | stroma | |
| A0A1S4B1Q8 | CBS domain-containing protein CBSX1 | O23193 | CBS domain-containing protein CBSX1 (CDCP2) | 63.6 | stroma | |
| A0A1S3Y8V8 | Glutathione S-transferase DHAR3 | Q8LE52 | GSH-dependent dehydroascorbate reductase 3 (DHAR3) | 72.5 | stroma | |
| A0A1S3ZZS2 | Probable L-ascorbate peroxidase 6 | Q42593 | L-ascorbate peroxidase T (tAPX) | 73.7 | thylakoid m | |
| A0A1S4A969 | Glutathione peroxidase | P52032 | Phospholipid hydroperoxide glutathione peroxidase 1 (PHGPx) | 72.4 | stroma | |
| W0KRH1 | Superoxide dismutase | Q9LU64 | Superoxide dismutase [Fe] 2 (FSD2) | 59.1 | 0 | thylakoid m |
| A0A1S4CCB3 | Thioredoxin-like | Q9SEU6 | Thioredoxin M4 (Trx m4) | 53.3 | stroma | |
| Photosynthesis | ||||||
| A0A140G1P8 | ATP synthase CF0 B subunit | P56759 | ATP synthase subunit b | 88.6 | 1 | thylakoid m |
| A0A1S4CSA5 | ATP synthase delta chain | Q9SSS9 | ATP synthase subunit delta | 60.5 | thylakoid m | |
| P00823 | ATP synthase subunit alpha | P56757 | ATP synthase subunit alpha | 94 | thylakoid m | |
| A0A140G1S2 | ATP synthase subunit beta | P19366 | ATP synthase subunit beta | 93 | thylakoid m | |
| A0A1S4CBW5 | Chlorophyll a-b binding protein | Q9SY97 | Photosystem I chlorophyll a-b binding protein 3-1 (Lhca3.1) | 89.4 | 0 | thylakoid m |
| Q0PWS6 | Chlorophyll a-b binding protein | Q9C639 | Photosystem I chlorophyll a-b binding protein 5 (Lhca5) | 38 | 0 | thylakoid m |
| Q40512 | Chlorophyll a-b binding protein | Q01667 | Photosystem I chlorophyll a-b binding protein 6 (Lhca1) | 87.4 | 3 | thylakoid m |
| A0A1S4BMB0 | Chlorophyll a-b binding protein | Q9SHR7 | Photosystem II chlorophyll a-b binding protein 2.1 (Lhcb2.1) | 89 | thylakoid m | |
| A0A1S4DIE1 | Chlorophyll a-b binding protein | Q9S7M0 | Photosystem II chlorophyll a-b binding protein 3 (Lhcb3) | 87.5 | thylakoid m | |
| Q0PWS7 | Chlorophyll a-b binding protein | Q07473 | Photosystem II chlorophyll a-b binding protein CP29.1 (Lhcb4.1) | 86.6 | thylakoid m | |
| A0A140G1T3 | Cytochrome b559 subunit alpha | P56779 | Cytochrome b559 subunit alpha | 99 | 0 | thylakoid m |
| A0A1S3XVT6 | Cytochrome b6 | P56773 | Cytochrome b6 | 98 | 2 | thylakoid m |
| A0A1S4B832 | Cytochrome b6-f complex iron-sulfur subunit | Q9ZR03 | Cytochrome b6-f complex iron-sulfur subunit (RISP) | 77.8 | 4 | thylakoid m |
| A0A140G1S8 | Cytochrome f | P56771 | Cytochrome f | 90 | 2 | thylakoid m |
| A0A1S3YVN4 | Ferredoxin | P16972 | Ferredoxin-2 (Fd2) | 65.7 | 4 | stroma |
| A0A1S4B5N2 | Ferredoxin-thioredoxin reductase | A0A1P8BDN6 | Ferredoxin-thioredoxin reductase subunit A (Variable subunit) 2 | 46.5 | stroma | |
| Q84QE8 | Oxygen evolving complex 33 kDa photosystem II protein | Q9S841 | Oxygen-evolving enhancer protein 1-2 (OEE-1) | 81.3 | thylakoid ls | |
| A0A1S4BMY9 | Oxygen-evolving enhancer protein 2-2 | Q42029 | Oxygen-evolving enhancer protein 2-1 (OEE-2) | 72.9 | thylakoid ls | |
| A0A1S3XRM3 | Oxygen-evolving enhancer protein 3-2 | Q41932 | Oxygen-evolving enhancer protein 3-2 (OEE-3) | 68.8 | 0 | thylakoid ls |
| A0A140G1 × 0 | Photosystem I iron-sulfur center | P62090 | Photosystem I iron-sulfur center (PSI-C) | 100 | 9 | thylakoid m |
| A0A140G1R3 | Photosystem I P700 chlorophyll a apoprotein A1 | P56766 | Photosystem I P700 chlorophyll a apoprotein A1 (PSI-A) | 98 | 4 | thylakoid m |
| A0A140G1R2 | Photosystem I P700 chlorophyll a apoprotein A2 | P56767 | Photosystem I P700 chlorophyll a apoprotein A2 (PSI-B) | 98 | 2 | thylakoid m |
| A0A1S3ZIE1 | Photosystem I reaction center subunit II | Q9SA56 | Photosystem I reaction center subunit II-2 (PSI-D2) | 76.6 | 1 | thylakoid m |
| A0A1S4CFV4 | Photosystem I reaction center subunit IV A | Q9S831 | Photosystem I reaction center subunit IV A (PSI-E1) | 0 | thylakoid m | |
| A0A1S4CYN6 | Photosystem I reaction center subunit IV B | Q9S714 | Photosystem I reaction center subunit IV B (PSI-E2) | 58.4 | 0 | thylakoid m |
| D2K7Z2 | Photosystem I reaction center subunit | P49107 | Photosystem I reaction center subunit N (PSI-N) | 69.1 | thylakoid m | |
| A0A1S4CR54 | Photosystem I reaction center subunit VI-1 | Q9SUI7 | Photosystem I reaction center subunit VI-1 (PSI-H1) | 77.2 | 0 | thylakoid m |
| A0A1S4BQS3 | Photosystem I reaction center subunit XI | Q9SUI4 | Photosystem I reaction center subunit XI (PSI-L) | 80.7 | 1 | thylakoid m |
| A0A1S3YQ87 | Photosystem II 22 kDa protein | Q9XF91 | Photosystem II 22 kDa protein (CP22) | 73.2 | 0 | thylakoid m |
| A0A140G1Q8 | Photosystem II CP43 reaction center protein | P56778 | Photosystem II CP43 reaction center protein | 98 | 3 | thylakoid m |
| A0A140G1U3 | Photosystem II CP47 reaction center protein | P56777 | Photosystem II CP47 reaction center protein | 98.6 | thylakoid m | |
| A0A140G1Q7 | Photosystem II D2 protein | P56761 | Photosystem II D2 protein | 99 | thylakoid m | |
| A0A140G1P2 | Photosystem II D1 protein | P83755 | Photosystem II D1 protein | 99.7 | thylakoid m | |
| A0A1S4A1K3 | Plastocyanin | P42699 | Plastocyanin major isoform | 67.7 | 0 | thylakoid ls |
| Carbon metabolism | ||||||
| A0A1S4A023 | Fructose-1,6-bisphosphatase | P25851 | Fructose-1,6-bisphosphatase 1 (FBPase 1) | 86.5 | stroma | |
| A7XAQ5 | Glucose-1-phosphate adenylyltransferase | P55228 | Glucose-1-phosphate adenylyltransferase small subunit (AGPase B) | 87.5 | stroma | |
| A0A1S3Z1 × 1 | Probable ribose-5-phosphate isomerase 3 | Q9S726 | Probable ribose-5-phosphate isomerase 3 | 68.3 | stroma | |
| A0A140G1S3 | Ribulose bisphosphate carboxylase large chain | O03042 | Ribulose bisphosphate carboxylase large chain (RuBisCO LSU) | 94 | stroma | |
| A0A1S3X2Z0 | Triosephosphate isomerase | Q9SKP6 | Triosephosphate isomerase (TPI) | 79.4 | stroma | |
| Protein folding | ||||||
| A0A1S4AH01 | 10 kDa chaperonin-like | Q9M1C2 | 10 kDa chaperonin 1 (CPN10) | 71 | 2 | stroma |
| A0A077DBL2 | 20 kDa chaperonin | O65282 | 20 kDa chaperonin (CPN20) | 74 | 0 | stroma |
| A0A1S4AWT3 | Peptidyl-prolyl cis-trans isomerase | Q9ASS6 | Peptidyl-prolyl cis-trans isomerase CYP20-2 | 69.3 | 0 | thylakoid ls |
| A0A1S3ZH83 | Peptidyl-prolyl cis-trans isomerase CYP38 | Q9SSA5 | Peptidyl-prolyl cis-trans isomerase CYP38 | 76.8 | 0 | lumen |
| A0A1S3XJV2 | Peptidyl-prolyl isomerase | O22870 | Peptidyl-prolyl cis-trans isomerase FKBP16-3 | 65.5 | 0 | lumen |
| A0A1S4DIY1 | RuBisCO large subunit-binding protein subunit beta | P21240 | Chaperonin 60 subunit beta 1 (CPN60) | 81.8 | stroma | |
| Transcription and translation regulation | ||||||
| A0A1S4CYJ5 | 29 kDa ribonucleoprotein A | Q9ZUU4 | RNA-binding protein CP29B | 59.9 | stroma | |
| A0A1S3XX03 | 31 kDa ribonucleoprotein | Q04836 | 31 kDa ribonucleoprotein | 53 | 0 | stroma |
| A0A1S3Z334 | Chloroplast stem-loop binding protein of 41 kDa b | Q9SA52 | Chloroplast stem-loop binding protein of 41 kDa b (CSP41-b) | 85.8 | stroma | |
| A0A1S3ZRR1 | Nucleoid-associated protein At4g30620 | Q9M098 | Nucleoid-associated protein At4g30620 | 76.4 | 1 | stroma |
| A0A1S4CGA5 | Pentatricopeptide repeat-containing protein At4g30825 | O65567 | Pentatricopeptide repeat-containing protein At4g30825 | 63.8 | 9 | stroma |
| A0A1S3YRF9 | Ribosome-recycling factor | Q9M1X0 | Ribosome-recycling factor (RRF) | 64.4 | 0 | stroma |
| Amino acid biosynthesis | ||||||
| A0A1S4CUE0 | Ferredoxin-dependent glutamate synthase | Q9ZNZ7 | Ferredoxin-dependent glutamate synthase 1 (Fd-GOGAT 1) | 83.9 | stroma | |
| A0A1S3YTZ2 | Ketol-acid reductoisomerase | Q05758 | Ketol-acid reductoisomerase | 84.8 | stroma | |
| A0A1S4APF3 | Ornithine carbamoyltransferase | O50039 | Ornithine carbamoyltransferase | 75.3 | stroma | |
| Response to stress | ||||||
| A0A1S4CDL2 | Protein CutA | P93009 | Protein CutA | 69.6 | 1 | inter-membrane |
| A0A1S4A194 | Soluble inorganic pyrophosphatase 6 | Q9LXC9 | Soluble inorganic pyrophosphatase 6 (Ppase 6) | 74.6 | 0 | stroma |
| Chlorophyll synthesis | ||||||
| A0A1S4C5X4 | Oxygen-dependent coproporphyrinogen-III oxidase | Q9LR75 | Coproporphyrinogen-III oxidase 1 (CPOX) | 80.4 | 2 | stroma |
| Photorespiration | ||||||
| A0A1S3X073 | Phosphoglycolate phosphatase 1B | P0DKC4 | Phosphoglycolate phosphatase 1B | 66.8 | 4 | stroma |
| PSII assembly | ||||||
| A0A1S4DN09 | Photosystem II repair protein PSB27-H1 | Q9LR64 | Photosystem II repair protein PSB27-H1 | 64.5 | 0 | thylakoid ls |
| A0A1S4DKC9 | Photosystem II stability/assembly factor HCF136 | O82660 | Photosystem II stability/assembly factor HCF136 | 78.3 | 0 | thylakoid ls |
| Sulfur metabolism | ||||||
| A0A1S4CCJ9 | Cysteine synthase | P47999 | Cysteine synthase | 83.5 | stroma | |
| Unknown function | ||||||
| A0A1S4BU42 | Thylakoid lumenal protein TL20.3 | Q8H1Q1 | Thylakoid lumenal protein TL20.3 | 75.8 | lumen | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancín, M.; Fernandez-Irigoyen, J.; Santamaria, E.; Larraya, L.; Fernández-San Millán, A.; Veramendi, J.; Farran, I. New In Vivo Approach to Broaden the Thioredoxin Family Interactome in Chloroplasts. Antioxidants 2022, 11, 1979. https://doi.org/10.3390/antiox11101979
Ancín M, Fernandez-Irigoyen J, Santamaria E, Larraya L, Fernández-San Millán A, Veramendi J, Farran I. New In Vivo Approach to Broaden the Thioredoxin Family Interactome in Chloroplasts. Antioxidants. 2022; 11(10):1979. https://doi.org/10.3390/antiox11101979
Chicago/Turabian StyleAncín, María, Joaquin Fernandez-Irigoyen, Enrique Santamaria, Luis Larraya, Alicia Fernández-San Millán, Jon Veramendi, and Inmaculada Farran. 2022. "New In Vivo Approach to Broaden the Thioredoxin Family Interactome in Chloroplasts" Antioxidants 11, no. 10: 1979. https://doi.org/10.3390/antiox11101979