
Exogenous L-Glutathione Improves Vitrification Outcomes in Murine Preimplantation Embryos
 , , , , ,
, , , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animal Husbandry and Maintenance
2.3. Experimental Design
2.4. Superovulation, Embryo Collection, and In Vitro Culture
2.5. Morphological Assessment and Grading of Two-Cell Embryos
2.6. Vitrification Warming of Eight-Cell Embryos
2.7. In Vitro Culture and Preimplantation Embryo Developmental Assessment
2.8. Measurement of Intracellular GSH
2.9. Measurement of Intracellular ROS
2.10. Measurement of Cytochrome C Activity
2.11. Reverse Transcription and Microfluidic qPCR
2.12. Statistical Analyses
3. Results and Discussion
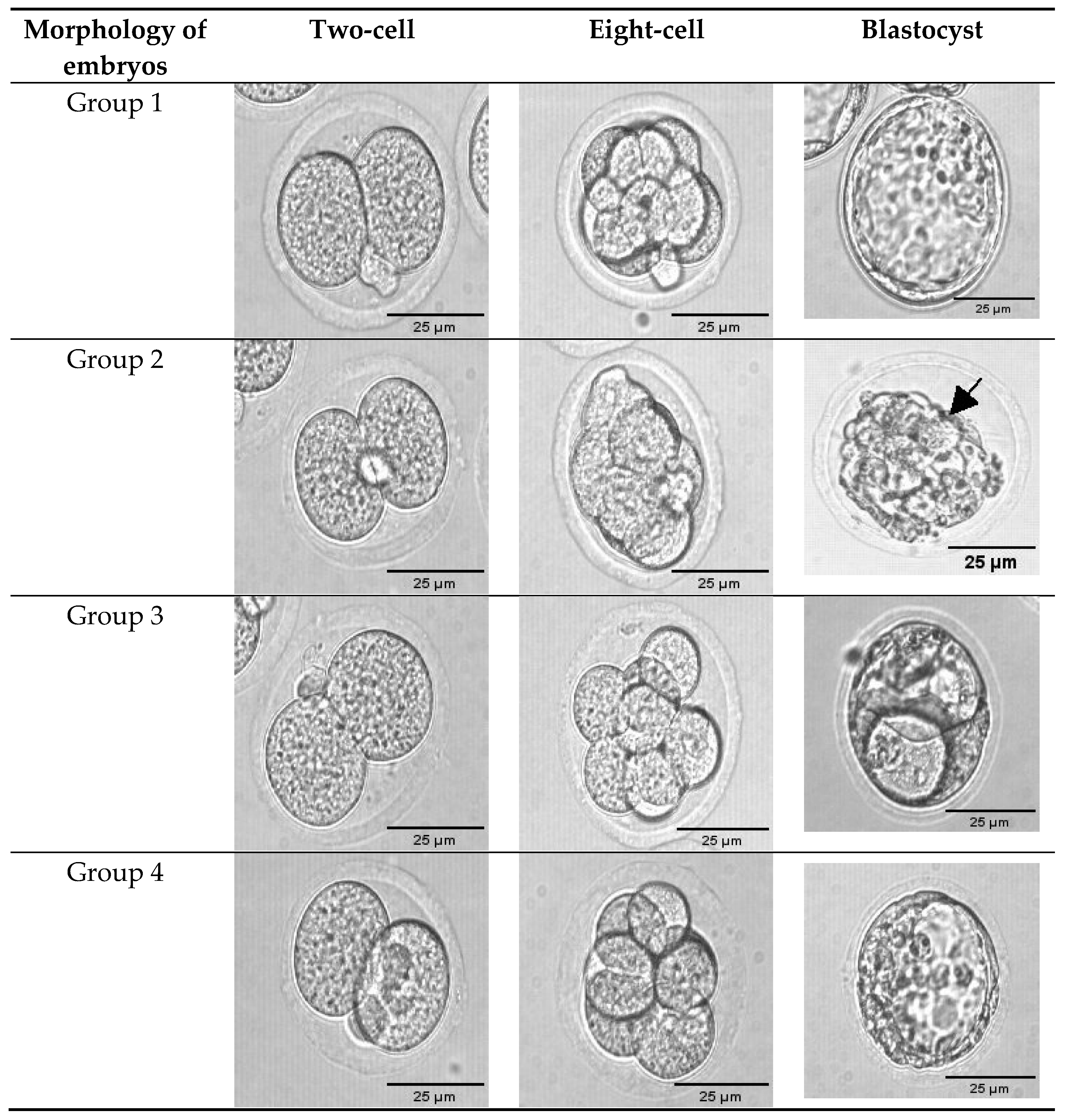
3.1. Exogenous GSH Supplementation Improved Preimplantation Embryo Morphology
3.2. Exogenous GSH Supplementation Enhanced Preimplantation Embryo Development
3.3. Exogenous GSH Supplementation Reduced Oxidative Stress in Vitrified Preimplantation Embryos
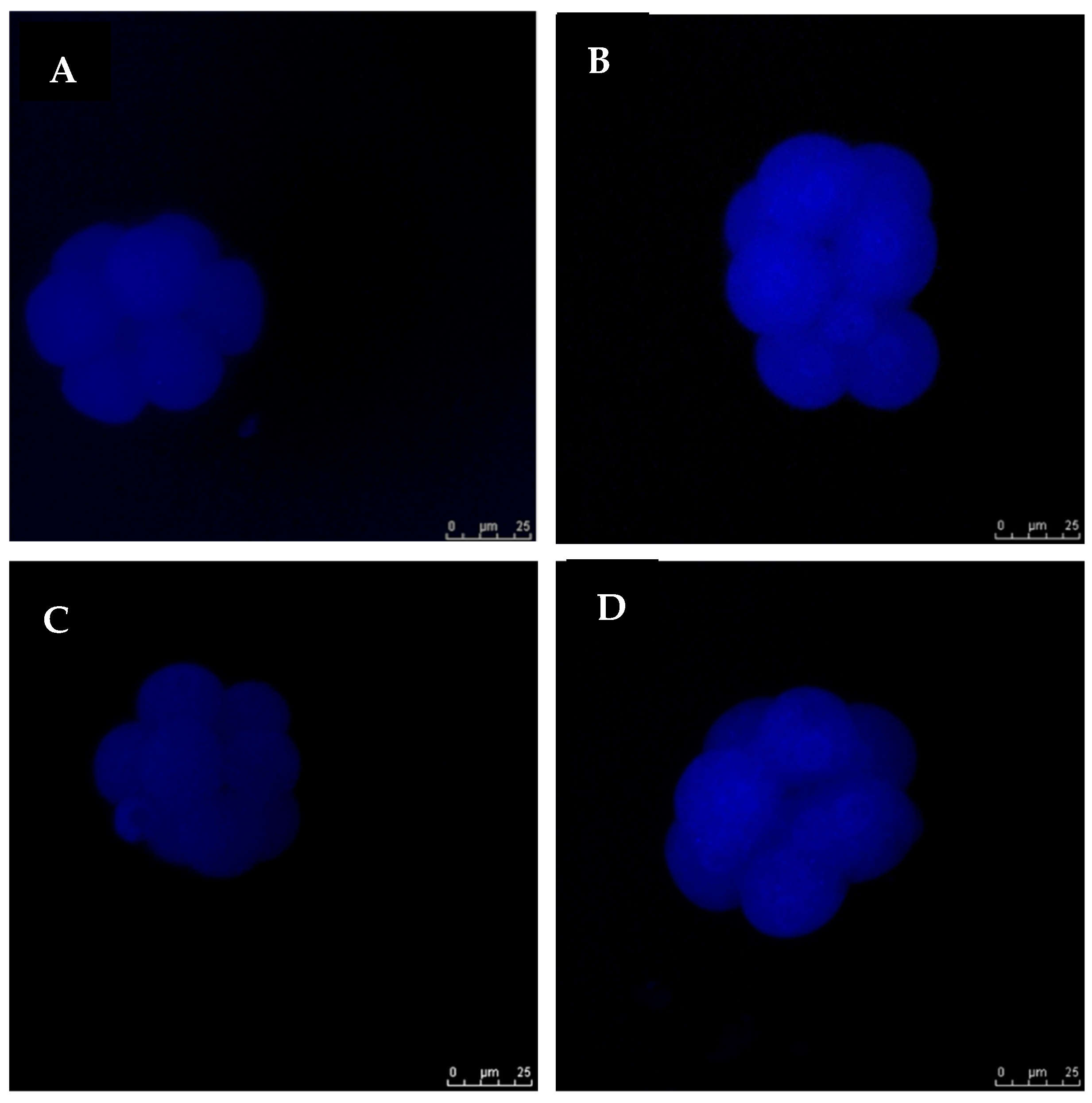
3.4. Exogenous GSH Supplementation Increased Endogenous GSH Levels in Vitrified Preimplantation Embryos
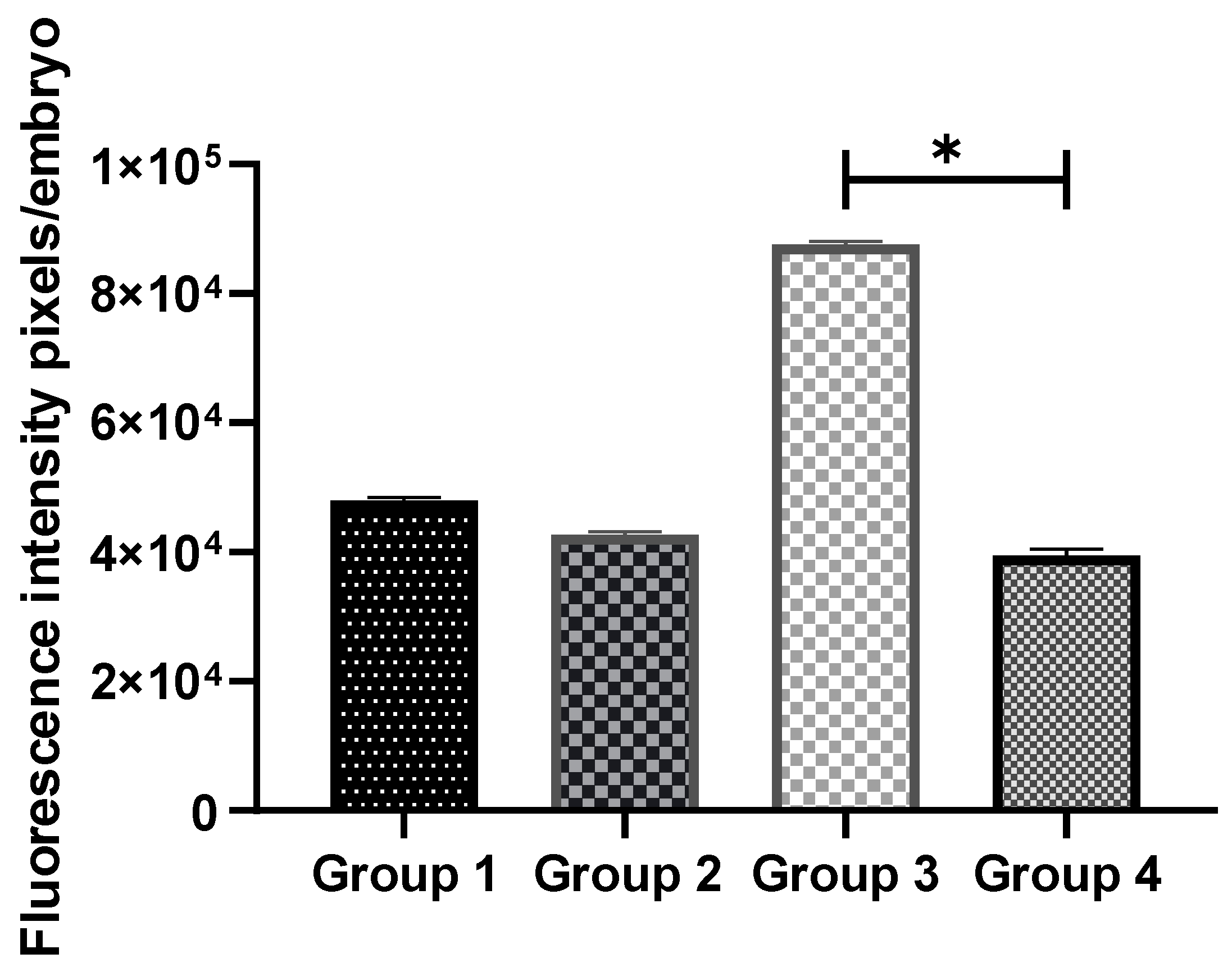
3.5. Exogenous GSH Supplementation Decreased Cytochrome C Expression in Vitrified Preimplantation Embryos
3.6. Exogenous GSH Supplementation Upregulated the Expression of Bcl2 Gene and Downregulated the Expression of Bax Gene in Vitrified Preimplantation Embryo
3.7. Exogenous GSH Supplementation Upregulated the Expression of Gpx1 and Sod1 Genes in Vitrified Preimplantation Embryo
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuller, B.; Paynter, S. Fundamentals of cryobiology in reproductive medicine. Reprod. Biomed. Online 2004, 9, 680–691. [Google Scholar] [CrossRef]
- Larman, M.G.; Minasi, M.G.; Rienzi, L.; Gardner, D.K. Maintenance of the meiotic spindle during vitrification in human and mouse oocytes. Reprod. Biomed. Online 2007, 15, 692–700. [Google Scholar] [CrossRef]
- Dalcin, L.; Silva, R.C.; Paulini, F.; Silva, B.D.; Neves, J.P.; Lucci, C.M. Cytoskeleton structure, pattern of mitochondrial activity and ultrastructure of frozen or vitrified sheep embryos. Cryobiology 2013, 67, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somoskoi, B.; Martino, N.A.; Cardone, R.A.; Lacalandra, G.M.; Dell’Aquila, M.E.; Cseh, S. Different chromatin and energy/redox responses of mouse morulae and blastocysts to slow freezing and vitrification. Reprod. Biol. Endocrinol. 2015, 13, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasiman, R.; Rahman, N.S.A.; Othman, S.; Mustafa, M.F.; Yusoff, N.J.M.; Jusoff, W.H.W.; Khan, N.A.M.N. Cytoskeletal alterations in different developmental stages of in vivo cryopreserved preimplantation murine embryos. Med. Sci. Monit. Basic Res. 2013, 19, 258–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikonomou, Z.; Chatzimeletiou, K.; Sioga, A.; Oikonomou, L.; Tarlatzis, B.C.; Kolibianakis, E. Effects of vitrification on blastomere viability and cytoskeletal integrity in mouse embryos. Zygote 2017, 25, 75–84. [Google Scholar] [CrossRef]
- Tachataki, M.; Winston, R.M.; Taylor, D.M. Quantitative RT–PCR reveals tuberous sclerosis gene, TSC2, mRNA degradation following cryopreservation in the human preimplantation embryo. MHR. Basic Sci. Reprod. Med. 2003, 9, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Larman, M.G.; Katz-Jaffe, M.G.; McCallie, B.; Filipovits, J.A.; Gardner, D.K. Analysis of global gene expression following mouse blastocyst cryopreservation. Hum. Reprod. 2011, 26, 2672–2680. [Google Scholar] [CrossRef] [Green Version]
- Stinshoff, H.; Wilkening, S.; Hanstedt, A.; Brüning, K.; Wrenzycki, C. Cryopreservation affects the quality of in vitro produced bovine embryos at the molecular level. Theriogenology 2011, 76, 1433–1441. [Google Scholar] [CrossRef]
- Monzo, C.; Haouzi, D.; Roman, K.; Assou, S.; Dechaud, H.; Hamamah, S. Slow freezing and vitrification differentially modify the gene expression profile of human metaphase II oocytes. Hum. Reprod. 2012, 27, 2160–2168. [Google Scholar] [CrossRef]
- Sahraei, S.S.; Shahhoseini, M.; Movaghar, B. Vitrification Has an Effect like Culture on Gene Expression and Histone Modification In Mouse Embryos. Cryo Lett. 2018, 39, 102–112. [Google Scholar]
- Harvey, A.J.; Kind, K.L.; Thompson, J.G. REDOX regulation of early embryo development. Reproduction 2002, 123, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Ajduk, A.; Zernicka-Goetz, M. Advances in embryo selection methods. Biol. Rep. 2012, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M. Oxidative stress and redox regulation on in vitro development of mammalian embryos. J. Reprod. Dev. 2012, 58, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of cytochrome c release from mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. BCL-2 family members and the mitochondria in apoptosis. Genes Dev. 1999, 13, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Bratton, S.B.; Salvesen, G.S. Regulation of the Apaf-1–caspase-9 apoptosome. J. Cell Sci. 2010, 123, 3209–3214. [Google Scholar] [CrossRef] [Green Version]
- Trapphoff, T.; Heiligentag, M.; Simon, J.; Staubach, N.; Seidel, T.; Otte, K.; Eichenlaub-Ritter, U. Improved cryotolerance and developmental potential of in vitro and in vivo matured mouse oocytes by supplementing with a glutathione donor prior to vitrification. Mol. Hum. Reprod. 2016, 22, 867–881. [Google Scholar]
- Lane, M.; Maybach, J.M.; Gardner, D.K. Addition of ascorbate during cryopreservation stimulates subsequent embryo development. Hum Reprod. 2002, 17, 2686–2693. [Google Scholar] [CrossRef] [Green Version]
- Somfai, T.; Ozawa, M.; Noguchi, J.; Kaneko, H.; Kuriani Karja, N.W.; Farhudin, M.; Dinnyés, A.; Nagai, T.; Kikuchi, K. Developmental competence of in vitro-fertilized porcine oocytes after in vitro maturation and solid surface vitrification: Effect of cryopreservation on oocyte antioxidative system and cell cycle stage. Cryobiology 2007, 55, 115–126. [Google Scholar] [CrossRef]
- Gupta, M.K.; Uhm, S.J.; Lee, H.T. Effect of vitrification and beta-mercaptoethanol on reactive oxygen species activity and in vitro development of oocytes vitrified before or after in vitro fertilization. Fertil. Steril. 2010, 93, 2602–2607. [Google Scholar] [CrossRef] [PubMed]
- Boonkusol, D.; Gal, A.B.; Bodo, S.; Gorhony, B.; Kitiyanant, Y.; Dinnyes, A. Gene expression profiles and in vitro development following vitrification of pronuclear and 8-cell stage mouse embryos. Mol. Reprod. Dev. 2006, 73, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Mamo, S.; Bodo, S.; Kobolak, J.; Polgar, Z.; Tolgyesi, G.; Dinnyes, A. Gene expression profiles of vitrified in vivo derived 8-cell stage mouse embryos detected by high density oligonucleotide microarrays. Mol. Reprod. Dev. 2006, 73, 1380–1392. [Google Scholar] [CrossRef] [PubMed]
- Balaban, B.; Urman, B.; Sertac, A.; Alatas, C.; Aksoy, S.; Mercan, R. Blastocyst quality affects the success of blastocyst-stage embryo transfer. Fertil. Steril. 2000, 74, 282–287. [Google Scholar] [CrossRef]
- Succu, S.; Bebbere, D.; Bogliolo, L.; Ariu, F.; Fois, S.; Leoni, G.G.; Berlinguer, F.; Naitana, S.; Ledda, S. Vitrification of in vitro matured ovine oocytes affects in vitro pre-implantation development and mRNA abundance. Mol. Reprod. Dev. 2007, 75, 538–546. [Google Scholar] [CrossRef]
- Bartolac, L.K.; Lowe, J.L.; Koustas, G.; Grupen, C.G.; Sjöblom, C. Vitrification, not cryoprotectant exposure, alters the expression of developmentally important genes in in vitro produced porcine blastocysts. Cryobiology 2018, 80, 70–76. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Forouzanfar, M.; Hajian, M.; Asgari, V.; Abedi, P.; Hosseini, L.; Nasr-Esfahani, M.H. Antioxidant supplementation of culture medium during embryo development and/or after vitrification-warming; Which is the most important? J. Assist. Reprod. Genet. 2009, 26, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Dehghani-Mohammadabadi, M.; Salehi, M.; Farifteh, F.; Nematollahi, S.; Arefian, E.; Hajjarizadeh, A.; Parivar, K.; Nourmohammadi, Z. Melatonin modulates the expression of BCL-xl and improve the development of vitrified embryos obtained by IVF in mice. J. Assist. Reprod. Genet. 2014, 31, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Kopeika, J.; Thornhill, A.; Khalaf, Y. The effect of cryopreservation on the genome of gametes and embryos: Principles of cryobiology and critical appraisal of the evidence. Hum. Reprod. Update 2015, 21, 209–227. [Google Scholar] [CrossRef] [Green Version]
- Menezo, Y.J.R.; Silvestris, E.; Dale, B.; Elder, K. Oxidative stress and alterations in DNA methylation: Two sides of the same coin in reproduction. Reprod. Biomed. Online 2016, 33, 668–683. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Majzoub, A. Role of Antioxidants in Assisted Reproductive Techniques. World J. Men’s Health 2017, 35, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Sakatani, M.; Kobayashi, S.; Takahashi, M. Effects of Heat Shock on In Vitro Development and Intracellular Oxidative State of Bovine Preimplantation Embryos. Mol. Reprod. Dev. 2004, 67, 77–82. [Google Scholar] [CrossRef] [PubMed]
- De Matos, D.G.; Furnus, C.C.; Moses, D.F.; Martinez, A.G.; Matkovic, M. Stimulation of glutathione synthesis of in vitro matured bovine oocytes and its effect on embryo development and freezability. Mol. Reprod. Dev. 1996, 45, 451–457. [Google Scholar] [CrossRef]
- Bavister, B.D.; Squirrell, J.M. Mitochondrial distribution and function in oocytes and early embryos. Hum. Reprod. 2000, 15, 189–198. [Google Scholar] [CrossRef]
- Diaz Vivancos, P.; Wolff, T.; Markovic, J.; Pallardo, F.V.; Foyer, C.H. A nuclear glutathione cycle within the cell cycle. Biochem. J. 2010, 431, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.; Chen, C. S-nitrosylation/denitrosylation and apoptosis of immune cells. Cell. Mol. Immunol. 2007, 4, 353–358. [Google Scholar]
- Cooper, A.J.; Pinto, J.T.; Callery, P.S. Reversible and irreversible protein glutathionylation: Biological and clinical aspects. Expert Opin. Drug Metab. Toxicol. 2011, 7, 891–910. [Google Scholar] [CrossRef] [Green Version]
- Zofia, L. The role of glutathione in mammalian gametes. Reprod. Biol. 2005, 5, 5–17. [Google Scholar]
- Hansen, J.M.; Harris, C. Glutathione during embryonic development. Biochim. Biophys. Acta 2015, 1850, 1527–1542. [Google Scholar] [CrossRef]
- Yao, T.; Asayama, Y. Human preimplantation embryo culture media: Past, present, and future. J. Mamm. Ova Res. 2016, 33, 17–34. [Google Scholar] [CrossRef]
- Ozawa, M.; Hirabayashi, M.; Kanai, Y. Developmental competence and oxidative state of mouse zygotes heat-stressed maternally or in vitro. Reproduction 2002, 124, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Yuzawa, K.; Sawada, T.; Hossain, M.S.; Takagi, Y.; Hamano, K.; Tsujii, H. Effect of glutathione on the development of rat embryos following microinsemination. Reprod. Med. Biol. 2009, 8, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhang, H.; Zhang, Y.; Yang, Q.; Zhao, S. Caspase-dependent mitochondrial apoptotic pathway is involved in astilbin-mediated cytotoxicity in breast carcinoma cells. Oncol. Rep. 2018, 40, 2278–2286. [Google Scholar] [CrossRef] [PubMed]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cui, L.; Yu, D.; Hao, H.; Liu, Y.; Zhao, X.; Du, W. Exogenous glutathione improves intracellular glutathione synthesis via the γ-glutamyl cycle in bovine zygotes and cleavage embryos. J. Cell. Physiol. 2019, 234, 7384–7394. [Google Scholar] [CrossRef]
- Cuevas Saiz, I.; Gatell, M.C.P.; Vargas, M.C.; Mendive, A.D.; Enedáguila, N.R.; Solanes, M.M.; Canal, C.B.; López, J.T.; Bonet, A.B.; de Mendoza, M.V.H. The Embryology Interest Group: Updating ASEBIR’s morphological scoring system for early embryos, morulae and blastocysts. Med. Reprod. Y Embriol. Clínica 2018, 5, 42–54. [Google Scholar]
- Kasai, M.; Komi, J.H.; Takakamo, A.; Tsudera, H.; Sakurai, T.; Machida, T. A simple method for mouse embryo cryopreservation in a low toxicity vitrification solution, without appreciable loss of viability. Reproduction 1990, 89, 91–97. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, J.; Ling, X.; Li, X.; Peng, Y.; Guo, X.; Heng, B.C.; Tong, G.Q. Vitrification of mouse embryos at 2-cell, 4-cell and 8-cell stages by cryotop method. J. Assist. Reprod. Genet. 2009, 26, 621–628. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.; Hardarson, T.; Lundin, K. There Is a Cutoff Limit in Diameter Between a Blastomere and a Small Anucleate Fragment. J. Assist. Reprod. Genet. 2003, 20, 309–313. [Google Scholar] [CrossRef]
- Hsieh, C.-Y.; Chen, C.-L.; Yang, K.-C.; Ma, C.-T.; Choi, P.-C.; Lin, C.-F. Detection of Reactive Oxygen Species During the Cell Cycle Under Normal Culture Conditions Using a Modified Fixed-Sample Staining Method. J. Immunoass. Immunochem. 2014, 36, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zatecka, E.; Ded, L.; Elzeinova, F.; Kubatova, A.; Dorosh, A.; Margaryan, H.; Dostalova, P.; Korenkova, V.; Hoskova, K.; Peknicova, J. Effect of zearalenone on reproductive parameters and expression of selected testicular genes in mice. Reprod. Toxicol. 2014, 45, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Ebner, T.; Moser, M.; Sommergruber, M.; Tews, G. Selection based on morphological assessment of oocytes and embryos at different stages of preimplantation development: A review. Hum. Reprod. Update 2003, 9, 251–262. [Google Scholar] [CrossRef]
- Montag, M.; Van Der Ven, H. Evaluation of pronuclear morphology as the only selection criterion for further embryo culture and transfer: Results of a prospective multicentre study. Hum. Reprod. 2001, 16, 2384–2389. [Google Scholar] [CrossRef]
- Tesarik, J.; Greco, E. The probability of abnormal preimplantation development can be predicted by a single static observation on pronuclear stage morphology. Hum. Reprod. 1999, 14, 1318–1323. [Google Scholar] [CrossRef] [Green Version]
- van Royen, E.; Mangelschots, K.; de Neubourg, D.; Valkenburg, M.; van de Meerssche, M.; Ryckaert, G.; Eestermans, W.; Gerris, J. Characterization of a top quality embryo, a step towards single-embryo transfer. Hum. Reprod. 1999, 14, 2345–2349. [Google Scholar] [CrossRef] [Green Version]
- Ziebe, S.; Petersen, K.; Lindenberg, S.; Andersen, A.G.; Gabrielsen, A. Embryo morphology or cleavage stage: How to select the best embryos for transfer after in-vitro fertilization. Hum. Reprod. 1997, 12, 1545–1549. [Google Scholar] [CrossRef]
- Gardner, D.K.; Phil, D.; Lane, M.; Stevens, J.; Schlenker, T.; Schoolcraft, W.B. Blastocyst score affects implantation and pregnancy outcome: Towards a single blastocyst transfer. Fertil. Steril. 2000, 73, 1155–1158. [Google Scholar] [CrossRef]
- Halvaei, I.; Khalili, M.A.; Esfandiari, N.; Safari, S.; Talebi, A.R.; Miglietta, S.; Nottola, S.A. Ultrastructure of cytoplasmic fragments in human cleavage stage embryos. J. Assist. Reprod. Genet. 2016, 33, 1677–1684. [Google Scholar] [CrossRef] [Green Version]
- Lian, H.; Gao, Y.; Jiao, G.; Sun, M.; Wu, X.; Wang, T.; Tan, J. Antioxidant supplementation overcomes the deleterious effects of maternal restraint stress-induced oxidative stress on mouse oocytes. Reproduction 2013, 146, 559–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunawan, M.; Nuriza, N.; Kaiin, E.M.; Sjahfirdi, L. The effect of glutathione antioxidant addition in maturation medium on the morphology of Garut sheep (Ovis aries) oocytes after vitrification. J. Phys. Conf. Ser. 2021, 1725, 012062. [Google Scholar] [CrossRef]
- Chappel, S. The Role of Mitochondria from Mature Oocyte to Viable Blastocyst. Obstet. Gynecol. Int. 2013, 2013, 183024. [Google Scholar] [CrossRef] [Green Version]
- Rienzi, L.; Gracia, C.; Maggiulli, R.; LaBarbera, A.R.; Kaser, D.J.; Ubaldi, F.M.; Racowsky, C. Oocyte, embryo and blastocyst cryopreservation in ART: Systematic review and meta-analysis comparing slow freezing versus vitrification to produce evidence for the development of global guidance. Hum. Reprod. Update 2017, 23, 139–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahromi, H.; Dehnavi, H. Morphological and morphometric study of early-cleavage mice embryos resulting from in vitro fertilization at different cleavage stages after vitrification. Iran. J. Vet. Res. 2016, 17, 55–58. [Google Scholar]
- Xia, P. Intracytoplasmic sperm injection: Correlation of oocyte grade based on polar body, perivitelline space and cytoplasmic inclusions with fertilization rate and embryo quality. Hum. Reprod. 1997, 12, 1750–1755. [Google Scholar] [CrossRef] [PubMed]
- Bartolacci, A.; Intra, G.; Coticchio, G.; Dell’Aquila, M.; Patria, G.; Borini, A. Does morphological assessment predict oocyte developmental competence? A systematic review and proposed score. J. Assist. Reprod. Genet. 2022, 39, 3–17. [Google Scholar] [CrossRef]
- Magli, M.C.; Gianaroli, L.; Ferraretti, A.P. Chromosomal abnormalities in embryos. Mol. Cell. Endocrinol. 2001, 183, S29–S34. [Google Scholar] [CrossRef]
- Maurer, M.; Ebner, T.; Puchner, M.; Mayer, R.B.; Shebl, O.; Oppelt, P.; Duba, H.C. Chromosomal Aneuploidies and Early Embryonic Developmental Arrest. Int. J. Fertil. Steril. 2015, 9, 346–353. [Google Scholar]
- Esfandiari, N.; Burjaq, H.; Gotlieb, L.; Casper, R.F. Brown oocytes: Implications for assisted reproductive technology. Fertil. Steril. 2006, 86, 1522–1525. [Google Scholar] [CrossRef]
- Barnett, D.K.; Bavister, B.D. What is the relationship between the metabolism of preimplantation embryos and their developmental competence? Mol. Reprod. Dev. 1996, 43, 105–133. [Google Scholar] [CrossRef]
- Phan, V.; Littman, E.; Harris, D.; La, A. Correlation between embryo morphology and development and chromosomal complement. Asian Pac. J. Reprod. 2014, 3, 85–89. [Google Scholar] [CrossRef]
- Khalili, M.A.; Anvari, M. The effect of in vitro culture on cleavage rates and morphology of the in vivo-developed embryos in mice. Iran. J. Reprod. Med. 2007, 5, 17–22. [Google Scholar]
- Dinara, S.; Sengoku, K.; Tamate, K.; Horikawa, M.; Ishikawa, M. Effects of supplementation with free radical scavengers on the survival and fertilization rates of mouse cryopreserved oocytes. Hum. Reprod. 2001, 16, 1976–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.J.; Pang, Y.W.; Liu, Y.; Hao, H.S.; Zhao, X.M.; Qin, T.; Du, W.H. Exogenous glutathione supplementation in culture medium improves the bovine embryo development after in vitro fertilization. Theriogenology 2015, 84, 716–723. [Google Scholar] [CrossRef]
- Mukaida, T.; Wada, S.; Takahashi, K.; Pedro, P.B.; An, T.Z.; Kasai, M. Vitrification of human embryos based on the assessment of suitable conditions for 8-cell mouse embryos. Hum. Reprod. 1998, 13, 2874–2879. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Zeng, Y.; Guo, J.; Meng, Q.; Meng, Q.; Jia, G.; Zhu, S. Vitrification transiently alters Oct-4, Bcl2 and P53 expression in mouse morulae but does not affect embryo development in vitro. Cryobiology 2016, 73, 120–125. [Google Scholar] [CrossRef]
- Chaves, D.F.; Corbin, E.; Almiñana, C.; Locatelli, Y.; Souza-Fabjan, J.M.G.; Bhat, M.H.; Mermillod, P. Vitrification of immature and in vitro matured bovine cumulus-oocyte complexes: Effects on oocyte structure and embryo development. Livest. Sci. 2017, 199, 50–56. [Google Scholar] [CrossRef]
- Tait, S.W.G.; Green, D.R. Mitochondrial regulation of cell death. Cold Spring Harb. Perspect. Biol. 2013, 5, a008706. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, Y.; Suzuki, K.; Yoneda, A.; Watanabe, T. Effects of oxygen concentration and antioxidants on the in vitro developmental ability, production of reactive oxygen species (ROS), and DNA fragmentation in porcine embryos. Theriogenology 2004, 62, 1186–1197. [Google Scholar] [CrossRef]
- Martín-romero, F.J.; Miguel-lasobras, E.M.; Domínguez-arroyo, J.A.; González-Carrera, E.; Alvarez, I.S. Contribution of culture media to oxidative stress and its effect on human oocytes. Reprod. Biomed. Online 2008, 17, 652–661. [Google Scholar] [CrossRef]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth Defects Res. Part C Embryo Today Rev. 2007, 81, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.K.; Kumar, A.; Hitchcock, D.B.; Fan, D.; Goodwin, R.; Lavoie, H.A.; Singh, U.S. Resveratrol prevents embryonic oxidative stress and apoptosis associated with diabetic embryopathy and improves glucose and lipid profile of diabetic dam. Mol. Nutr. Food Res. 2011, 55, 1186–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truong, T.T.; Soh, Y.M.; Gardner, D.K. Antioxidants improve mouse preimplantation embryo development and viability. Hum. Reprod. 2016, 31, 1445–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moawad, A.R.; Tan, S.L.; Taketo, T. Beneficial effects of glutathione supplementation during vitrification of mouse oocytes at the germinal vesicle stage on their preimplantation development following maturation and fertilization in vitro. Cryobiology 2017, 76, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Guo, N.; Liu, J.-Q.; Tan, M.-H.; Li, Y.-F. Vitrification of in vitro matured oocytes: Effects on meiotic spindle configuration and mitochondrial function. Int. J. Clin. Exp. Pathol. 2014, 7, 1159–1165. [Google Scholar]
- Mahfouz, M.; Kummerow, F. Vitamin C or Vitamin B6 supplementation prevent the oxidative stress and decrease of prostacyclin generation in homocysteinemic rats. Int. J. Biochem. Cell Biol. 2004, 36, 1919–1932. [Google Scholar] [CrossRef]
- Gardiner, C.S.; Reed, D.J. Status of Glutathione during Oxidant-Induced Oxidative Stress in the Preimplantation Mouse Embryo1. Biol. Reprod. 1994, 51, 1307–1314. [Google Scholar] [CrossRef] [Green Version]
- Jang, W.; Lee, S.; Choi, H.; Lim, J.; Heo, Y.; Cui, X.; Kim, N. Vitrification of immature-mouse oocytes by the modified-cut standard straw method. Cell Biol. Int. 2013, 38, 164–171. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Li, X.-X.; Xu, Y.-K.; Wu, H.; Zheng, J.-J.; Yu, X.-L. Developmental competence and gene expression of immature oocytes following liquid helium vitrification in bovine. Cryobiology 2014, 69, 428–433. [Google Scholar] [CrossRef]
- Kuzmany, A.; Havlicek, V.; Wrenzycki, C.; Wilkening, S.; Brem, G.; Besenfelder, U. Expression of mRNA, before and after freezing, in bovine blastocysts cultured under different conditions. Theriogenology 2011, 75, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Madrid Gaviria, S.; López Herrera, A.; Urrego, R.; Restrepo Betancur, G.; Echeverri Zuluaga, J.J. Effect of resveratrol on vitrified in vitro produced bovine embryos: Recovering the initial quality. Cryobiology 2019, 89, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.A.; Zangeronimo, M.G.; Castillo-Martín, M.; Gadani, B.; Chaves, B.R.; Rodríguez-Gil, J.E.; Yeste, M. Supplementing maturation medium with insulin growth factor I and vitrification-warming solutions with reduced glutathione enhances survival rates and development ability of in vitro matured vitrified-warmed pig oocytes. Front. Physiol. 2019, 9, 1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Gu, R.; Lu, X.; Zhao, S.; Feng, Y.; Sun, Y. Preincubation with glutathione ethyl ester improves the developmental competence of vitrified mouse oocytes. J. Assist. Reprod. Genet. 2018, 35, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Glutathione and modulation of cell apoptosis. Biochim. Et Biophys. Acta 2012, 1823, 1767–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Tian, X.; Zhang, L.; Tan, D.; Reiter, R.J.; Liu, G. Melatonin promotes the in vitro development of pronuclear embryos and increases the efficiency of blastocyst implantation in murine. J. Pineal Res. 2013, 55, 267–274. [Google Scholar] [CrossRef]
- Gao, C.; Han, H.-B.; Tian, X.-Z.; Tan, D.-X.; Wang, L.; Zhou, G.-B.; Zhu, S.-E.; Liu, G.-S. Melatonin promotes embryonic development and reduces reactive oxygen species in vitrified mouse 2-cell embryos. J. Pineal Res. 2011, 52, 305–311. [Google Scholar] [CrossRef]
- Castillo-Martín, M.; Yeste, M.; Soler, A.; Morató, R.; Bonet, S. Addition of L-ascorbic acid to culture and vitrification media of IVF porcine blastocysts improves survival and reduces HSPA1A levels of vitrified embryos. Reprod. Fertil. Dev. 2015, 27, 1115–1123. [Google Scholar] [CrossRef]
- Zarbakhsh, S. Effect of antioxidants on preimplantation embryo development in vitro: A review. Zygote 2021, 29, 179–193. [Google Scholar] [CrossRef]
- Roy, P.; Lei, X.G. Knockout of SOD1 or GPX1 led to decreased bone remodeling in young adult female mice. FASEB J. 2007, 21, A114. [Google Scholar] [CrossRef]
- Ribas, V.; García-Ruiz, C.; Fernández-Checa, J.C. Glutathione and mitochondria. Front. Pharmacol. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
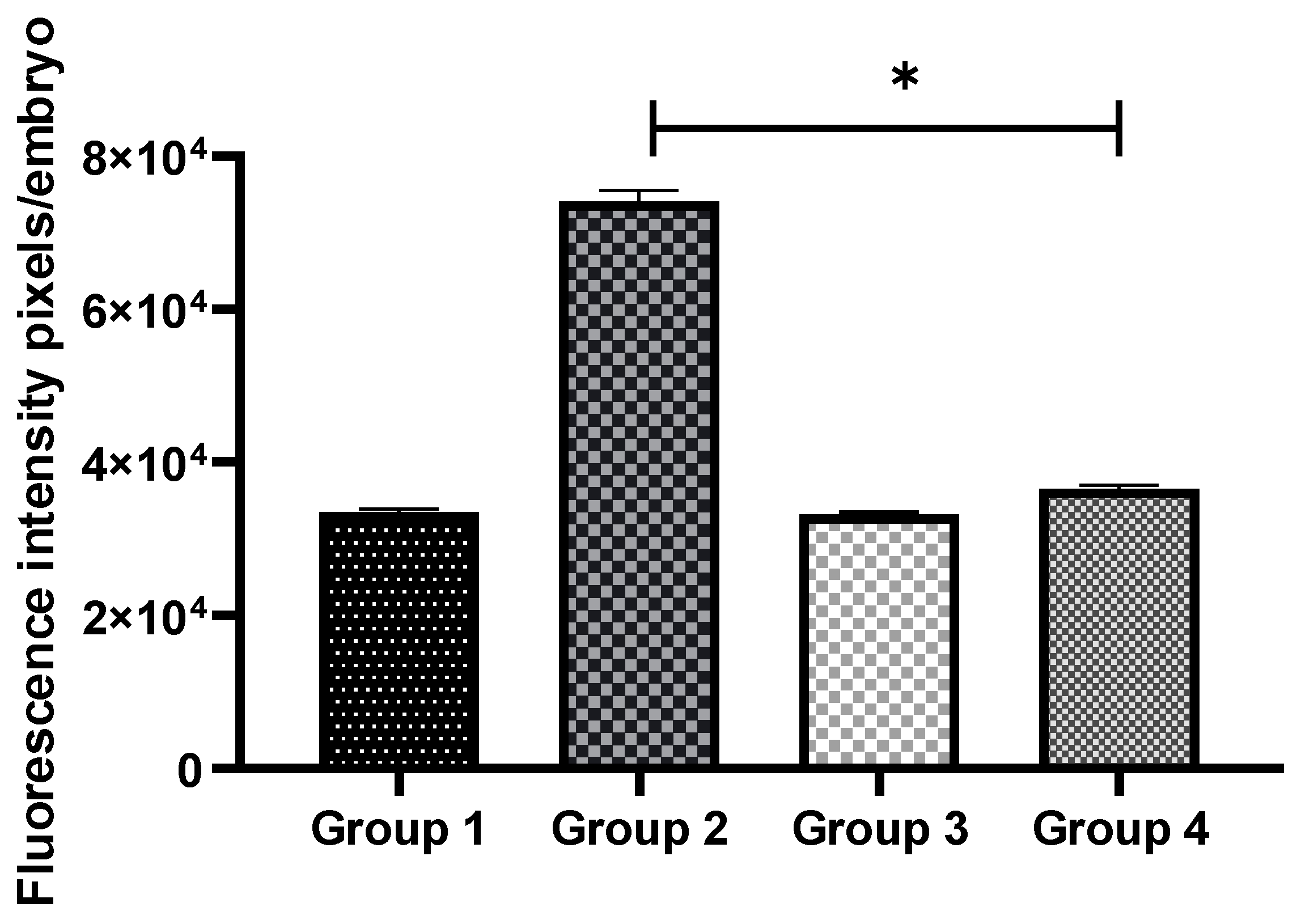{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
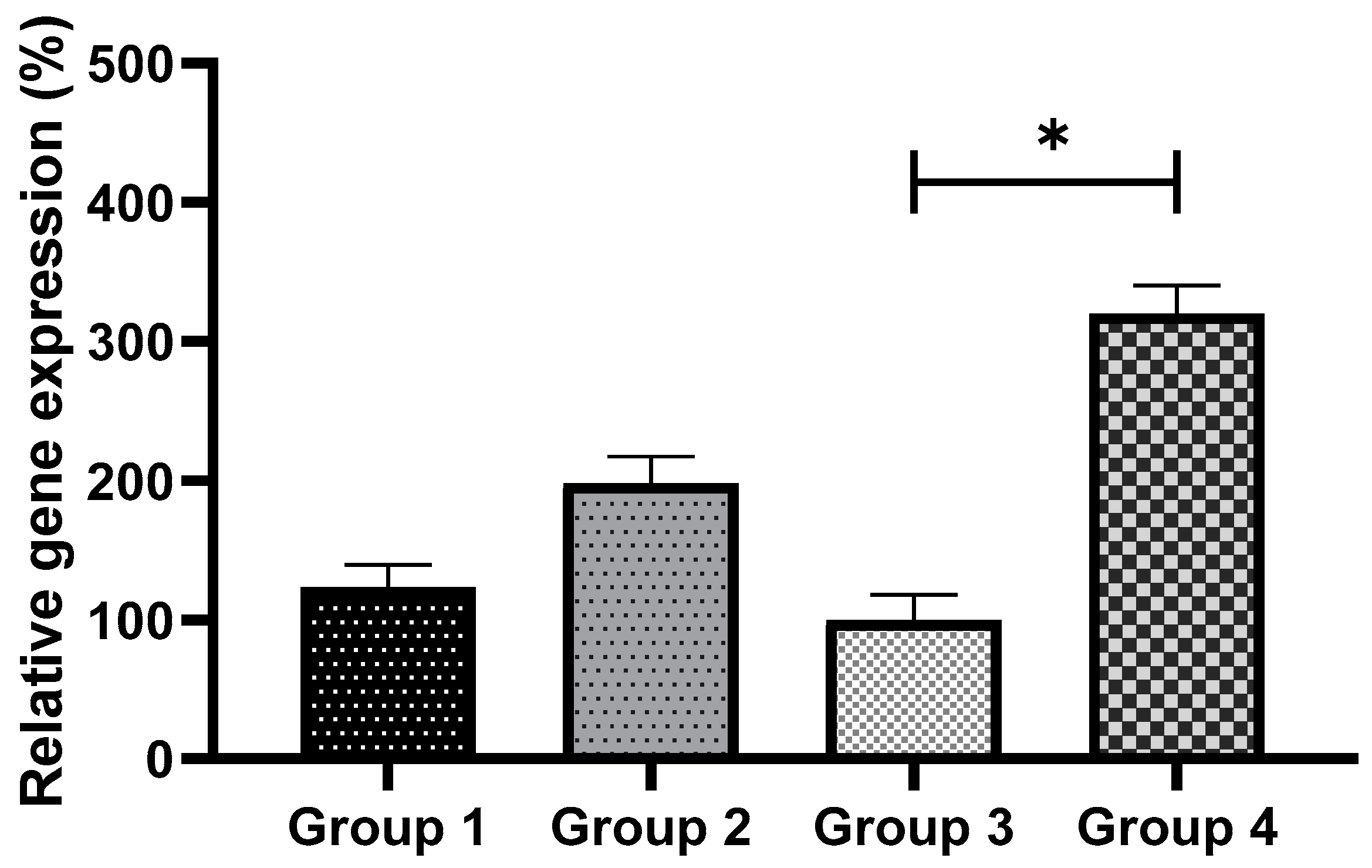{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) Forward | Primer Sequence (3′–5′) Reverse | Accession Number |
|---|---|---|---|
| Gapdh | CAAGGTCATCCCAGAGCTGAA | CAGATCCACGACGGACACA | NM_001289726.N |
| Bax | ATGTGTGTGGAGAGCGTCAA | CTGATCAGCTCGGGCACTTTA | NM_007527.N |
| Bcl2 | GCGTGGTTGCCCTCTTCTA | GATGCCGGTTCAGGTACTCA | NM_009741.N |
| Gpx1 | TCGGTTTCCCGTGCAATCA | GTCGGACGTACTTGAGGGAA | NM_008160.N |
| Sod1 | CTCACTCTCAGGAGAGCATTCC | TTCCACCTTTGCCCAAGTCA | NM_011434.N |
| Groups | No. of Two-Cell Embryos (%) | No. of Eight-Cell Embryos (%) | No. of Blastocysts (%) |
|---|---|---|---|
| n | (n ± SD) | (n ± SD) | |
| Group 1 | 76 (100%) | 21.00 ± 3.49 (83%) | 17.67 ± 3.06 (69%) a |
| Group 2 | 60 (100%) | 15.00 ± 2.86 (75%) | 12.67 ± 1.89 (63%) a |
| Group 3 | 111 (100%) | 31.67± 5.14 (86%) | 29.67 ± 0.62 (80%) b |
| Group 4 | 90 (100%) | 26.67± 2.78 (89%) | 24.33 ± 1.43 (81%) c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul Rahman, N.-S.; Mohamed Noor Khan, N.-A.; Eshak, Z.; Sarbandi, M.-S.; Mohammad Kamal, A.-A.; Abd Malek, M.; Abdullah, F.; Abdullah, M.A.; Othman, F. Exogenous L-Glutathione Improves Vitrification Outcomes in Murine Preimplantation Embryos. Antioxidants 2022, 11, 2100. https://doi.org/10.3390/antiox11112100
Abdul Rahman N-S, Mohamed Noor Khan N-A, Eshak Z, Sarbandi M-S, Mohammad Kamal A-A, Abd Malek M, Abdullah F, Abdullah MA, Othman F. Exogenous L-Glutathione Improves Vitrification Outcomes in Murine Preimplantation Embryos. Antioxidants. 2022; 11(11):2100. https://doi.org/10.3390/antiox11112100
Chicago/Turabian StyleAbdul Rahman, Nor-Shahida, Nor-Ashikin Mohamed Noor Khan, Zolkapli Eshak, Mimi-Sophia Sarbandi, Aqila-Akmal Mohammad Kamal, Mastura Abd Malek, Fathiah Abdullah, Maizaton Atmadini Abdullah, and Fezah Othman. 2022. "Exogenous L-Glutathione Improves Vitrification Outcomes in Murine Preimplantation Embryos" Antioxidants 11, no. 11: 2100. https://doi.org/10.3390/antiox11112100