
Role for Plant-Derived Antioxidants in Attenuating Cancer Cachexia


Abstract
:
1. Introduction
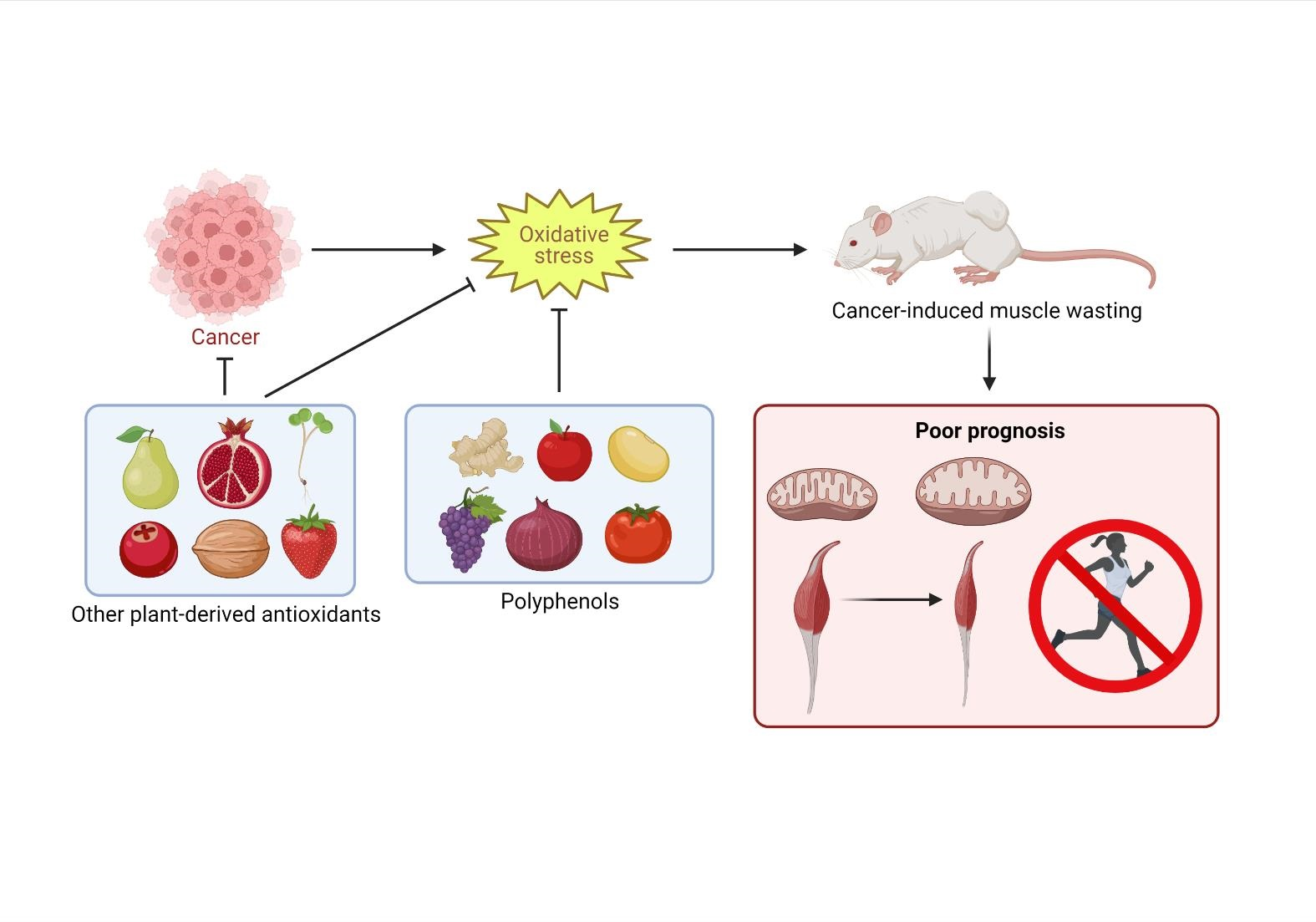
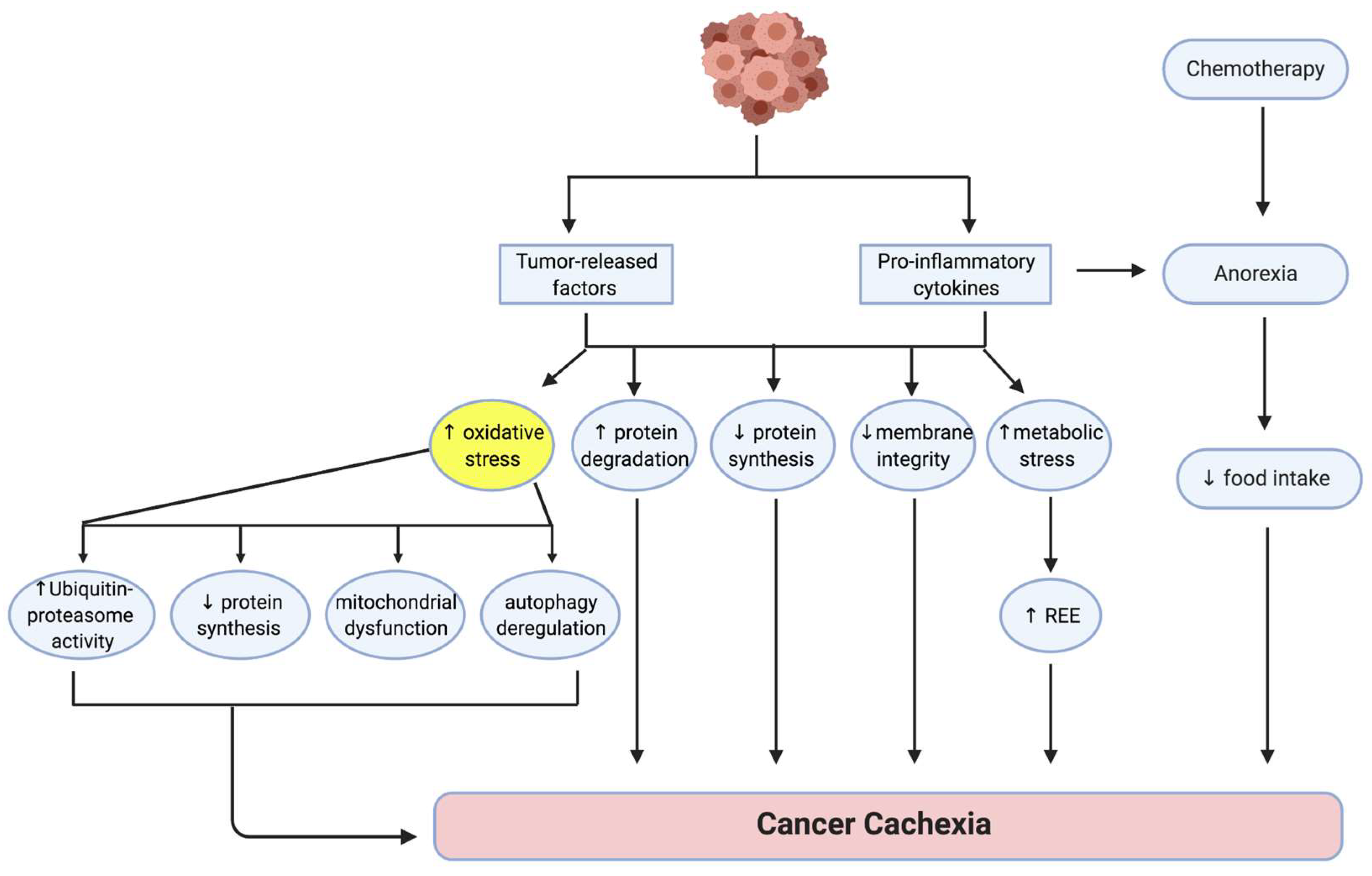
2. Pathogenesis of Cancer Cachexia
3. The Role of Oxidative Stress in the Etiology of Cancer Cachexia
4. Signaling Pathways Involved in Oxidative Stress
4.1. Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells Signaling Pathway
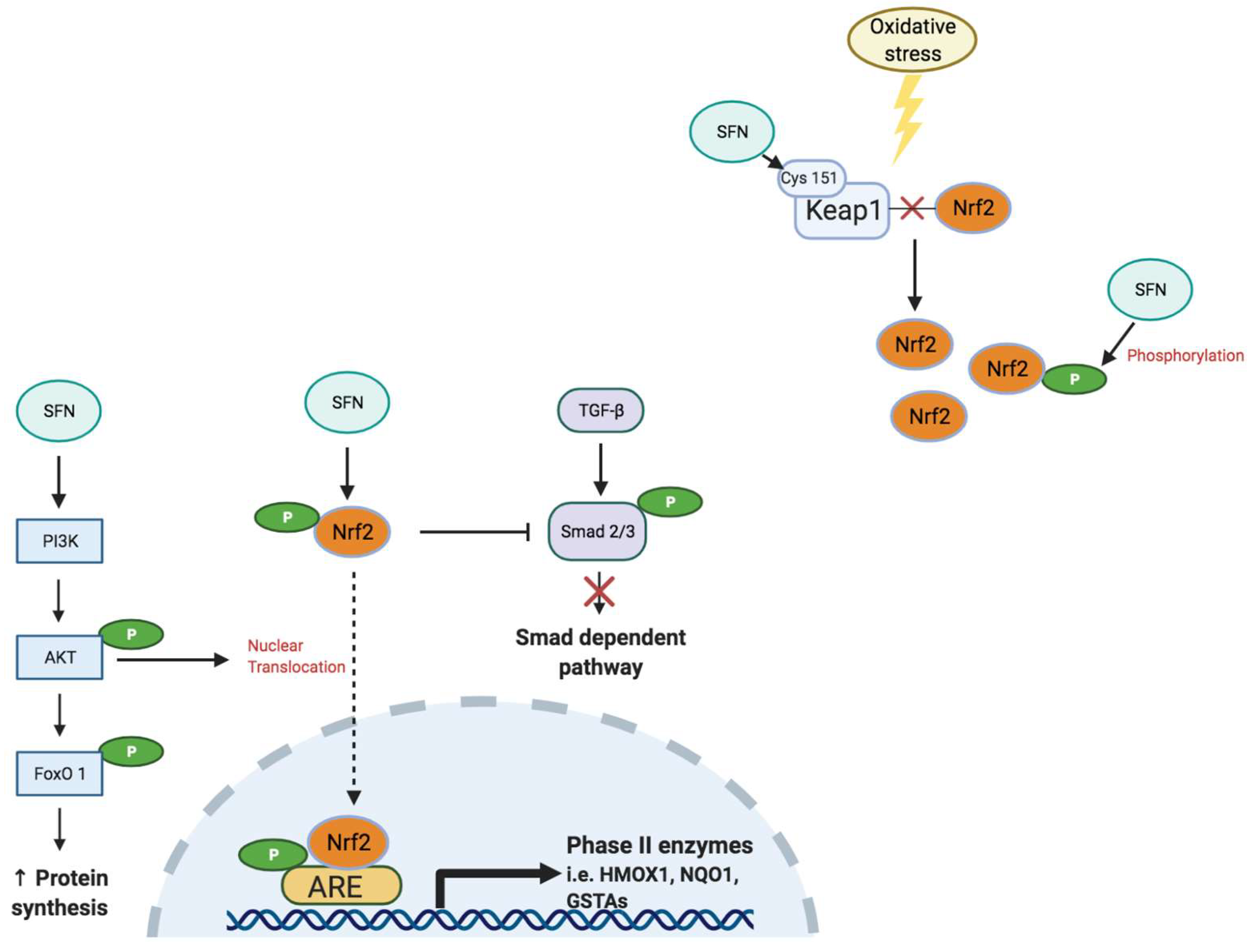
4.2. Nuclear Factor Erythroid 2-Related Factor 2
5. Polyphenols to Reduce Oxidative Stress in Cancer Cachexia
5.1. Epigallocatechin-3-Gallate
5.2. Resveratrol

{kind=link}
{kind=link}
{kind=link}
| Types | Experimental Setting | Treatments | Findings | References |
|---|---|---|---|---|
| EGCG | In vivo 6–8-week-old male LLC-tumor-bearing mice (C57BL/6) | Low dose (0.2 mg/kg/day), high dose (0.6 mg/kg/day) via oral gavage; | ↓ NF-κB | [34] |
| ↓ NF-κB-mediated ubiquitin– proteasome proteolysis | ||||
| 12 days pre-treatment or 30 days post-tumor treatment | ↓ atrogin-1 and MuRF1 expression | |||
| ↓ tumor-induced muscle atrophy | ||||
| Resveratrol | In vivo 6–10-week-old female C-26 tumor-bearing mice (CD2F1) | 200 mg/kg/day via oral gavage for 11 days | ↓ NF-κB | [56] |
| ↓ atrogin-1 and MuRF1 expression | ||||
| ↓ tumor-induced muscle atrophy | ||||
| No effect on tumor growth | ||||
| In vivo 5-week-old male Wistar AH-130 tumor-bearing rats | 1 mg/kg/day via intraperitoneal (i.p.) injection to AH-130 tumor bearing rats for 7 days | No effect on skeletal muscle and whole body mass | [57] | |
| 12-week-old male LLC-tumor-bearing mice (C57BL/6) | 5 or 25 mg/kg/day via i.p. injection to LLC-tumor bearing mice for 15 days | Failed to attenuate cancer cachexia in different tumor-bearing rodents | ||
| In vivo 10-week-old female BALB/c mice | 20 mg/kg/day via i.p. injection for 15 days | ↓ muscle wasting | [58] | |
| ↑ gastrocnemius and soleus muscle mass | ||||
| ↓ tumor growth | ||||
| ↑ limb strength gain | ||||
| ↑ muscle fiber (I & II) cross-sectional area, ↓ muscle abnormalities | ||||
| ↑ sirtuin-1 protein expression | ||||
| ↓ atrogin-1 and MuRF1 expression | ||||
| ↓ forkhead box O3 (FoxO3) | ||||
| ↓ signaling markers NF-κB and p50 | ||||
| Curcumin | In vivo 10-week-old female LP07 tumor-bearing BALB/c mice | 1 mg/kg/day via i.p. injection for 15 days | ↓ muscle wasting | [58] |
| ↑ gastrocnemius and soleus muscle mass | ||||
| ↑ limb strength gain | ||||
| No effect on tumor growth | ||||
| ↑ muscle fiber (I & II) cross-sectional area, ↓ muscle abnormalities | ||||
| ↑ sirtuin-1 protein expression | ||||
| ↓ atrogin-1 and MuRF1 expression | ||||
| ↓ FoxO3 | ||||
| ↓ signaling markers NF-κB and p50 | ||||
| In vivo MAC16-colon tumor-bearing mice | Low dose (100 mg/kg/day), high dose (250 mg/kg/day) via oral gavage for 20 days | ↓ muscle wasting with low dosage | [28] | |
| ↑ body weight, muscle hypertrophy with high dosage | ||||
| ↓ proteasome complex activity | ||||
| Inhibited NF-κB pathway | ||||
| In vivo Male Wistar AH-130 tumor-bearing rats | 20 μg/kg body weight via i.p. injection for 6 days | ↓ tumor growth | [59] | |
| Failed to attenuate cancer cachexia | ||||
| Carnosol | In vitro C2C12 myotube | 3.125 μM to 25 μM concentration of carnosol incubated with C-26 cancer medium for 48 h in C2C12 myotubes; | In vitro: High dose (25 μM) had no toxic effect to C2C12 myotubes; | [60] |
| ↓ C-26 tumor-induced muscle wasting in C2C12 myotubes in dose-dependent manner | ||||
| ↑ MyoD, p-Akt at high dose of carnosol | ||||
| ↓ MuRF1, p-p65/p65 at high dose of carnosol | ||||
| In vivo 6–8-week-old male C-26 tumor-bearing, BALB/c mice | 10 mg/kg/day via i.p. injection from the day after tumor injection for 16 days | In vivo: ↑ body weight | ||
| No effect on tumor growth | ||||
| ↑ MyoD, myosin heavy chain | ||||
| ↓ p-p65/p65 ratio | ||||
| Quercetin | In vivo 15-week-old ApcMin/+mice | 25 mg/kg/day via oral gavage for 3 weeks | Attenuated ↓ body mass | [61] |
| ↑ gastrocnemius and quadriceps muscle mass | ||||
| No change in soleus muscle mass | ||||
| No improvement in muscle function | ||||
| ↓ plasma IL-6 | ||||
| In vivo 9-week-old C-26 tumor-bearing male CD2F1 mice | 250 mg/kg added to daily chow diet for 20 days | ↑ body weight | [62] | |
| ↑ food intake | ||||
| No change grip strength | ||||
| Prevented tumor-induced ↓ muscle volume | ||||
| No change in tumor weight | ||||
| ↑ gastrocnemius and tibialis anterior muscle mass | ||||
| Rutin | In vivo 6-week-old K14-HPV16 mice | 413 mg/kg/day to daily diet for 24 weeks | ↑ survival | [63] |
| No change in body weight | ||||
| ↑ gastrocnemius muscle weight | ||||
| ↓ NF-κB signaling pathway | ||||
| Genistein and daidzein | In vivo 8-week-old male C57BL/6 mice with LLC tumors | Normal diet mixed with 40.74% of soyaflavone HG (containing high genistein and daidzein contents) for 3 weeks | No change in food intake or body mass | [64] |
| ↑ gastrocnemius muscle weight and myofiber size | ||||
| No change in tumor mass | ||||
| No change in plasma IL-6 or TNF-α | ||||
| ↓ atrogin-1 and MuRF1 expression | ||||
| ↓ phosphorylation of extracellular signal-regulated kinase (ERK) | ||||
| Morin | In vitro LLC cells and C2C12 myotubes | In vitro: 10, 50, 100, 200 μM treated to LLC cells and C2C12 myotubes for 48 h | In vitro: ↓ cell viability of LLC cells with 100 and 200 μM | [65] |
| ↑ cell viability of C2C12 myotubes with 10 μM; no cell death at high dose (100 and 200 μM) | ||||
| ↓ protein synthesis shown in LLC cells using SUnSET method; no significant changes were found with C2C12 myotubes. | ||||
| In vivo 6-week-old male C57BL/6 mice with LLC tumors | In vivo: Morin-rich (0.1% w/w) diet for 3 weeks | In vivo: Attenuated↓muscle mass and gastrocnemius muscle myofiber size | ||
| ↓ tumor mass |
5.3. Curcumin
5.4. Carnosol
5.5. Quercetin and Rutin
5.6. Genistein, Daidzein and Morin
6. Other Antioxidants with Therapeutic Potential for Cancer Cachexia
6.1. Ursolic Acid
6.2. Sulforaphane
| Target | Experimental Setting | SFN Treatment | Findings | References |
|---|---|---|---|---|
| Attenuation of muscle atrophy via regulation of Akt/FoxO1 | In vitro C2C12 myotubes | Dexamethasone (5 μM) and SFN (5 μM) for 24 h | ↑ Akt phosphorylation | [40] |
| ↑ protein synthesis | ||||
| ↑ MyoD | ||||
| ↓ atrogin-1 via FoxO1 signaling | ||||
| ↓ myocyte viability and no myotube death at 20 μM of SFN | ||||
| Nrf2/ARE signaling pathway vs. menadione-induced oxidative stress | In vitro Dystrophin knock-down muscle cells, C2 DysKD myotube | Myotubes were pre-treated with SFN (5 μM) for 5 h prior to menadione (20 μM) exposure | ↑ Nrf2 translocation into nucleus | [39] |
| ↑ phosphorylation of Akt and Nrf2 | ||||
| ↑ expression of total Akt | ||||
| Repression of myostatin and myostatin related signaling pathway | In vitro porcine satellite cells | Myoblasts were treated with 5, 10 or 15 μM SFN for 48 h | ↓ myostatin gene expression | [115] |
| ↑ Smad7, Smurf1 gene expression at all doses | ||||
| ↓ myostatin signaling pathway | ||||
| ↓ MyoD | ||||
| ↓ cell proliferation at 15 μM | ||||
| ↓ Caspase 3 and 9 activities at 10 μM | ||||
| ↑ cell viability at 5 μM | ||||
| Modulation of CX3CL1/CX3CR1 axis and inflammation against palmitic acid-induced cell injury | In vitro C2C12 myotubes | C2C12 cells were pre-treated with SFN (5 µM) before exposure to 750 µM palmitate for 24 h | ↑ cell viability | [118] |
| ↓ IL-6, TNF-α | ||||
| ↑ expression of Nrf2, HMOX1 | ||||
| ↓ palmitic acid-induced ROS level | ||||
| Attenuation of serum starvation-induced and oxidative stress-induced muscle atrophy via Nrf2 activation | In vitro C2C12 myotubes | C2C12 cells were pre-treated for 3 h with SFN (5 µM) before exposure to 20 µM menadione, or culturing without serum for 3 h | In both conditions: Prevention against muscle damage | [116] |
| ↓ dichlorofluorescin diacetate (DCFDA) intensity, a general ROS indicator | ||||
| Attenuation of dystrophic pathology and muscle inflammation via Nrf2 pathway | In vivo 4-week-old mdx mice | 2 mg/kg/day SFN via oral gavage for either 4 or 8 weeks | ↑ body weight, ↓ gastric and myocardial muscle hypertrophy | [109,119] |
| ↓ creatine kinase (CK) and lactate dehydrogenase (LDH) levels | ||||
| Activated Nrf2/ARE pathway | ||||
| Attenuation of muscle fibrosis via Nrf2 pathway | In vivo 3-month-old mdx mice | 2 mg/kg/day SFN via oral gavage for 3 months | ↓ CK and LDH levels | [43] |
| ↓ MDA, GSH/oxidized glutathione (GSSG) ratio | ||||
| Anti-fibrosis function in liver and lungs | ||||
| ↓ p-smad2/3 = suppress profibrogenic gene | ||||
| ↓ IL-6, CD45, TNF-α = ↓ inflammatory | ||||
| Inhibition of TGF-β/Smad signaling | ||||
| Enhancement of exercise endurance capacity via Nrf2 activation | In vivo 13-week-old male mice: Nrf2+/+ or Nrf2−/− | 25 mg/kg SFN via i.p. injection 4 times in 3 days | ↑ HMOX1, NQO1, gamma-glutamylcysteine synthetase (γ-GCS), and catalase | [120] |
| ↓ CK and LDH levels | ||||
| ↓ GSH/GSSG ratio, thiobarbituric acid reactive substances (TBARS) | ||||
| Activated Nrf2 to ↓ muscle fatigue | ||||
| Protection against muscle damage induced by exhaustive exercise | In vivo 4-month-old male Wistar rats | 25 mg/kg/day SFN via i.p. injection for 3 days prior to intensive exercise | ↓ CK and LDH levels | [121] |
| ↑ glutathione reductase (GR), GST, NQO1 | ||||
| Activated Nrf2/ARE pathway | ||||
| Prevention of age-associated muscle dysfunction via Nrf2 signaling pathway | In vivo 21/22-month-old male C57BL/6 mice | 442.5 mg/kg D, L-SFN supplemented diet for 12 weeks | ↑ survival, no body weight change | [122] |
| ↑ exercise capacity | ||||
| ↑ MyoD, paired box 7 in satellite cell-derived myoblasts isolated from extensor digitorum longus muscles of lower hindlimbs | ||||
| ↓ myostatin, 8OHdG (oxidation marker), apoptosis | ||||
| ↑ Catalase, SOD1, Gpx1, GSTA4 and Nrf2 mRNA | ||||
| Activated Nrf2/ARE pathway | ||||
| Protection against type 2 diabetes related muscle dysfunction | In vivo 20-week-old db/db type 2 diabatic mice | daily 0.5 mg/kg SFN via i.p. injection for one month | ↑ grip strength | [123] |
| ↑ lean mass, ↓ fat mass | ||||
| Restored muscle fiber structure | ||||
| ↓P65, TNF-α, plasminogen activator inhibitor-1, TGF-β1, Caspase 3 and Caspase 8 | ||||
| Activated Nrf2/ARE pathway |
6.3. Honokiol and Magnolol
6.4. Pomegranate Extract
6.5. Ellagic Acid and Urolithin A, B
6.6. Other Polyphenols
7. Dual-Function of Antioxidants in Cancer Cachexia
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Fearon, K.C.; Voss, A.C.; Hustead, D.S.; Cancer Cachexia Study, G. Definition of cancer cachexia: Effect of weight loss, reduced food intake, and systemic inflammation on functional status and prognosis. Am. J. Clin. Nutr. 2006, 83, 1345–1350. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Keshavarz-Fathi, M.; Baracos, V.; Arends, J.; Mahmoudi, M.; Rezaei, N. Cancer cachexia: Diagnosis, assessment, and treatment. Crit. Rev. Oncol. Hematol. 2018, 127, 91–104. [Google Scholar] [CrossRef]
- Acharyya, S.; Guttridge, D.C. Cancer cachexia signaling pathways continue to emerge yet much still points to the proteasome. Clin. Cancer Res. 2007, 13, 1356–1361. [Google Scholar] [CrossRef] [Green Version]
- Fearon, K.C.; Glass, D.J.; Guttridge, D.C. Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell Metab. 2012, 16, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Argiles, J.M.; Busquets, S.; Felipe, A.; Lopez-Soriano, F.J. Molecular mechanisms involved in muscle wasting in cancer and ageing: Cachexia versus sarcopenia. Int. J. Biochem. Cell Biol. 2005, 37, 1084–1104. [Google Scholar] [CrossRef]
- Aoyagi, T.; Terracina, K.P.; Raza, A.; Matsubara, H.; Takabe, K. Cancer cachexia, mechanism and treatment. World J. Gastrointest. Oncol. 2015, 7, 17–29. [Google Scholar] [CrossRef]
- Mantovani, G.; Madeddu, C. Cancer cachexia: Medical management. Support Care Cancer 2010, 18, 1–9. [Google Scholar] [CrossRef]
- Fearon, K.; Arends, J.; Baracos, V. Understanding the mechanisms and treatment options in cancer cachexia. Nat. Rev. Clin. Oncol. 2013, 10, 90–99. [Google Scholar] [CrossRef]
- Powers, S.K.; Lynch, G.S.; Murphy, K.T.; Reid, M.B.; Zijdewind, I. Disease-Induced Skeletal Muscle Atrophy and Fatigue. Med. Sci. Sports Exerc. 2016, 48, 2307–2319. [Google Scholar] [CrossRef] [PubMed]
- Eley, H.L.; Tisdale, M.J. Skeletal muscle atrophy, a link between depression of protein synthesis and increase in degradation. J. Biol. Chem. 2007, 282, 7087–7097. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, S.J.; Ogden, J.; Ramjee, G.; Rund, J. Contribution of elevated protein turnover and anorexia to cachexia in patients with hepatocellular carcinoma. Cancer Res. 1990, 50, 1226–1230. [Google Scholar] [CrossRef] [Green Version]
- Acharyya, S.; Butchbach, M.E.; Sahenk, Z.; Wang, H.; Saji, M.; Carathers, M.; Ringel, M.D.; Skipworth, R.J.; Fearon, K.C.; Hollingsworth, M.A.; et al. Dystrophin glycoprotein complex dysfunction: A regulatory link between muscular dystrophy and cancer cachexia. Cancer Cell 2005, 8, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Damrauer, J.S.; Stadler, M.E.; Acharyya, S.; Baldwin, A.S.; Couch, M.E.; Guttridge, D.C. Chemotherapy-induced muscle wasting: Association with NF-kappaB and cancer cachexia. Eur. J. Transl. Myol. 2018, 28, 7590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrigo, J.; Elorza, A.A.; Riedel, C.A.; Vilos, C.; Simon, F.; Cabrera, D.; Estrada, L.; Cabello-Verrugio, C. Role of Oxidative Stress as Key Regulator of Muscle Wasting during Cachexia. Oxid. Med. Cell Longev. 2018, 2018, 2063179. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, E.; de la Puente, B.; Busquets, S.; Lopez-Soriano, F.J.; Gea, J.; Argiles, J.M. Both oxidative and nitrosative stress are associated with muscle wasting in tumour-bearing rats. FEBS Lett. 2005, 579, 1646–1652. [Google Scholar] [CrossRef] [Green Version]
- Gomes-Marcondes, M.C.; Tisdale, M.J. Induction of protein catabolism and the ubiquitin-proteasome pathway by mild oxidative stress. Cancer Lett. 2002, 180, 69–74. [Google Scholar] [CrossRef]
- White, J.P.; Puppa, M.J.; Sato, S.; Gao, S.; Price, R.L.; Baynes, J.W.; Kostek, M.C.; Matesic, L.E.; Carson, J.A. IL-6 regulation on skeletal muscle mitochondrial remodeling during cancer cachexia in the ApcMin/+ mouse. Skelet. Muscle 2012, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Bashan, N.; Kovsan, J.; Kachko, I.; Ovadia, H.; Rudich, A. Positive and negative regulation of insulin signaling by reactive oxygen and nitrogen species. Physiol. Rev. 2009, 89, 27–71. [Google Scholar] [CrossRef] [Green Version]
- Pigna, E.; Berardi, E.; Aulino, P.; Rizzuto, E.; Zampieri, S.; Carraro, U.; Kern, H.; Merigliano, S.; Gruppo, M.; Mericskay, M.; et al. Aerobic Exercise and Pharmacological Treatments Counteract Cachexia by Modulating Autophagy in Colon Cancer. Sci. Rep. 2016, 6, 26991. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.; Moldawer, L.L.; Marano, M.; Wei, H.; Barber, A.; Manogue, K.; Tracey, K.J.; Kuo, G.; Fischman, D.A.; Cerami, A.; et al. Cachectin/TNF or IL-1 alpha induces cachexia with redistribution of body proteins. Am. J. Physiol. 1989, 256, R659–R665. [Google Scholar] [CrossRef]
- Strassmann, G.; Fong, M.; Kenney, J.S.; Jacob, C.O. Evidence for the involvement of interleukin 6 in experimental cancer cachexia. J. Clin. Investig. 1992, 89, 1681–1684. [Google Scholar] [CrossRef] [Green Version]
- Prado, B.L.; Qian, Y. Anti-cytokines in the treatment of cancer cachexia. Ann. Palliat. Med. 2019, 8, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Osthoff, K.; Bakker, A.; Vanhaesebroeck, B.; Beyaert, R.; Jacob, W.A.; Fiers, W. Cytotoxic activity of tumor necrosis factor is mediated by early damage of mitochondrial functions. Evidence for the involvement of mitochondrial radical generation. J. Biol. Chem. 1992, 267, 5317–5323. [Google Scholar] [CrossRef]
- Mantovani, G.; Maccio, A.; Madeddu, C.; Gramignano, G.; Lusso, M.R.; Serpe, R.; Massa, E.; Astara, G.; Deiana, L. A phase II study with antioxidants, both in the diet and supplemented, pharmaconutritional support, progestagen, and anti-cyclooxygenase-2 showing efficacy and safety in patients with cancer-related anorexia/cachexia and oxidative stress. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1030–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puig-Vilanova, E.; Rodriguez, D.A.; Lloreta, J.; Ausin, P.; Pascual-Guardia, S.; Broquetas, J.; Roca, J.; Gea, J.; Barreiro, E. Oxidative stress, redox signaling pathways, and autophagy in cachectic muscles of male patients with advanced COPD and lung cancer. Free Radic. Biol. Med. 2015, 79, 91–108. [Google Scholar] [CrossRef]
- Mantovani, G.; Maccio, A.; Madeddu, C.; Mura, L.; Gramignano, G.; Lusso, M.R.; Massa, E.; Mocci, M.; Serpe, R. Antioxidant agents are effective in inducing lymphocyte progression through cell cycle in advanced cancer patients: Assessment of the most important laboratory indexes of cachexia and oxidative stress. J. Mol. Med. 2003, 81, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.A.; Hassan, S.; Harvey, K.A.; Rasool, T.; Das, T.; Mukerji, P.; DeMichele, S. Attenuation of proteolysis and muscle wasting by curcumin c3 complex in MAC16 colon tumour-bearing mice. Br. J. Nutr. 2009, 102, 967–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.P.; Schwartz, R.J.; Waddell, I.D.; Holloway, B.R.; Reid, M.B. Skeletal muscle myocytes undergo protein loss and reactive oxygen-mediated NF-kappaB activation in response.e.e to tumor necrosis factor alpha. FASEB J. 1998, 12, 871–880. [Google Scholar] [CrossRef]
- Baldwin, A.S., Jr. The NF-kappa B and I kappa B proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, A.S. Control of oncogenesis and cancer therapy resistance by the transcription factor NF-kappaB. J. Clin. Investig. 2001, 107, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Lv, Y.; Zhang, W.; Chai, X.; Feng, L.; Fang, Y.; Liu, X.; Zhang, X. Pyrrolidine Dithiocarbamate (PDTC) Attenuates Cancer Cachexia by Affecting Muscle Atrophy and Fat Lipolysis. Front. Pharm. 2017, 8, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyke, S.M.; Russell, S.T.; Tisdale, M.J. Induction of proteasome expression in skeletal muscle is attenuated by inhibitors of NF-kappaB activation. Br. J. Cancer 2004, 91, 1742–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Lai, Y.J.; Chan, Y.L.; Li, T.L.; Wu, C.J. Epigallocatechin-3-gallate effectively attenuates skeletal muscle atrophy caused by cancer cachexia. Cancer Lett. 2011, 305, 40–49. [Google Scholar] [CrossRef]
- Schmid, H.H. Reasons for low incidence of peptic ulcer in pregnancy. Zent. Gynakol. 1950, 72, 1839–1840. [Google Scholar]
- Huang, Y.; Li, W.; Su, Z.Y.; Kong, A.N. The complexity of the Nrf2 pathway: Beyond the antioxidant response. J. Nutr. Biochem. 2015, 26, 1401–1413. [Google Scholar] [CrossRef]
- Kensler, T.W.; Egner, P.A.; Agyeman, A.S.; Visvanathan, K.; Groopman, J.D.; Chen, J.G.; Chen, T.Y.; Fahey, J.W.; Talalay, P. Keap1-nrf2 signaling: A target for cancer prevention by sulforaphane. Top. Curr. Chem. 2013, 329, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Russo, M.; Spagnuolo, C.; Russo, G.L.; Skalicka-Wozniak, K.; Daglia, M.; Sobarzo-Sanchez, E.; Nabavi, S.F.; Nabavi, S.M. Nrf2 targeting by sulforaphane: A potential therapy for cancer treatment. Crit. Rev. Food Sci. Nutr. 2018, 58, 1391–1405. [Google Scholar] [CrossRef]
- Choi, S.J.; Kim, H.S. Deregulation of Nrf2/ARE signaling pathway causes susceptibility of dystrophin-deficient myotubes to menadione-induced oxidative stress. Exp. Cell Res. 2018, 364, 224–233. [Google Scholar] [CrossRef]
- Son, Y.H.; Jang, E.J.; Kim, Y.W.; Lee, J.H. Sulforaphane prevents dexamethasone-induced muscle atrophy via regulation of the Akt/Foxo1 axis in C2C12 myotubes. Biomed. Pharm. 2017, 95, 1486–1492. [Google Scholar] [CrossRef]
- Dozio, E.; Ruscica, M.; Passafaro, L.; Dogliotti, G.; Steffani, L.; Marthyn, P.; Pagani, A.; Demartini, G.; Esposti, D.; Fraschini, F.; et al. The natural antioxidant alpha-lipoic acid induces p27(Kip1)-dependent cell cycle arrest and apoptosis in MCF-7 human breast cancer cells. Eur. J. Pharm. 2010, 641, 29–34. [Google Scholar] [CrossRef]
- Zhang, D.D.; Hannink, M. Distinct cysteine residues in Keap1 are required for Keap1-dependent ubiquitination of Nrf2 and for stabilization of Nrf2 by chemopreventive agents and oxidative stress. Mol. Cell Biol. 2003, 23, 8137–8151. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Li, S.; Li, D. Sulforaphane mitigates muscle fibrosis in mdx mice via Nrf2-mediated inhibition of TGF-beta/Smad signaling. J. Appl. Physiol. 2016, 120, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Bessler, H.; Djaldetti, M. Broccoli and human health: Immunomodulatory effect of sulforaphane in a model of colon cancer. Int. J. Food Sci. Nutr. 2018, 69, 946–953. [Google Scholar] [CrossRef]
- Sporn, M.B.; Liby, K.T. NRF2 and cancer: The good, the bad and the importance of context. Nat. Rev. Cancer 2012, 12, 564–571. [Google Scholar] [CrossRef]
- Moon, E.J.; Giaccia, A. Dual roles of NRF2 in tumor prevention and progression: Possible implications in cancer treatment. Free Radic. Biol. Med. 2015, 79, 292–299. [Google Scholar] [CrossRef] [Green Version]
- Shibata, T.; Kokubu, A.; Gotoh, M.; Ojima, H.; Ohta, T.; Yamamoto, M.; Hirohashi, S. Genetic alteration of Keap1 confers constitutive Nrf2 activation and resistance to chemotherapy in gallbladder cancer. Gastroenterology 2008, 135, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Bodas, M.; Wakabayashi, N.; Bunz, F.; Biswal, S. Gain of Nrf2 function in non-small-cell lung cancer cells confers radioresistance. Antioxid. Redox. Signal. 2010, 13, 1627–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, H.; Moriguchi, T.; Takai, J.; Ebina, M.; Yamamoto, M. Nrf2 prevents initiation but accelerates progression through the Kras signaling pathway during lung carcinogenesis. Cancer Res. 2013, 73, 4158–4168. [Google Scholar] [CrossRef] [Green Version]
- Salucci, S.; Falcieri, E. Polyphenols and their potential role in preventing skeletal muscle atrophy. Nutr. Res. 2020, 74, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Yang, C.S.; Wang, X. Green tea and cancer prevention. Nutr. Cancer 2010, 62, 931–937. [Google Scholar] [CrossRef]
- Yang, C.S.; Lambert, J.D.; Hou, Z.; Ju, J.; Lu, G.; Hao, X. Molecular targets for the cancer preventive activity of tea polyphenols. Mol. Carcinog. 2006, 45, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Carbo, N.; Costelli, P.; Baccino, F.M.; Lopez-Soriano, F.J.; Argiles, J.M. Resveratrol, a natural product present in wine, decreases tumour growth in a rat tumour model. Biochem. Biophys. Res. Commun. 1999, 254, 739–743. [Google Scholar] [CrossRef]
- Rai, G.; Mishra, S.; Suman, S.; Shukla, Y. Resveratrol improves the anticancer effects of doxorubicin in vitro and in vivo models: A mechanistic insight. Phytomedicine 2016, 23, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Shadfar, S.; Couch, M.E.; McKinney, K.A.; Weinstein, L.J.; Yin, X.; Rodriguez, J.E.; Guttridge, D.C.; Willis, M. Oral resveratrol therapy inhibits cancer-induced skeletal muscle and cardiac atrophy in vivo. Nutr. Cancer 2011, 63, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Busquets, S.; Fuster, G.; Ametller, E.; Olivan, M.; Figueras, M.; Costelli, P.; Carbo, N.; Argiles, J.M.; Lopez-Soriano, F.J. Resveratrol does not ameliorate muscle wasting in different types of cancer cachexia models. Clin. Nutr. 2007, 26, 239–244. [Google Scholar] [CrossRef]
- Penedo-Vázquez, A.; Duran, X.; Mateu, J.; López-Postigo, A.; Barreiro, E. Curcumin and Resveratrol Improve Muscle Function and Structure through Attenuation of Proteolytic Markers in Experimental Cancer-Induced Cachexia. Molecules 2021, 26, 4904. [Google Scholar] [CrossRef]
- Busquets, S.l.; Carbó, N.; Almendro, V.; Quiles, M.a.T.; López-Soriano, F.J.; Argilés, J.M. Curcumin, a natural product present in turmeric, decreases tumor growth but does not behave as an anticachectic compound in a rat model. Cancer Lett. 2001, 167, 33–38. [Google Scholar] [CrossRef]
- Lu, S.; Li, Y.; Shen, Q.; Zhang, W.; Gu, X.; Ma, M.; Li, Y.; Zhang, L.; Liu, X.; Zhang, X. Carnosol and its analogues attenuate muscle atrophy and fat lipolysis induced by cancer cachexia. J. Cachexia Sarcopenia Muscle 2021, 12, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, K.T.; Enos, R.T.; Narsale, A.A.; Puppa, M.J.; Davis, J.M.; Murphy, E.A.; Carson, J.A. Quercetin Supplementation Attenuates the Progression of Cancer Cachexia in ApcMin/+Mice. J. Nutr. 2014, 144, 868–875. [Google Scholar] [CrossRef] [Green Version]
- Levolger, S.; van den Engel, S.; Ambagtsheer, G.; Ijzermans, J.N.M.; de Bruin, R.W.F. Quercetin supplementation attenuates muscle wasting in cancer-associated cachexia in mice. Nutr. Healthy Aging 2021, 6, 35–47. [Google Scholar] [CrossRef]
- Gil da Costa, R.M.; Aragão, S.; Moutinho, M.; Alvarado, A.; Carmo, D.; Casaca, F.; Silva, S.; Ribeiro, J.; Sousa, H.; Ferreira, R.; et al. HPV16 induces a wasting syndrome in transgenic mice: Amelioration by dietary polyphenols via NF-κB inhibition. Life Sci. 2017, 169, 11–19. [Google Scholar] [CrossRef]
- Hirasaka, K.; Saito, S.; Yamaguchi, S.; Miyazaki, R.; Wang, Y.; Haruna, M.; Taniyama, S.; Higashitani, A.; Terao, J.; Nikawa, T.; et al. Dietary Supplementation with Isoflavones Prevents Muscle Wasting in Tumor-Bearing Mice. J. Nutr. Sci. Vitaminol. 2016, 62, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, T.; Saitoh, K.; Sun, L.; Wang, Y.; Taniyama, S.; Yamaguchi, K.; Uchida, T.; Ohkubo, T.; Higashitani, A.; Nikawa, T.; et al. Morin suppresses cachexia-induced muscle wasti.ing by binding to ribosomal protein S10 in carcinoma cells. Biochem. Biophys. Res. Commun. 2018, 506, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.T.; Yin, Y.; Yang, Y.J.; Lv, P.J.; Shi, Y.; Lu, L.; Wei, L.B. Resveratrol prevents TNF-alpha-induced muscle atrophy via regulation of Akt/mTOR/FoxO1 signaling in C2C12 myotubes. Int. Immunopharmacol. 2014, 19, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Menzies, K.J.; Singh, K.; Saleem, A.; Hood, D.A. Sirtuin 1-mediated effects of exercise and resveratrol on mitochondrial biogenesis. J. Biol. Chem. 2013, 288, 6968–6979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, A.M.; Candau, R.B.; Bernardi, H. FoxO transcription factors: Their roles in the maintenance of skeletal muscle homeostasis. Cell Mol. Life Sci. 2014, 71, 1657–1671. [Google Scholar] [CrossRef]
- Quan-Jun, Y.; Jun, B.; Li-Li, W.; Yong-Long, H.; Bin, L.; Qi, Y.; Yan, L.; Cheng, G.; Gen-Jin, Y. NMR-based metabolomics reveals distinct pathways mediated by curcumin in cachexia mice bearing CT26 tumor. RSC Adv. 2015, 5, 11766–11775. [Google Scholar] [CrossRef]
- Dhillon, N.; Aggarwal, B.B.; Newman, R.A.; Wolff, R.A.; Kunnumakkara, A.B.; Abbruzzese, J.L.; Ng, C.S.; Badmaev, V.; Kurzrock, R. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin. Cancer Res. 2008, 14, 4491–4499. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.A.; Euden, S.A.; Platton, S.L.; Cooke, D.N.; Shafayat, A.; Hewitt, H.R.; Marczylo, T.H.; Morgan, B.; Hemingway, D.; Plummer, S.M.; et al. Phase I clinical trial of oral curcumin: Biomarkers of systemic activity and compliance. Clin. Cancer Res. 2004, 10, 6847–6854. [Google Scholar] [CrossRef] [Green Version]
- Shehzad, A.; Wahid, F.; Lee, Y.S. Curcumin in cancer chemoprevention: Molecular targets, pharmacokinetics, bioavailability, and clinical trials. Arch. Pharm. 2010, 343, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Pala, R.; Tuzcu, M.; Ozdemir, O.; Orhan, C.; Sahin, N.; Juturu, V. Curcumin prevents muscle damage by regulating NF-kappaB and Nrf2 pathways and improves performance: An in vivo model. J. Inflamm. Res. 2016, 9, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, P.; Sharma, Y.K.; Sharma, S.; Singh, S.N.; Suryakumar, G. High altitude mediated skeletal muscle atrophy: Protective role of curcumin. Biochimie 2019, 156, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Assi, M.; Rebillard, A. The Janus-Faced Role of Antioxidants in Cancer Cachexia: New Insights on the Established Concepts. Oxid. Med. Cell Longev. 2016, 2016, 9579868. [Google Scholar] [CrossRef] [Green Version]
- Loussouarn, M.; Krieger-Liszkay, A.; Svilar, L.; Bily, A.; Birtic, S.; Havaux, M. Carnosic Acid and Carnosol, Two Major Antioxidants of Rosemary, Act through Different Mechanisms. Plant. Physiol. 2017, 175, 1381–1394. [Google Scholar] [CrossRef] [Green Version]
- Lo, A.H.; Liang, Y.C.; Lin-Shiau, S.Y.; Ho, C.T.; Lin, J.K. Carnosol, an antioxidant in rosemary, suppresses inducible nitric oxide synthase through down-regulating nuclear factor-kappaB in mouse macrophages. Carcinogenesis 2002, 23, 983–991. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.; Wu, S. The Mechanisms of Carnosol in Chemoprevention of Ultraviolet B-Light-Induced Non-Melanoma Skin Cancer Formation. Sci. Rep. 2018, 8, 3574. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.E.; Carothers, A.M.; Weyant, M.J.; Redston, M.; Bertagnolli, M.M. Carnosol inhibits β-catenin tyrosine phosphorylation and prevents adenoma formation in the C57BL/6J/Min/+(Min/+) mouse. Cancer Res. 2005, 65, 1097–1104. [Google Scholar]
- Alsamri, H.; El Hasasna, H.; Al Dhaheri, Y.; Eid, A.H.; Attoub, S.; Iratni, R. Carnosol, a Natural Polyphenol, Inhibits Migration, Metastasis, and Tumor Growth of Breast Cancer via a RO.OS-Dependent Proteasome Degradation of STAT3. Front. Oncol. 2019, 9, 743. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Yu, L.; Lu, P.; Wei, Y.; Wang, X.; Chen, L. Evaluate the inhibition of cytochrome P450 1A1 for enhancing breast cancer chemotherapy with a turn-on fluorescent probe. Sens. Actuators B Chem. 2021, 344, 130233. [Google Scholar] [CrossRef]
- Quercetin Reduces Tumor Necrosis Factor Alpha-Induced Muscle Atrophy by Upregulation of Heme Oxygenase-1. J. Med. Food 2018, 21, 551–559. [CrossRef] [PubMed]
- Camargo, C.A.; da Silva, M.E.F.; da Silva, R.A.; Justo, G.Z.; Gomes-Marcondes, M.C.C.; Aoyama, H. Inhibition of tumor growth by quercetin with increase of survival and prevention of cachexia in Walker 256 tumor-bearing rats. Biochem. Biophys. Res. Commun. 2011, 406, 638–642. [Google Scholar] [CrossRef]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.R.; Moselhy, A.A.A.; El-Mleeh, A.; Aljuaydi, S.H.; Ismail, T.A.; Di Cerbo, A.; Crescenzo, G.; Abou-Zeid, S.M. Quercetin Alleviates the Immunotoxic Impact Mediated by Oxidative Stress and Inflammation Induced by Doxorubicin Exposure in Rats. Antioxidants 2021, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Morand, C.; Demigné, C.; Texier, O.; Régérat, F.; Rémésy, C. Bioavailability of rutin and quercetin in rats. FEBS Lett. 1997, 409, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Moutinho, M.S.S.; Aragão, S.; Carmo, D.; Casaca, F.; Silva, S.; Ribeiro, J.; Sousa, H.; Pires, I.; Queiroga, F.; Colaço, B.; et al. Curcumin and Rutin Down-regulate Cyclooxygenase-2 and Reduce Tumor-associated Inflammation in HPV16-Transgenic Mice. Anticancer Res. 2018, 38, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Satari, A.; Ghasemi, S.; Habtemariam, S.; Asgharian, S.; Lorigooini, Z. Rutin: A Flavonoid as an Effective Sensitizer for Anticancer Therapy; Insights into Multifaceted Mechanisms and Applicability for Combination Therapy. Evid. -Based Complementary Altern. Med. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-J.; Xie, M.-Y.; Kluxen, F.M.; Diel, P. Genistein modulates the anti-tumor activity of cisplatin in MCF-7 breast and HT-29 colon cancer cells. Arch. Toxicol. 2014, 88, 625–635. [Google Scholar] [CrossRef]
- Qiu, S.; Jiang, C. Soy and isoflavones consumption and breast cancer survival and recurrence: A systematic review and meta-analysis. Eur. J. Nutr. 2019, 58, 3079–3090. [Google Scholar] [CrossRef]
- Ma, X.; Yu, X.; Min, J.; Chen, X.; Liu, R.; Cui, X.; Cheng, J.; Xie, M.; Diel, P.; Hu, X. Genistein interferes with antitumor effects of cisplatin in an ovariectomized breast cancer xenograft tumor model. Toxicol. Lett. 2022, 355, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, M.U.; Shahzadi, S.; Zahra, Z.; Samad, A.; Ehsan, N.; Afsar, T.; Almajwal, A.; Al-disi, D.; Abulmeaty, M.; Razak, S. Ameliorative effects of morin on cisplatin-induced toxicity in renal mitochondria isolated from rats. J. King Saud Univ. Sci. 2021, 33, 101655. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Dai, X.; Kumar, A.P.; Tan, B.K.; Sethi, G.; Bishayee, A. Ursolic acid in cancer prevention and treatment: Molecular targets, pharmacokinetics and clinical studies. Biochem. Pharm. 2013, 85, 1579–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, H.S.; Seo, D.Y.; Chung, Y.M.; Kim, D.H.; Lee, S.J.; Lee, S.R.; Kwak, H.B.; Kim, T.N.; Kim, M.; Oh, K.M.; et al. Ursolic acid supplementation decreases markers of skeletal muscle damage during resistance training in resistance-trained men: A pilot study. Korean J. Physiol. Pharm. 2017, 21, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, M.K.; Rajendran, P.; Li, F.; Nema, T.; Vali, S.; Abbasi, T.; Kapoor, S.; Sharma, A.; Kumar, A.P.; Ho, P.C.; et al. Ursolic acid inhibits multiple cell survival pathways leading to suppression of growth of prostate cancer xenograft in nude mice. J. Mol. Med. 2011, 89, 713–727. [Google Scholar] [CrossRef]
- Lin, J.; Chen, Y.; Wei, L.; Hong, Z.; Sferra, T.J.; Peng, J. Ursolic acid inhibits colorectal cancer angiogenesis through suppression of multiple signaling pathways. Int. J. Oncol. 2013, 43, 1666–1674. [Google Scholar] [CrossRef] [Green Version]
- Kunkel, S.D.; Suneja, M.; Ebert, S.M.; Bongers, K.S.; Fox, D.K.; Malmberg, S.E.; Alipour, F.; Shields, R.K.; Adams, C.M. mRNA expression signatures of human skeletal muscle atrophy identify a natural compound that increases muscle mass. Cell Metab. 2011, 13, 627–638. [Google Scholar] [CrossRef] [Green Version]
- Rathor, R.; Agrawal, A.; Kumar, R.; Suryakumar, G.; Singh, S.N. Ursolic acid ameliorates hypobaric hypoxia-induced skeletal muscle protein loss via upregulating Akt pathway: An experimental study using rat model. IUBMB Life 2021, 73, 375–389. [Google Scholar] [CrossRef]
- Kunkel, S.D.; Elmore, C.J.; Bongers, K.S.; Ebert, S.M.; Fox, D.K.; Dyle, M.C.; Bullard, S.A.; Adams, C.M. Ursolic acid increases skeletal muscle and brown fat and decreases diet-induced obesity, glucose intolerance and fatty liver disease. PLoS ONE 2012, 7, e39332. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, R.; Sato, K.; Higashida, K.; Nakazato, K.; Fujita, S. Ursolic acid stimulates mTORC1 signaling after resistance exercise in rat skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E760–E765. [Google Scholar] [CrossRef] [Green Version]
- Bakhtiari, N.; Hosseinkhani, S.; Soleimani, M.; Hemmati, R.; Noori-Zadeh, A.; Javan, M.; Tashakor, A. Short-term ursolic acid promotes skeletal muscle rejuvenation through enhancing of SIRT1 expression and satellite cells proliferation. Biomed. Pharm. 2016, 78, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Bakhtiari, N.; Hosseinkhani, S.; Tashakor, A.; Hemmati, R. Ursolic acid ameliorates aging-metabolic phenotype through promoting of skeletal muscle rejuvenation. Med. Hypotheses 2015, 85, 1–6. [Google Scholar] [CrossRef]
- Zhang, Y.; Talalay, P.; Cho, C.G.; Posner, G.H. A major inducer of anticarcinogenic protective enzymes from broccoli: Isolation and elucidation of structure. Proc. Natl. Acad. Sci. USA 1992, 89, 2399–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahey, J.W.; Zhang, Y.; Talalay, P. Broccoli sprouts: An exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc. Natl. Acad. Sci. USA 1997, 94, 10367–10372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Beltran, C.E.; Calderon-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective effect of sulforaphane against oxidative stress: Recent advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef]
- Hanlon, N.; Coldham, N.; Gielbert, A.; Kuhnert, N.; Sauer, M.J.; King, L.J.; Ioannides, C. Absolute bioavailability and dose-dependent pharmacokinetic behaviour of dietary doses of the chemopreventive isothiocyanate sulforaphane in rat. Br. J. Nutr. 2008, 99, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Houghton, C.A. Sulforaphane: Its “Coming of Age” as a Clinically Relevant Nutraceutical in the Prevention and Treatment of Chronic Disease. Oxid. Med. Cell Longev. 2019, 2019, 2716870. [Google Scholar] [CrossRef] [Green Version]
- Thimmulappa, R.K.; Mai, K.H.; Srisuma, S.; Kensler, T.W.; Yamamoto, M.; Biswal, S. Identification of Nrf2-regulated genes induced by the chemopreventive agent sulforaphane by oligonucleotide microarray. Cancer Res. 2002, 62, 5196–5203. [Google Scholar]
- Sun, C.; Yang, C.; Xue, R.; Li, S.; Zhang, T.; Pan, L.; Ma, X.; Wang, L.; Li, D. Sulforaphane alleviates muscular dystrophy in mdx mice by activation of Nrf2. J. Appl. Physiol. 2015, 118, 224–237. [Google Scholar] [CrossRef]
- Kanematsu, S.; Yoshizawa, K.; Uehara, N.; Miki, H.; Sasaki, T.; Kuro, M.; Lai, Y.C.; Kimura, A.; Yuri, T.; Tsubura, A. Sulforaphane inhibits the growth of KPL-1 human breast cancer cells in vitro and suppresses the growth and metastasis of orthotopically transplanted KPL-1 cells in female athymic mice. Oncol. Rep. 2011, 26, 603–608. [Google Scholar] [CrossRef]
- Wang, D.X.; Zou, Y.J.; Zhuang, X.B.; Chen, S.X.; Lin, Y.; Li, W.L.; Lin, J.J.; Lin, Z.Q. Sulforaphane suppresses EMT and metastasis in human lung cancer through miR-616-5p-mediated GSK3beta/beta-catenin signaling pathways. Acta Pharm. Sin. 2017, 38, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Su, Z.Y.; Khor, T.O.; Shu, L.; Kong, A.N. Sulforaphane enhances Nrf2 expression in prostate cancer TRAMP C1 cells through epigenetic regulation. Biochem. Pharm. 2013, 85, 1398–1404. [Google Scholar] [CrossRef] [Green Version]
- Barve, A.; Khor, T.O.; Nair, S.; Reuhl, K.; Suh, N.; Reddy, B.; Newmark, H.; Kong, A.N. Gamma-tocopherol-enriched mixed tocopherol diet inhibits prostate carcinogenesis in TRAMP mice. Int. J. Cancer 2009, 124, 1693–1699. [Google Scholar] [CrossRef] [Green Version]
- Traka, M.H.; Melchini, A.; Coode-Bate, J.; Al Kadhi, O.; Saha, S.; Defernez, M.; Troncoso-Rey, P.; Kibblewhite, H.; O’Neill, C.M.; Bernuzzi, F.; et al. Transcriptional changes in prostate of men on active surveillance after a 12-mo glucoraphanin-rich broccoli intervention-results from the Effect of Sulforaphane on prostate CAncer PrEvention (ESCAPE) randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 1133–1144. [Google Scholar] [CrossRef]
- Fan, H.; Zhang, R.; Tesfaye, D.; Tholen, E.; Looft, C.; Holker, M.; Schellander, K.; Cinar, M.U. Sulforaphane causes a major epigenetic repression of myostatin in porcine satellite cells. Epigenetics 2012, 7, 1379–1390. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.Y.; Kim, D.J.; Kim, H.S. Sulforaphane ameliorates serum starvation-induced muscle atrophy via activation of the Nrf2 pathway in cultured C2C12 cells. Cell Biol. Int. 2020, 44, 1831–1839. [Google Scholar] [CrossRef]
- Whitman, S.A.; Long, M.; Wondrak, G.T.; Zheng, H.; Zhang, D.D. Nrf2 modulates contractile and metabolic properties of skeletal muscle in streptozotocin-induced diabetic atrophy. Exp. Cell Res. 2013, 319, 2673–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faridvand, Y.; Haddadi, P.; Nejabati, H.R.; Ghaffari, S.; Zamani-Gharehchamani, E.; Nozari, S.; Nouri, M.; Jodati, A. Sulforaphane modulates CX3CL1/CX3CR1 axis and inflammation in palmitic acid-induced cell injury in C2C12 skeletal muscle cells. Mol. Biol. Rep. 2020, 47, 7971–7977. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.C.; Li, S.J.; Yang, C.L.; Xue, R.L.; Xi, Y.Y.; Wang, L.; Zhao, Q.L.; Li, D.J. Sulforaphane Attenuates Muscle Inflammation in Dystrophin-deficient mdx Mice via NF-E2-related Factor 2 (Nrf2)-mediated Inhibition of NF-kappaB Signaling Pathway. J. Biol. Chem. 2015, 290, 17784–17795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.; Komine, S.; Warabi, E.; Akiyama, K.; Ishii, A.; Ishige, K.; Mizokami, Y.; Kuga, K.; Horie, M.; Miwa, Y.; et al. Nuclear factor (erythroid derived 2)-like 2 activation increases exercise endurance capacity via redox modulation in skeletal muscles. Sci. Rep. 2017, 7, 12902. [Google Scholar] [CrossRef] [Green Version]
- Malaguti, M.; Angeloni, C.; Garatachea, N.; Baldini, M.; Leoncini, E.; Collado, P.S.; Teti, G.; Falconi, M.; Gonzalez-Gallego, J.; Hrelia, S. Sulforaphane treatment protects skeletal muscle against damage induced by exhaustive exercise in rats. J. Appl. Physiol. 2009, 107, 1028–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, C.; Alves, I.; Singh, P.; Palade, P.T.; Carvalho, E.; Borsheim, E.; Jun, S.R.; Cheema, A.; Boerma, M.; Awasthi, S.; et al. Sulforaphane prevents age-associated cardiac and muscular dysfunction through Nrf2 signaling. Aging Cell 2020, 19, e13261. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Pu, D.; Zhao, Y.; Chen, J.; Zhu, S.; Lu, A.; Liao, Z.; Sun, Y.; Xiao, Q. Sulforaphane protects against skeletal muscle dysfunction in spontaneous type 2 diabetic db/db mice. Life Sci. 2020, 255, 117823. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.C.; Ni, C.L.; Shen, Y.C.; Huang, Y.L.; Kuo, C.H.; Wu, T.S.; Chen, C.C. Phenolic constituents from the stem bark of Magnolia officinalis. J. Nat. Prod. 2009, 72, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Zhang, L.; Ye, P.P.; Tang, X.J.; Zhang, Y.D. Protective effect of magnolol against hydrogen peroxide-induced oxidative stress in human lens epithelial cells. Am. J. Chin. Med. 2009, 37, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, E.; Park, J.; Jung, K.; Lee, S.; Hong, S.; Park, J.; Park, E.; Kim, J.; Park, S.; et al. Anti-inflammatory effects of magnolol and honokiol are mediated through inhibition of the downstream pathway of MEKK-1 in NF-kappaB activation signaling. Planta Med. 2005, 71, 338–343. [Google Scholar] [CrossRef]
- Chao, L.K.; Liao, P.C.; Ho, C.L.; Wang, E.I.; Chuang, C.C.; Chiu, H.W.; Hung, L.B.; Hua, K.F. Anti-inflammatory bioactivities of honokiol through inhibition of protein kinase C, mitogen-activated protein kinase, and the NF-kappaB pathway to reduce LPS-induced TNFalpha and NO expression. J. Agric. Food Chem. 2010, 58, 3472–3478. [Google Scholar] [CrossRef]
- Saito, J.; Shibuya, K.; Nagase, H. Anti-clastogenic effect of magnolol on benzo(a)pyrene-induced clastogenicity in mice. Food Chem. Toxicol. 2008, 46, 694–700. [Google Scholar] [CrossRef]
- Lee, Y.J.; Lee, Y.M.; Lee, C.K.; Jung, J.K.; Han, S.B.; Hong, J.T. Therapeutic applications of compounds in the Magnolia family. Pharmacol. Ther. 2011, 130, 157–176. [Google Scholar] [CrossRef]
- Chiang, J.; Shen, Y.C.; Wang, Y.H.; Hou, Y.C.; Chen, C.C.; Liao, J.F.; Yu, M.C.; Juan, C.W.; Liou, K.T. Honokiol protects rats against eccentric exercise-induced skeletal muscle damage by inhibiting NF-kappaB induced oxidative stress and inflammation. Eur. J. Pharm. 2009, 610, 119–127. [Google Scholar] [CrossRef]
- Ge, Z.; Liu, D.; Shang, Y.; Li, Y.; Chen, S.Z. Magnolol inhibits myotube atrophy induced by cancer cachexia through myostatin signaling pathway in vitro. J. Nat. Med. 2020, 74, 741–749. [Google Scholar] [CrossRef]
- Chen, M.C.; Chen, Y.L.; Lee, C.F.; Hung, C.H.; Chou, T.C. Supplementation of Magnolol Attenuates Skeletal Muscle Atrophy in Bladder Cancer-Bearing Mice Undergoing Chemotherapy via Suppression of FoxO3 Activation and Induction of IGF-1. PLoS ONE 2015, 10, e0143594. [Google Scholar] [CrossRef] [PubMed]
- Rettig, M.B.; Heber, D.; An, J.; Seeram, N.P.; Rao, J.Y.; Liu, H.; Klatte, T.; Belldegrun, A.; Moro, A.; Henning, S.M.; et al. Pomegranate extract inhibits androgen-independent prostate cancer growth through a nuclear factor-kappaB-dependent mechanism. Mol. Cancer 2008, 7, 2662–2671. [Google Scholar] [CrossRef] [Green Version]
- Adhami, V.M.; Khan, N.; Mukhtar, H. Cancer Chemoprevention by Pomegranate: Laboratory and Clinical Evidence. Nutr. Cancer 2009, 61, 811–815. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.-Z.; Wang, C.-M.; Li, L.; Ding, N.; Gao, X.-L. Effect of pomegranate peel polyphenols on human prostate cancer PC-3 cells in vivo. Food Sci. Biotechnol. 2015, 24, 1887–1892. [Google Scholar] [CrossRef]
- Bell, C.; Hawthorne, S. Ellagic acid, pomegranate and prostate cancer—A mini review. J. Pharm. Pharmacol. 2008, 60, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirode, A.B.; Kovvuru, P.; Chittur, S.V.; Henning, S.M.; Heber, D.; Reliene, R. Antiproliferative effects of pomegranate extract in MCF-7 breast cancer cells are associated with reduced DNA repair gene expression and induction of double strand breaks. Mol. Carcinog. 2014, 53, 458–470. [Google Scholar] [CrossRef]
- Paller, C.J.; Ye, X.; Wozniak, P.J.; Gillespie, B.K.; Sieber, P.R.; Greengold, R.H.; Stockton, B.R.; Hertzman, B.L.; Efros, M.D.; Roper, R.P.; et al. A randomized phase II study of pomegranate extract for men with rising PSA following initial therapy for localized prostate cancer. Prostate Cancer Prostatic Dis. 2013, 16, 50–55. [Google Scholar] [CrossRef]
- Rodriguez, J.; Caille, O.; Ferreira, D.; Francaux, M. Pomegranate extract prevents skeletal muscle of mice against wasting induced by acute TNF-α injection. Mol. Nutr. Food Res. 2017, 61, 1600169. [Google Scholar] [CrossRef]
- Trombold, J.R.; Reinfeld, A.S.; Casler, J.R.; Coyle, E.F. The effect of pomegranate juice supplementation on strength and soreness after eccentric exercise. J. Strength Cond. Res. 2011, 25, 1782–1788. [Google Scholar] [CrossRef]
- D’Angelo, S.; Rosa, R. The impact of supplementation with Pomegranate fruit (Punica Granatum L.) on sport performance. Sport Sci. 2020, 13, 29–37. [Google Scholar]
- Rapa, S.F.; Magliocca, G.; Pepe, G.; Amodio, G.; Autore, G.; Campiglia, P.; Marzocco, S. Protective Effect of Pomegranate on Oxidative Stress and Inflammatory Response Induced by 5-Fluorouracil in Human Keratinocytes. Antioxidants 2021, 10, 203. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhao, L.; Li, H.; Xu, H.; Chen, W.W.; Tao, L. Research progress on the anticarcinogenic actions and mechanisms of ellagic acid. Cancer Biol. Med. 2014, 11, 92–100. [Google Scholar] [CrossRef]
- Ekinci Akdemir, F.N.; Gülçin, İ.; Karagöz, B.; Soslu, R.; Alwasel, S.H. A comparative study on the antioxidant effects of hesperidin and ellagic acid against skeletal muscle ischemia/reperfusion injury. J. Enzym. Inhib. Med. Chem. 2016, 31, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Aslan, A.; Beyaz, S.; Gok, O.; Erman, O. The effect of ellagic acid on caspase-3/bcl-2/Nrf-2/NF-kB/TNF-α /COX-2 gene expression product apoptosis pathway: A new approach for muscle damage therapy. Mol. Biol. Rep. 2020, 47, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Periago, P.; Espín, J.C.; Tomás-Barberán, F.A. Identification of urolithin a as a metabolite produced by human colon microflora from ellagic acid and related compounds. J. Agric. Food Chem. 2005, 53, 5571–5576. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Pierre, N.; Naslain, D.; Bontemps, F.; Ferreira, D.; Priem, F.; Deldicque, L.; Francaux, M. Urolithin B, a newly identified regulator of skeletal muscle mass. J. Cachexia Sarcopenia Muscle 2017, 8, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Ceci, C.; Graziani, G.; Faraoni, I.; Cacciotti, I. Strategies to improve ellagic acid bioavailability: From natural or semisynthetic derivatives to nanotechnological approaches based on innovative carriers. Nanotechnology 2020, 31, 382001. [Google Scholar] [CrossRef]
- Al-Harbi, S.A.; Abdulrahman, A.O.; Zamzami, M.A.; Khan, M.I. Urolithins: The Gut Based Polyphenol Metabolites of Ellagitannins in Cancer Prevention, a Review. Front. Nutr. 2021, 8, 647582. [Google Scholar] [CrossRef]
- Luan, P.; D’Amico, D.; Andreux, P.A.; Laurila, P.-P.; Wohlwend, M.; Li, H.; Lima, T.I.d.; Place, N.; Rinsch, C.; Zanou, N.; et al. Urolithin A improves muscle function by inducing mitophagy in muscular dystrophy. Sci. Transl. Med. 2021, 13, eabb0319. [Google Scholar] [CrossRef]
- Ferreira de Oliveira, J.M.P.; Santos, C.; Fernandes, E. Therapeutic potential of hesperidin and its aglycone hesperetin: Cell cycle regulation and apoptosis induction in cancer models. Phytomedicine 2020, 73, 152887. [Google Scholar] [CrossRef]
- Khedr, N.F.; Khalil, R.M. Effect of hesperidin on mice bearing Ehrlich solid carcinoma maintained on doxorubicin. Tumor Biol. 2015, 36, 9267–9275. [Google Scholar] [CrossRef]
- Cheng, Y.; Di, S.; Fan, C.; Cai, L.; Gao, C.; Jiang, P.; Hu, W.; Ma, Z.; Jiang, S.; Dong, Y.; et al. SIRT1 activation by pterostilbene attenuates the skeletal muscle oxidative stress injury and mitochondrial dysfunction induced by ischemia reperfusion injury. Apoptosis 2016, 21, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Obrador, E.; Salvador-Palmer, R.; Jihad-Jebbar, A.; López-Blanch, R.; Dellinger, T.H.; Dellinger, R.W.; Estrela, J.M. Pterostilbene in Cancer Therapy. Antioxidants 2021, 10, 492. [Google Scholar] [CrossRef]
- Chen, R.J.; Ho, C.T.; Wang, Y.J. Pterostilbene induces autophagy and apoptosis in sensitive and chemoresistant human bladder cancer cells. Mol. Nutr. Food Res. 2010, 54, 1819–1832. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Liu, W.; Zhu, X.; Ran, L.; Lang, H.; Yi, L.; Mi, M.; Zhu, J. Pterostilbene Enhances Endurance Capacity via Promoting Skeletal Muscle Adaptations to Exercise Training in Rats. Molecules 2020, 25, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metti, S.; Gambarotto, L.; Chrisam, M.; Spina, M.L.; Baraldo, M.; Braghetta, P.; Blaauw, B.; Bonaldo, P. The Polyphenol Pterostilbene Ameliorates the Myopathic Phenotype of Collagen VI Deficient Mice via Autophagy Induction. Front. Cell Dev. Biol. 2020, 8, 997. [Google Scholar] [CrossRef]
- Assi, M.; Derbre, F.; Lefeuvre-Orfila, L.; Rebillard, A. Antioxidant supplementation accelerates cachexia development by promoting tumor growth in C26 tumor-bearing mice. Free Radic. Biol. Med. 2016, 91, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, V.; Park, Y.; Chen, C.C.; Xu, P.Z.; Chen, M.L.; Tonic, I.; Unterman, T.; Hay, N. Akt determines replicative senescence and oxidative or oncogenic premature senescence and sensitizes cells to oxidative apoptosis. Cancer Cell 2008, 14, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, G.; Madeddu, C.; Maccio, A. Cachexia and oxidative stress in cancer: An innovative therapeutic management. Curr. Pharm. Des. 2012, 18, 4813–4818. [Google Scholar] [CrossRef] [PubMed]


| Target | Experimental Setting | UA treatments | Findings | References |
|---|---|---|---|---|
| Regulating muscle mass and overcoming different muscle wasting in various conditions | In vivo 6–8-week-old male C57BL/6 mice | 24 h fasting: i.p. injection of 200 mg/kg given at 0 and 12 h time points | Reduced fasting-induced muscle atrophy: | [97] |
| ↑ lower limb muscle mass by 7% ± 2% | ||||
| ↓ atrogin-1 and MuRF1 mRNA expression | ||||
| Denervation: i.p. injection of 200 mg/kg, twice daily for 7 days | Decreased denervation-induced muscle loss: | |||
| ↑ muscle fiber size (hindlimb muscles) | ||||
| Hypertrophy test: Chow containing 0.27% UA for 5 weeks | Induced muscle hypertrophy: | |||
| ↑ muscle mass, fiber size and grip strength | ||||
| ↑ IGF-1 mRNA expression | ||||
| Sustaining resistance exercise-induced mTORC1 activity | In vivo 10-week-old male Sprague–Dawley rats | 250 mg/kg, i.p. injection right after exercise | Sustained phosphorylation of p70S6K | [100] |
| ↑ Akt Thr308 phosphorylation | ||||
| Failed to increase muscle IGF-I concentrations with UA treatment alone | ||||
| Increasing muscle mass and function | In vivo 8-week-old male C57BL/6 mice | High fat diet supplemented with 0.14% UA for 6 weeks | ↑ Akt phosphorylation | [99] |
| ↑ muscle mass | ||||
| ↑ slow and fast muscle fiber size | ||||
| ↑ grip strength | ||||
| ↑ food intake, energy expenditure | ||||
| Promoting muscle regeneration | In vitro Skeletal muscle satellite cells isolated from 10-day-old C57BL/6 mice | In vitro 10 μM treatment for 10 days | In vitro | [101,102] |
| ↑ Pax7 expression | ||||
| ↑ sirtuin 1 (SIRT1), Peroxisome proliferator-activated receptor-gamma coactivator-1α (PGC-1α) expression | ||||
| In vivo 10-month-old male C57BL/6 mice | In vivo i.p. injection of 200 mg/kg, twice daily for 7 days | In vivo | ||
| ↓ body weight | ||||
| Change muscle composition to be more oxidative | ||||
| ↑ myoglobin expression | ||||
| ↓ cellular energy status (ATP, ADP) | ||||
| ↑ fiber generation | ||||
| Attenuating hypobaric hypoxia-induced skeletal muscle wasting via Akt signaling pathway | In vivo Male Sprague–Dawley rats | 20 mg/kg/day via oral gavage for 3 days | ↓ ROS level, protein oxidation | [98] |
| ↑ antioxidative enzymes: GPx, GR, SOD1, SOD2 and catalase | ||||
| ↑ glutathione (GSH) activation | ||||
| ↓ caspases 3 | ||||
| ↓ IL-1β, IL-10, IL-4, TNF-α | ||||
| ↓ weight loss | ||||
| ↑ muscle protein contents | ||||
| ↑ grip strength | ||||
| Enhanced Akt phosphorylation, IGF-1 protein expression, p70S6K |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Swiderski, K.; Murphy, K.T.; Lynch, G.S. Role for Plant-Derived Antioxidants in Attenuating Cancer Cachexia. Antioxidants 2022, 11, 183. https://doi.org/10.3390/antiox11020183
Li W, Swiderski K, Murphy KT, Lynch GS. Role for Plant-Derived Antioxidants in Attenuating Cancer Cachexia. Antioxidants. 2022; 11(2):183. https://doi.org/10.3390/antiox11020183
Chicago/Turabian StyleLi, Wenlan, Kristy Swiderski, Kate T. Murphy, and Gordon S. Lynch. 2022. "Role for Plant-Derived Antioxidants in Attenuating Cancer Cachexia" Antioxidants 11, no. 2: 183. https://doi.org/10.3390/antiox11020183