
Concurrent Production of α- and β-Carotenes with Different Stoichiometries Displaying Diverse Antioxidative Activities via Lycopene Cyclases-Based Rational System

 , ,
, , 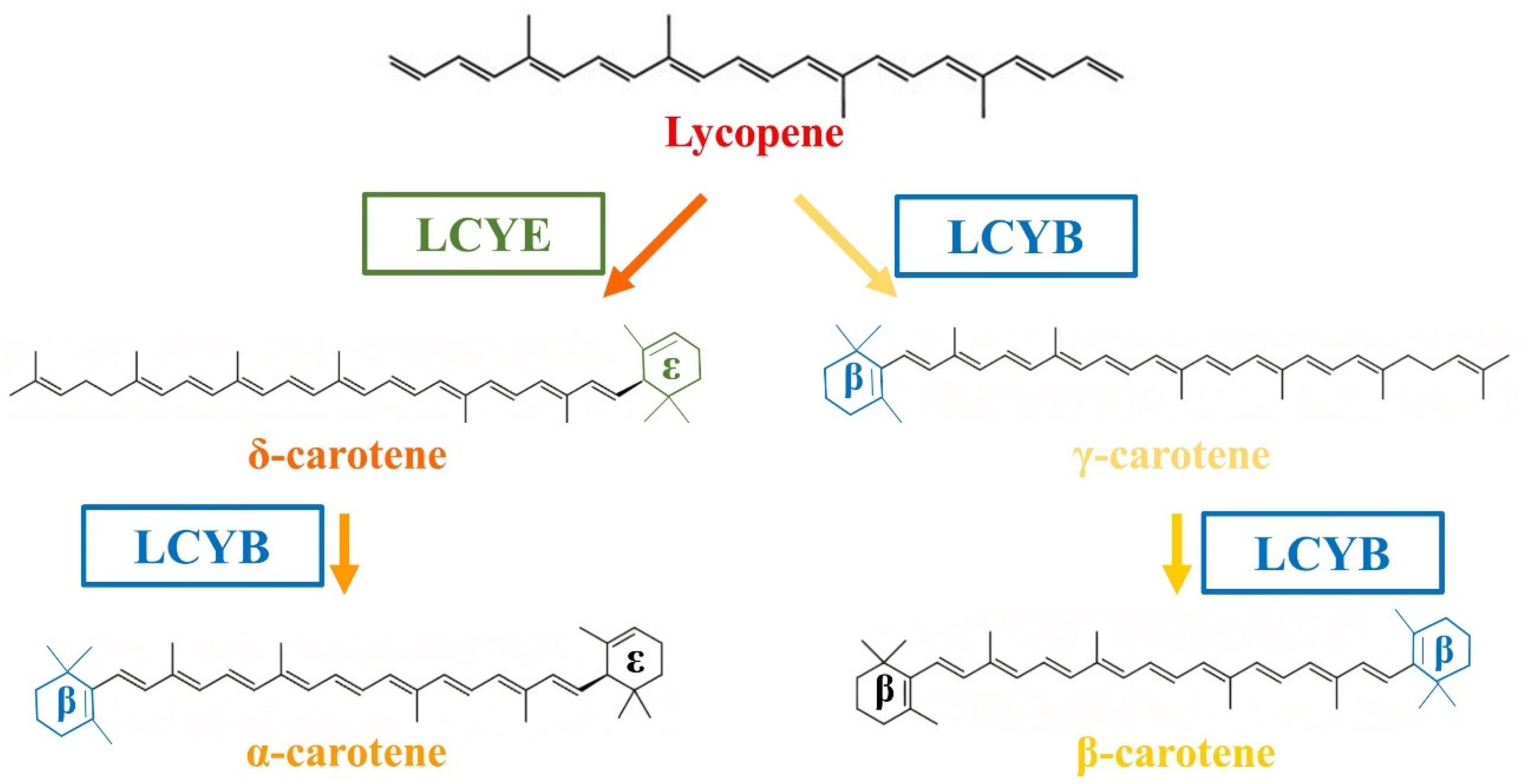{kind=link}
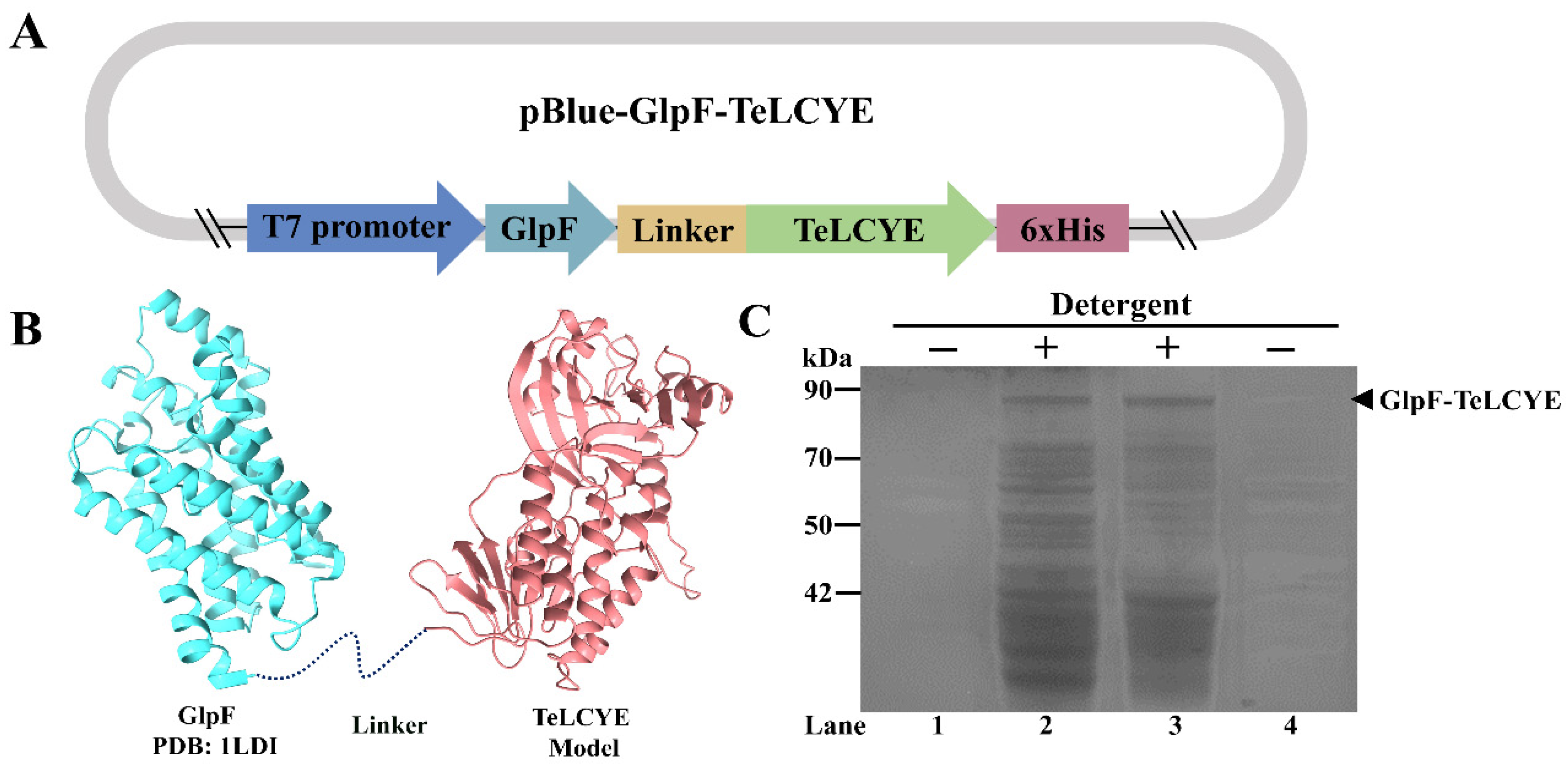{kind=link}
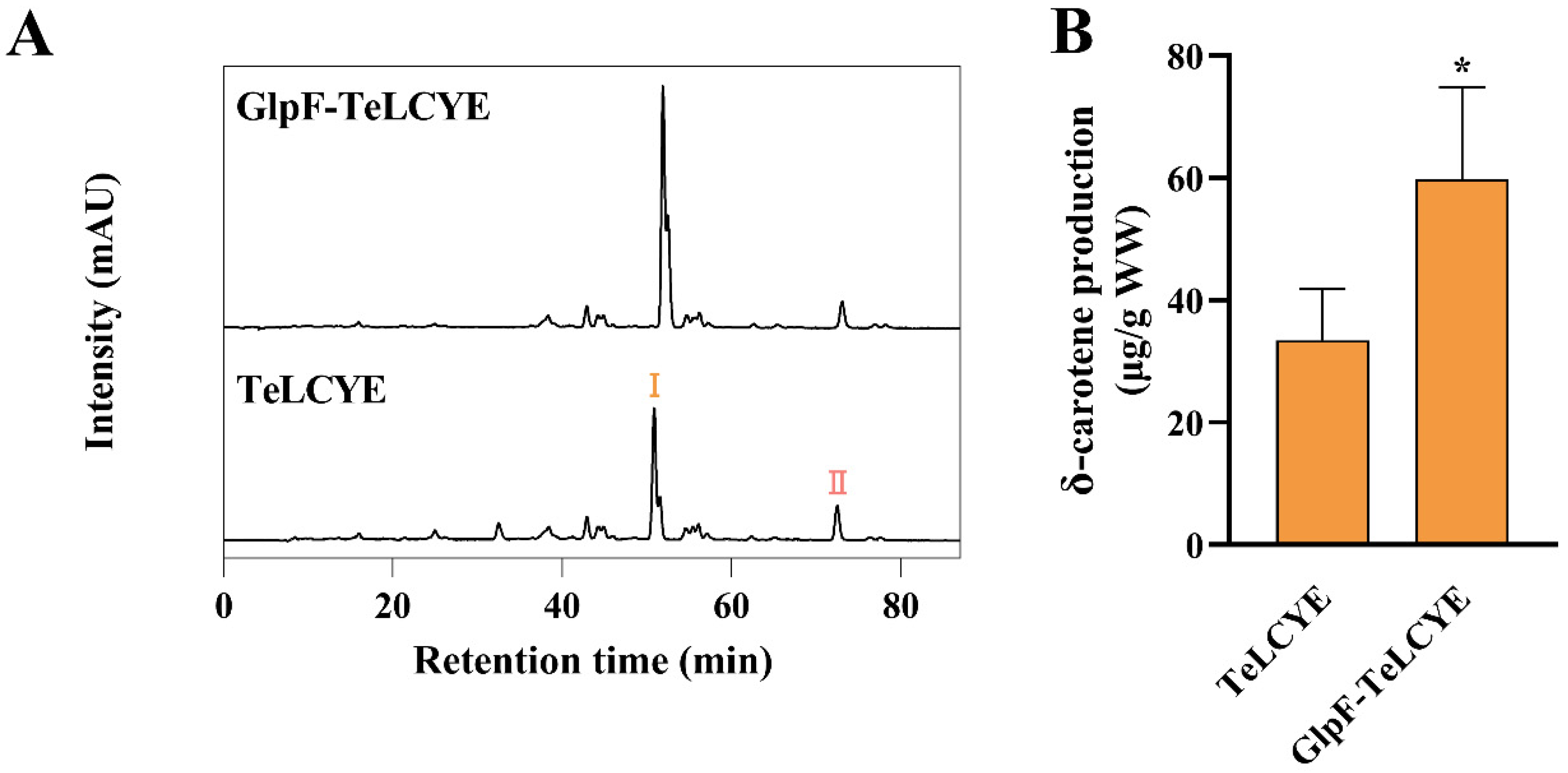{kind=link}
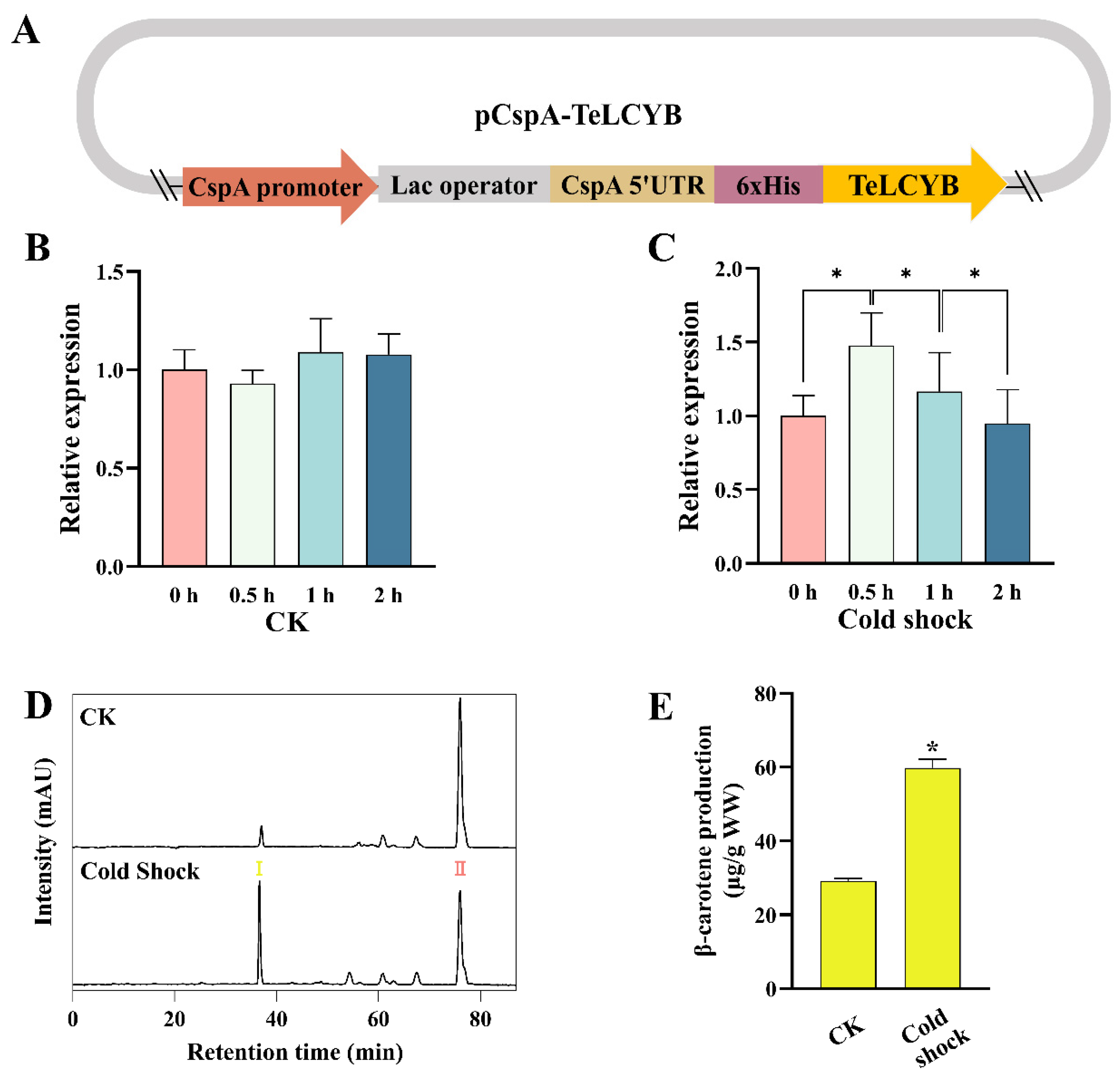{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Reagents
2.2. Plasmid Constructions for TeLCYE and TeLCYB
2.3. Protein Expression Validation of Membrane-Targeted TeLCYE
2.4. Quantitative Real-Time Reverse Transcription PCR
2.5. Carotenoid Quantification
2.6. Determination of Antioxidant Property
2.6.1. DPPH Free Radical Scavenging Ability Assay
2.6.2. Determination of ABTS Free Radical Scavenging Ability
2.6.3. Determination of Ferric-Reducing Antioxidant Power (FRAP)
2.6.4. Determination of MDA Content
2.7. Statistical Analysis
3. Results and Discussion
3.1. Construction and Expression of Membrane-Targeted TeLCYE
3.2. TeLCYE Catalytic Efficiency Enhanced via Spatially Membrane-Targeted Regulation
3.3. TeLCYB Catalytic Activity Reprogammed via Temporally Promoter-Centered Regulation
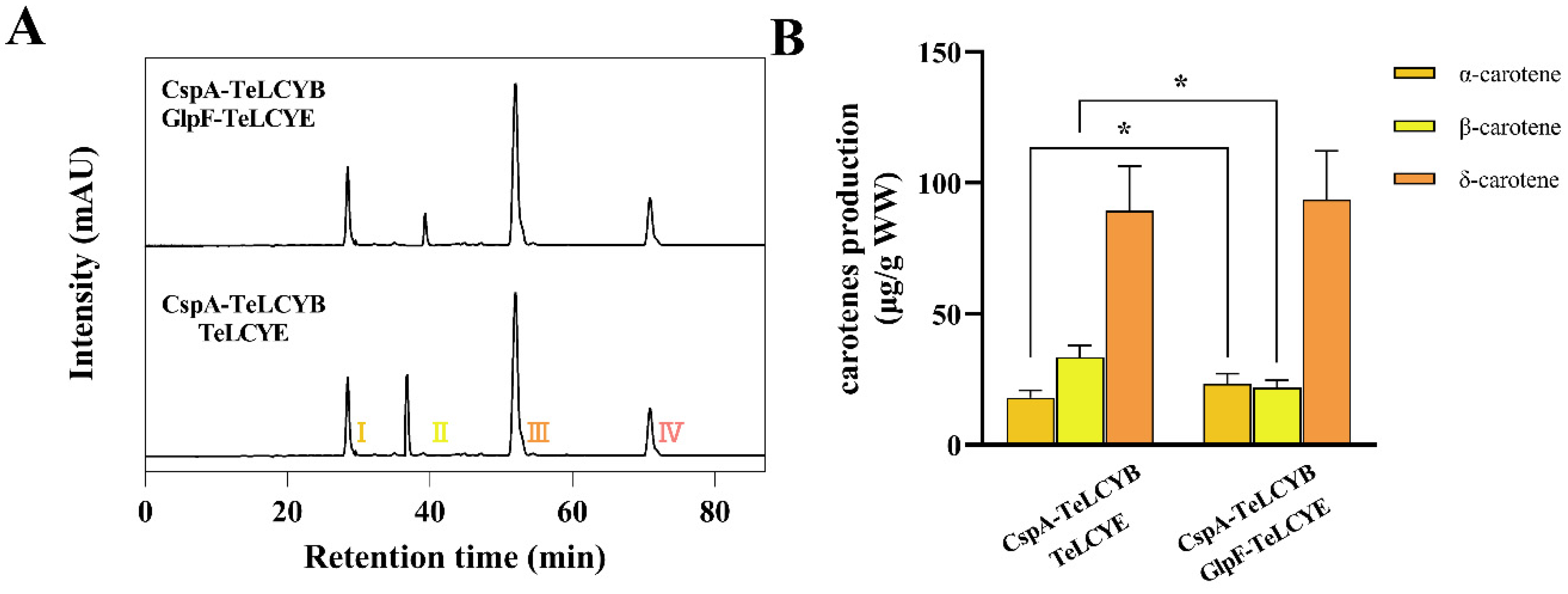
3.4. The Initially Efficient Boosted TeLCYE Is Essential for High α-/β-Carotene Stoichiometry
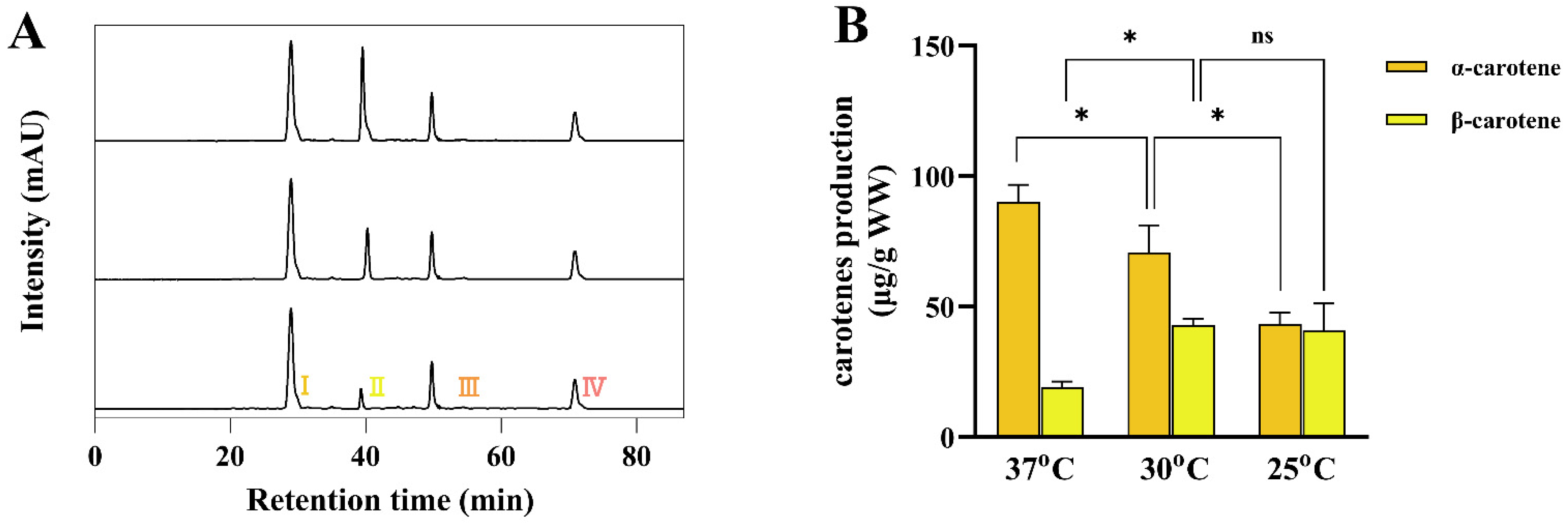
3.5. The High α-/β-Carotene Stoichiometries Induced by Dynamically Temperature-Programmed Regulation
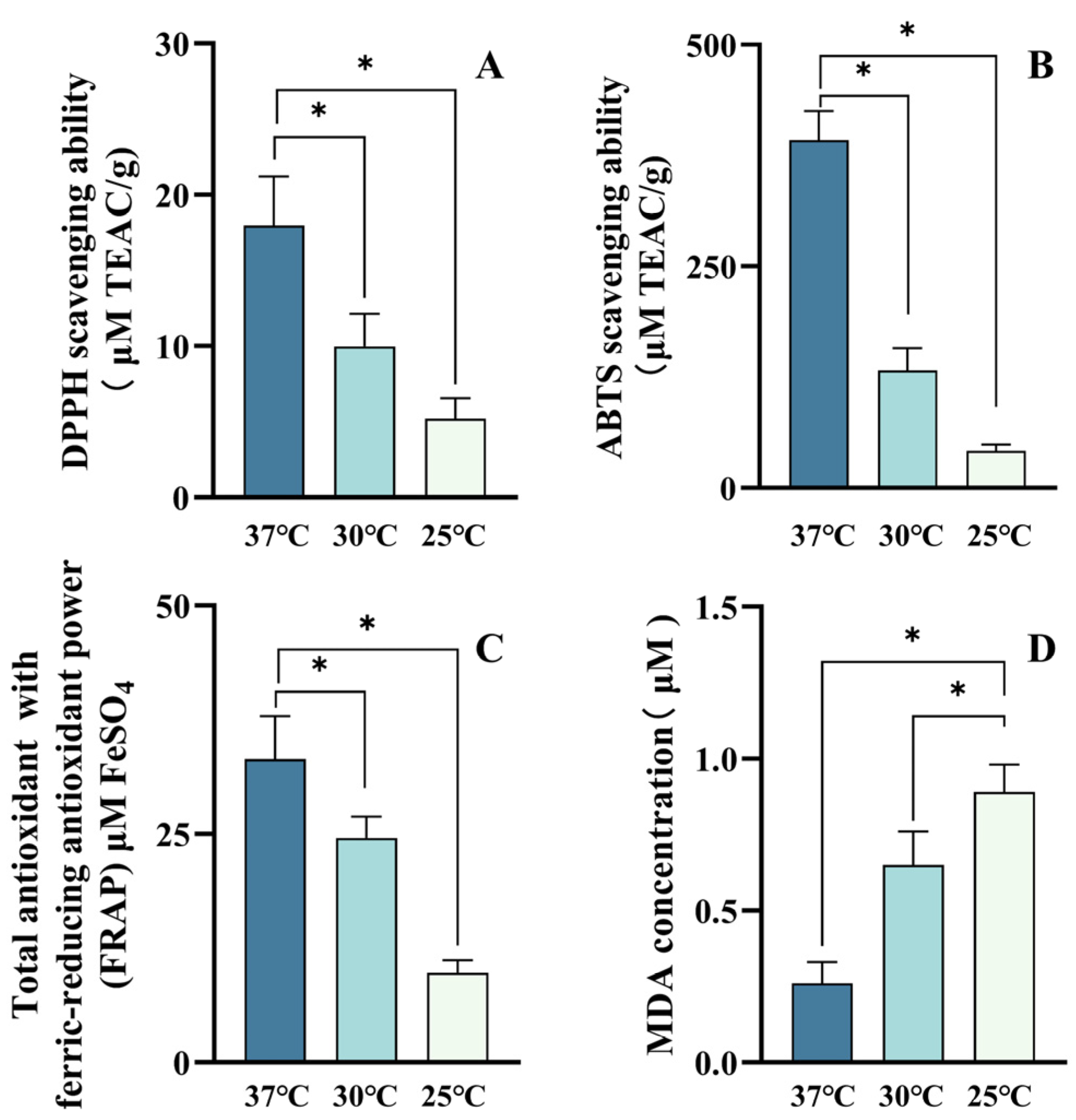
3.6. High α-/β-Carotene Stoichiometries Contributed Distinct Antioxidant Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Englert, M.; Hammann, S.; Vetter, W. Isolation of β-carotene, α-carotene and lutein from carrots by countercurrent chromatography with the solvent system modifier benzotrifluoride. J. Chromatogr. A 2015, 1388, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hu, B.; Cheng, Y.; Guo, Y.; Yao, W.; Qian, H. Carotenoids from fungi and microalgae: A review on their recent production, extraction, and developments. Bioresour. Technol. 2021, 337, 125398. [Google Scholar] [CrossRef] [PubMed]
- During, A.; Harrison, E.H. Intestinal absorption and metabolism of carotenoids: Insights from cell culture. Arch. Biochem. Biophys. 2004, 430, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Böhm, V.; Borge, G.I.A.; Cano, M.P.; Fikselová, M.; Gruškienė, R.; Lavelli, V.; Loizzo, M.R.; Mandic, A.I.; Brahm, P.M.; et al. Carotenoids: Considerations for their use in functional foods, nutraceuticals, nutricosmetics, supplements, botanicals, and novel foods in the context of sustainability, circular economy, and climate change. Annu. Rev. Food Sci. Technol. 2021, 12, 433–460. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ford, E.S.; Zhao, G.; Balluz, L.S.; Giles, W.H.; Liu, S. Serum α-carotene concentrations and risk of death among US Adults: The Third National Health and Nutrition Examination Survey Follow-up Study. Arch. Intern. Med. 2011, 171, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Murakoshi, M.; Nishino, H. Potent Cancer Chemopreventive Activity of α-Carotene against Carcinogenesis; Springer: Tokyo, Japan, 1997; pp. 533–537. [Google Scholar]
- Yang, Z.; Chen, W. Physiological significance of link between polycyclic aromatic hydrocarbons and serum nutritional biomarkers. Food Chem. 2021, 353, 129445. [Google Scholar] [CrossRef]
- Wawrzyniak, A.; Hamułka, J.; Friberg, E.; Wolk, A. Dietary, anthropometric, and lifestyle correlates of serum carotenoids in postmenopausal women. Eur. J. Nutr. 2013, 52, 1919–1926. [Google Scholar] [CrossRef]
- Shalini, T.; Jose, S.S.; Prasanthi, P.S.; Balakrishna, N.; Viswanath, K.; Reddy, G.B. Carotenoid status in type 2 diabetes patients with and without retinopathy. Food Funct. 2021, 12, 441–442. [Google Scholar] [CrossRef]
- Shahriarpour, Z.; Nasrabadi, B.; Hejri-Zarifi, S.; Shariati-Bafghi, S.; Yousefian-Sanny, M.; Karamati, M.; Rashidkhani, B. Oxidative balance score and risk of osteoporosis among postmenopausal Iranian women. Arch. Osteoporos. 2021, 16, 4. [Google Scholar] [CrossRef]
- Liu, X.; Dhana, K.; Furtado, J.D.; Agarwal, P.; Aggarwal, N.T.; Tangney, C.; Laranjo, N.; Carey, V.; Barnes, L.L.; Sacks, F.M. Higher circulating α-carotene was associated with better cognitive function: An evaluation among the MIND trial participants. J. Nutr. Sci. 2021, 10, e64. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Iso, H. Chapter 21-Overview of the role of antioxidant vitamins as protection against cardiovascular disease: Implications for aging. In Aging; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 213–224. [Google Scholar]
- Qu, M.; Shi, H.; Wang, K.; Wang, X.; Yu, N.; Guo, B. The associations of plasma/serum carotenoids with alzheimer’s disease: A systematic review and meta-analysis. J. Alzheimers Dis. 2021, 82, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Furtado, J.D.; Cronjé, H.T.; DeKosky, S.T.; Fitzpatrick, A.L.; Lopez, O.L.; Kuller, L.H.; Mukamal, K.J.; Jensen, M.K. Plasma antioxidants and risk of dementia in older adults. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2021, 7, e12208. [Google Scholar] [CrossRef] [PubMed]
- Arango, J.; Jourdan, M.; Geoffriau, E.; Beyer, P.; Welsch, R. Carotene hydroxylase activity determines the levels of both α-carotene and total carotenoids in orange carrots. Plant Cell 2014, 26, 2223–2233. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; He, W.; Li, D.; Bao, Y.; Riaz, A.; Xiao, Y.; Song, J.; Liu, C. Effect of methyl jasmonate on carotenoids biosynthesis in germinated maize kernels. Food Chem. 2020, 307, 125525. [Google Scholar] [CrossRef] [PubMed]
- Burri, B.J.; Chang, J.S.T.; Neidlinger, T.R. β-Cryptoxanthin- and α-carotene-rich foods have greater apparent bioavailability than β-carotene-rich foods in Western diets. Br. J. Nutr. 2010, 105, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Libson, A.; Miercke, L.; Weitzman, C.; Nollert, P.; Krucinski, J.; Stroud, R.M. Structure of a glycerol-conducting channel and the basis for its selectivity. Science 2000, 290, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.S.; Eweys, A.S.; Zhang, J.Y.; Zhu, Y.; Bai, J.; Darwesh, O.M.; Zhang, H.B.; Xiao, X. Fermentation affects the antioxidant activity of plant-based food material through the release and production of bioactive components. Antioxidants 2021, 10, 2004. [Google Scholar] [CrossRef]
- Shin, K.C.; Seo, M.J.; Kim, Y.S.; Yeom, S.J. Molecular Properties of β-Carotene Oxygenases and Their Potential in Industrial Production of Vitamin A and Its Derivatives. Antioxidants 2022, 11, 1180. [Google Scholar] [CrossRef]
- Siziya, I.N.; Hwang, C.Y.; Seo, M.J. Antioxidant Potential and Capacity of Microorganism-Sourced C30 Carotenoids—A Review. Antioxidants 2022, 11, 1963. [Google Scholar] [CrossRef]
- Blatt, A.; Bauch, M.E.; Pörschke, Y.; Lohr, M. A lycopene β-cyclase/lycopene ε-cyclase/light-harvesting complex-fusion protein from the green alga Ostreococcus lucimarinus can be modified to produce α-carotene and β-carotene at different ratios. Plant J. 2015, 82, 582–595. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Swofford, C.A.; Rückert, C.; Chatzivasileiou, A.O.; Sinskey, A.J. Heterologous production of α-carotene in Corynebacterium glutamicum using a multi-copy chromosomal integration method. Bioresour. Technol. 2021, 241, 125782. [Google Scholar] [CrossRef] [PubMed]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef]
- Schaub, P.; Yu, Q.; Gemmecker, S.; Poussin-Courmontagne, P.; Mailliot, J.; Mcewen, A.G.; Ghisla, S.; Al-Babili, S.; Cavarelli, J.; Beyer, P. On the structure and function of the phytoene desaturase CRTI from Pantoea ananatis, a membrane-peripheral and FAD-dependent oxidase/isomerase. PLoS ONE 2012, 7, e39550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, L.; Zhu, X.; Wu, T.; Wang, W.; Zhao, D.; Bi, C.; Zhang, X. Optimizing the localization of astaxanthin enzymes for improved productivity. Biotechnol. Biofuels 2018, 11, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Q.; Zhou, P.; Yao, Z.; Li, M.; Yu, H.; Ye, L. Heterologous biosynthesis of lutein in S. cerevisiae enabled by temporospatial pathway control. Metab. Eng. 2021, 67, 19–28. [Google Scholar] [CrossRef]
- Siefermann-Harms, D. Light and temperature control of season-dependent changes in the α- and β-carotene content of spruce needles. J. Plant Physiol. 1994, 143, 488–494. [Google Scholar] [CrossRef]
- Qing, G.; Ma, L.; Khorchid, A.; Swapna, G.V.T.; Mal, T.K.; Takayama, M.M.; Xia, B.; Phadtare, S.; Ke, H.; Acton, T.; et al. Cold-shock induced high-yield protein production in Escherichia coli. Nat. Biotechnol. 2004, 22, 877–882. [Google Scholar] [CrossRef]
- Cunningham, F.X.; Sun, Z.; Chamovitz, D.; Gantt, H.E. Molecular structure and enzymatic function of lycopene cyclase from the cyanobacterium Synechococcus sp. Strain PCC7942. Plant Cell 1994, 6, 1107–1121. [Google Scholar]
- Sawczuk, R.; Karpińska, J.; Filipowska, D.; Bajguz, A.; Hryniewicka, M. Evaluation of total phenols content, anti-DPPH activity and the content of selected antioxidants in the honeybee drone brood homogenate. Food Chem. 2021, 368, 130745. [Google Scholar] [CrossRef]
- Müller, L.; Froehlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Kim, D.; Cho, W.; Yeon, S.; Choi, S.; Lee, C. Effects of lotus (Nelumbo nucifera) leaf on quality and antioxidant activity of yogurt during refrigerated storage. Food Sci. Anim. Resour. 2019, 39, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Hossen, S.; Sukhan, Z.P.; Cho, Y.; Lee, W.K.; Kho, K.H. Antioxidant activity and oxidative stress-oriented apoptosis pathway in saccharides supplemented cryopreserved sperm of pacific abalone, Haliotis discus hannai. Antioxidants 2022, 11, 1303. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.H.; Zhu, J.; Jiang, J.G. Carotenoids biosynthesis and cleavage related genes from bacteria to plants. Crit. Rev. Food Technol. 2018, 58, 2314–2333. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, Y.; Wang, W.; Cao, Z.; Fu, Q.; Bao, M.; He, Y. Functional analysis of the marigold (Tagetes erecta) lycopene ε-cyclase (TeLCYe) promoter in transgenic tobacco. Mol. Biotechnol. 2019, 61, 703–713. [Google Scholar] [CrossRef]
- Lee, P.C.; Yoon, Y.G.; Schmidt-Dannert, C. Investigation of cellular targeting of carotenoid pathway enzymes in Pichia pastoris. J. Biotechnol. 2009, 140, 227–233. [Google Scholar] [CrossRef]
- Villar-Martínez, A.A.D.; García-Saucedo, P.A.; Carabez-Trejo, A.; Cruz-Hernández, A.; Paredes-López, O. Carotenogenic gene expression and ultrastructural changes during development in marigold. J. Plant Physiol. 2005, 162, 1046–1056. [Google Scholar] [CrossRef]
- Xu, Q.; Eguchi, T.; Mathews, I.I.; Rife, C.L.; Chiu, H.J.; Farr, C.L.; Feuerhelm, J.; Jaroszewski, L.; Klock, H.E.; Knuth, M.W. Insights into substrate specificity of geranylgeranyl reductases revealed by the structure of digeranylgeranylglycerophospholipid reductase, an essential enzyme in the biosynthesis of archaeal membrane lipids. J. Mol. Biol. 2010, 404, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Goldenberg, D.; Azar, I.; Oppenheim, A.B. Differential mRNA stability of the cspA gene in the cold-shock response of Escherichia coli. Mol. Microbiol. 1996, 19, 241–248. [Google Scholar] [CrossRef]
- Phadtare, S. Recent developments in bacterial cold-shock response. Curr. Issues Mol. Biol. 2004, 6, 125–136. [Google Scholar]
- Fu, X.; Cheng, S.; Feng, C.; Kang, M.; Huang, B.; Jiang, Y.; Duan, X.; Grierson, D.; Yang, Z. Lycopene cyclases determine high α-/β-carotene ratio and increased carotenoids in bananas ripening at high temperatures. Food Chem. 2019, 283, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.; Stinco, C.M.; Meléndez-Martínez, A.J. Free radical scavenging properties of phytofluene and phytoene isomers as compared to lycopene: A combined experimental and theoretical study. J. Phys. Chem. B 2014, 1, 9819–9825. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Takaichi, S.; Steiger, S.; Wang, Z.Y.; Sandmann, G. Novel hydroxycarotenoids with improved antioxidative properties produced by gene combination in Escherichia coli. Nat. Biotechnol. 2000, 18, 843–846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hou, J.; Yuan, Q.; Xin, P.; Cheng, H.; Gu, Z.; Wu, J. Arginine derivatives assist dopamine-hyaluronic acid hybrid hydrogels to have enhanced antioxidant activity for wound healing. Chem. Eng. J. 2020, 392, 123775. [Google Scholar] [CrossRef]
- Mansour, A.T.; Mahboub, H.H.; Elshopakey, G.E.; Aziz, E.K.; Alhajji, A.H.M.; Rayan, G.; Ghazzawy, H.S.; El-Houseiny, W. Physiological performance, antioxidant and immune status, columnaris resistance, and growth of nile tilapia that received Alchemilla vulgaris-supplemented diets. Antioxidants 2022, 11, 1494. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, H.; He, W.; Dai, Z.; Zhang, Z.; Bao, Y.; Li, D.; Zhu, P. Concurrent Production of α- and β-Carotenes with Different Stoichiometries Displaying Diverse Antioxidative Activities via Lycopene Cyclases-Based Rational System. Antioxidants 2022, 11, 2267. https://doi.org/10.3390/antiox11112267
Luo H, He W, Dai Z, Zhang Z, Bao Y, Li D, Zhu P. Concurrent Production of α- and β-Carotenes with Different Stoichiometries Displaying Diverse Antioxidative Activities via Lycopene Cyclases-Based Rational System. Antioxidants. 2022; 11(11):2267. https://doi.org/10.3390/antiox11112267
Chicago/Turabian StyleLuo, Hao, Weiwei He, Zhuqing Dai, Zhongyuan Zhang, Yihong Bao, Dajing Li, and Ping Zhu. 2022. "Concurrent Production of α- and β-Carotenes with Different Stoichiometries Displaying Diverse Antioxidative Activities via Lycopene Cyclases-Based Rational System" Antioxidants 11, no. 11: 2267. https://doi.org/10.3390/antiox11112267
APA StyleLuo, H., He, W., Dai, Z., Zhang, Z., Bao, Y., Li, D., & Zhu, P. (2022). Concurrent Production of α- and β-Carotenes with Different Stoichiometries Displaying Diverse Antioxidative Activities via Lycopene Cyclases-Based Rational System. Antioxidants, 11(11), 2267. https://doi.org/10.3390/antiox11112267