1. Introduction
The gut microbiota of ruminants is composed of a diverse microbial community of mostly bacteria, protozoa, fungi, and archaea, responsible for feeds digestion for supplying nutrients to the host animals [
1]. The symbiotic relationship of the rumen microbes helps the utilization of fibrous feeds and the production of microbial proteins. However, all the microbes present in the rumen are not always good for animals, as some of them might not have any beneficial effect on feed utilization and animal productivity [
2]. Protozoa do not have specific role to play in rumen fermentation of animals fed poor quality fibrous feeds [
3]. Methanogenic archaea present in the rumen produce methane from the carbon dioxide and hydrogen produced as byproduct of feed fermentation and emitted to the environment via eructation mainly [
4]. Livestock production contributes about 37–44% of global methane emissions and is a major source of methane production in the agriculture sector [
5]. The excess degradation of dietary proteins by rumen microbes beyond the capacity of microbial protein synthesis results in ammonia emissions, which are, further, converted into nitrous oxide, a potent greenhouse gas triggering environmental pollution [
6]. These inefficiencies in the rumen are responsible for a loss of feed energy (5–12%) and nitrogen (25–35%) [
7,
8], which also attract the attention of animal nutritionists for their minimization and diversion into animal production. Therefore, the modulation of rumen functions to improve animal production and reduce environmental pollution through phytogenic feed additives have gained focus to lessen the use of antibiotics in animal feeds [
9] owing to consumer demand for healthy animal products.
In monogastric animals, especially poultry and pigs, phytogenic feed additives or phytobiotics are used as an alternative to antibiotics to increase feed efficiency, nutrient bioavailability, and health status by modulating gut microbiome and host immunity [
10]. Many essential oil compounds (cinnamaldehyde, thymol, carvacrol, and eugenol), used either individually or in blends, have been reported to enhance innate immunity and resistance against enteric diseases in poultry, in addition to improving feed efficiency [
11]. The effectiveness of a phytochemical depends on its type, composition, active component, inclusion level, and environmental circumstances. The dietary inclusion of phytochemicals in pigs and poultry is reported to decrease intestinal challenges by stabilizing gut microbiota and reducing toxic metabolites produced by pathogenic microbes. A reduction in oxidative stress with increased antioxidant activities in tissues was established in phytochemical supplemented broiler birds. Modulation of the immune system by increased proliferation of immune cells, suppression of proinflammatory cytokines, and enhanced membrane integrity and antibody titer was described in monogastricts with dietary supplementation of phytogenic feed additives [
12,
13].
Phytochemicals (saponins, tannins, and essential oils) present in feed additives of natural sources are designated to modulate the functioning of the rumen microbiome for reducing methanogenesis and protein degradation with the goal to enhance the nutritional value of feeds and further the abatement of environmental pollution from livestock production [
6,
14]. The antimicrobial properties of plant bioactive compounds, especially tannins and essential oils, facilitate the modulation of the rumen microbial ecosystem; however, their effects on health performance, including antioxidant status and immune response, is very scanty. Limited investigation into ruminants suggests improvements in antioxidant enzymes and immunity indicators by dietary supplementation of tanniferous leaves or rumen protected capsicum [
15,
16,
17].
Buffaloes are an integral part of the livestock production system in South Asian countries, and 57% of global buffalo population is present in India [
18]. Contributing nearly 50% of total milk production, buffaloes play an important role in food and the nutritional security of the country [
19]. With the contribution of buffalo meat alone, India is one of the largest beef exporting countries of the world. Poplar (
Populus sp.) is a deciduous plant widespread in Europe and Asia, and leaves are rich in phytochemicals (phenolic compounds, terpenoids, flavones, flavanones, and many compounds in essential oils) [
20,
21]. Eucalyptus (
Eucalyptus sp.) leaves contain an array of volatile essential oil compounds, such as 1,8-cineole, limonene, and α-terpineol, and non-volatile phenolic compounds, such as quercetin, epicatechin, and catechin, which are associated with health benefits [
22]. The phytochemicals present in these tree leaves may modulate rumen fermentation towards an inhibition of methanogenesis, as well stimulate the immune system and antioxidant status of animals. However, there is no study investigating the effect of dietary poplar–eucalyptus leaf-meal blends on rumen fermentation, growth performance, and health status in ruminants. Therefore, the present investigation was designed to examine the impact of dietary phytogenic composite feed additives containing a mixture of dried leaves of
P. deltoides and
E. citriodora on immune response, antioxidant status, growth performance and nutrient utilization of buffalo (
Bubalus bubalis) calves. The outcome of the present experiment would explore the potential of these tree leaf blends as a natural feed additive for improving immunity in, and the health status of, buffaloes, in addition to enhancing growth performance and reducing environmental pollution through enteric methane mitigation.
3. Results
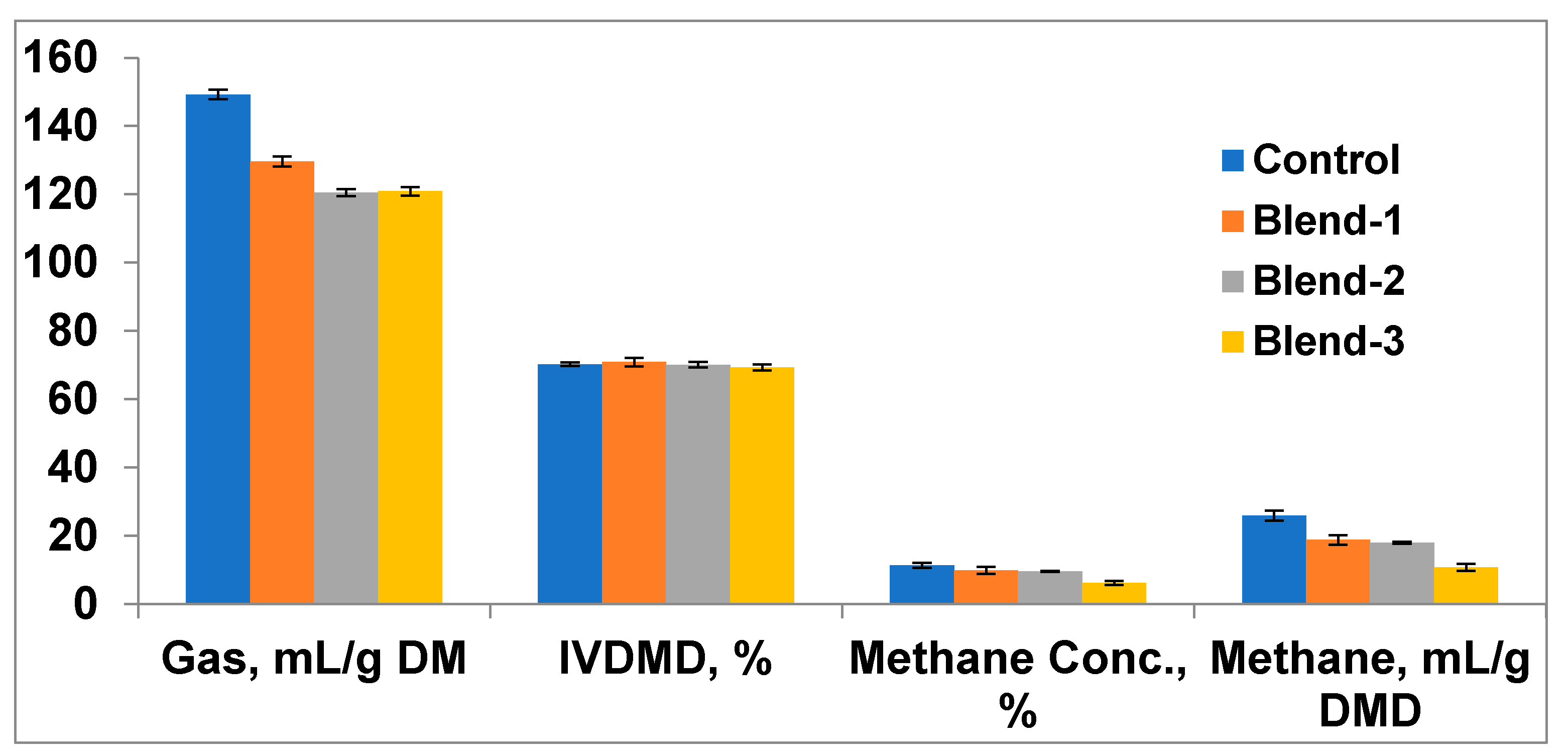
The total gas production after 24 h incubation of oat hay was reduced (
p < 0.01) in Blend-2 and Blend-3; however, it remained comparable (
p > 0.05) with the control for Blend-1 (
Table 1). The methane concentration (%) in the head space gas and total methane production were reduced (
p < 0.01) in all the treatments (Blend-1, Blend-2, and Blend-3) as compared to the control. Among the treatments, the methane reduction (
p < 0.01) was more pronounced in Blend-3 supplementation in comparison to Blend-1 and Blend-2. There was no significant (
p > 0.05) difference in gas production and methane production between Blend-1 and Blend-2. The concentration of ammonia nitrogen in the fermentation fluid was reduced (
p < 0.05) in all the treatment groups in comparison to the control. Although differences (
p < 0.05) were observed among the treatments, the highest reduction was recorded with Blend-3 supplementation, followed by Blend-2 and Blend-1. The inclusion of eucalyptus–poplar leaf-meal extract at any of the experimental dose levels showed comparable (
p > 0.05) feed degradability (IVDMD) with the non-supplemented control.
Among the volatile fatty acids, acetate production remained similar (
p > 0.05), irrespective of treatments; however, propionate concentration was increased (
p < 0.001) in Blend-2 and Blend-3 treatments as compared to the control, while Blend-1 remained comparable with both the control and Blend-2 (
Table 1). Butyrate concentration was not affected in either Blend-1 or Blend-2, as compared to the control; however, an increased (
p < 0.001) concentration was recorded with the supplementation of Blend-3. The increase in propionate production, owing to the supplementation of various blends of eucalyptus–poplar leaves extract, reduced (
p < 0.01) the ratio of acetate to propionate in all supplemented treatments as compared to the control; however, it was more (
p < 0.01) pronounced in Blend-3 supplementation. Among the treatments, Blend-1 and Blend-2 were comparable in all these parameters; however, methane production was reduced significantly (
p < 0.05) in Blend-3 as compared to Blend-1 and Blend-2. Therefore, using equivalent dose levels to treatment groups, Blend-1 and Blend-3 were used as additives for animal feeding in the in vivo experiment.
The basal feed consisted of the concentrate mixture, green oats fodder, and wheat straw (
Table 2), confirmed to have sufficient nutrient quality to satisfy the requirements of growing buffalo calves [
27]. The protein content of poplar leaves was more than the eucalyptus leaves; however, the crude extracted oil is higher in eucalyptus leaves than poplar, resulting in higher essential oils in the extract. The total phenolics and tannin fractions remained higher in eucalyptus leaves in comparison to poplar leaves, suggesting the presence of higher bioactives. The average initial as well as final body weight of all the experimental buffalo calves (CONT, EPLM-1, and EPLM-2) remained comparable (
p > 0.05), which ultimately resulted in a similar body weight gain for all the experimental groups. The quantities of concentrate mixture, green oats fodder, and supplements offered to individual buffalo calves during the entire experimental feeding period of 90 days were consumed in whole, resulting in a similar (
p > 0.05) intake of these feed ingredients. The phytogenic feed supplement provided (per head/day) total phenolics, tannin phenolics, and condensed tannins of 3.19 g, 2.30 g, and 0.71 g and 9.57 g, 6.90 g, and 2.14 g, respectively for EPLM-1 and EPLM-2 group calves. The comparable (
p > 0.05) total feed intake for all three experimental groups (
Table 3), although wheat straw was given ad libitum, culminated in a similar (
p > 0.05) feed conversion ratio (FCR) as well as feed efficiency (FE).
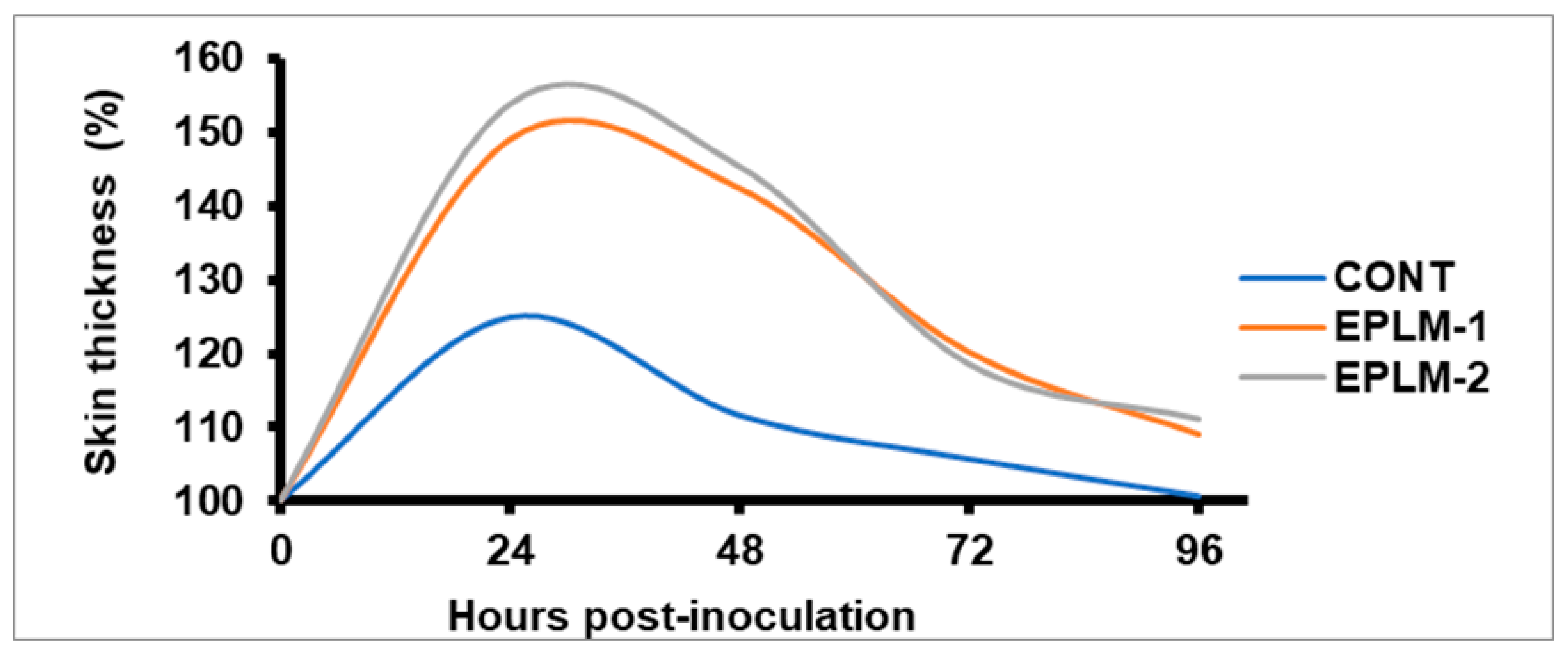
The skin thickness (%) was highest at 24 h after injection of PHA-P in all the experimental buffalo calves, before subsiding gradually, and coming near to normal after 96 h (
Table 4). At any hour post-injection (24, 48, 72, and 96 h), it remained higher (
p < 0.05) for both groups of EPLM supplemented calves than for the control. However, no difference (
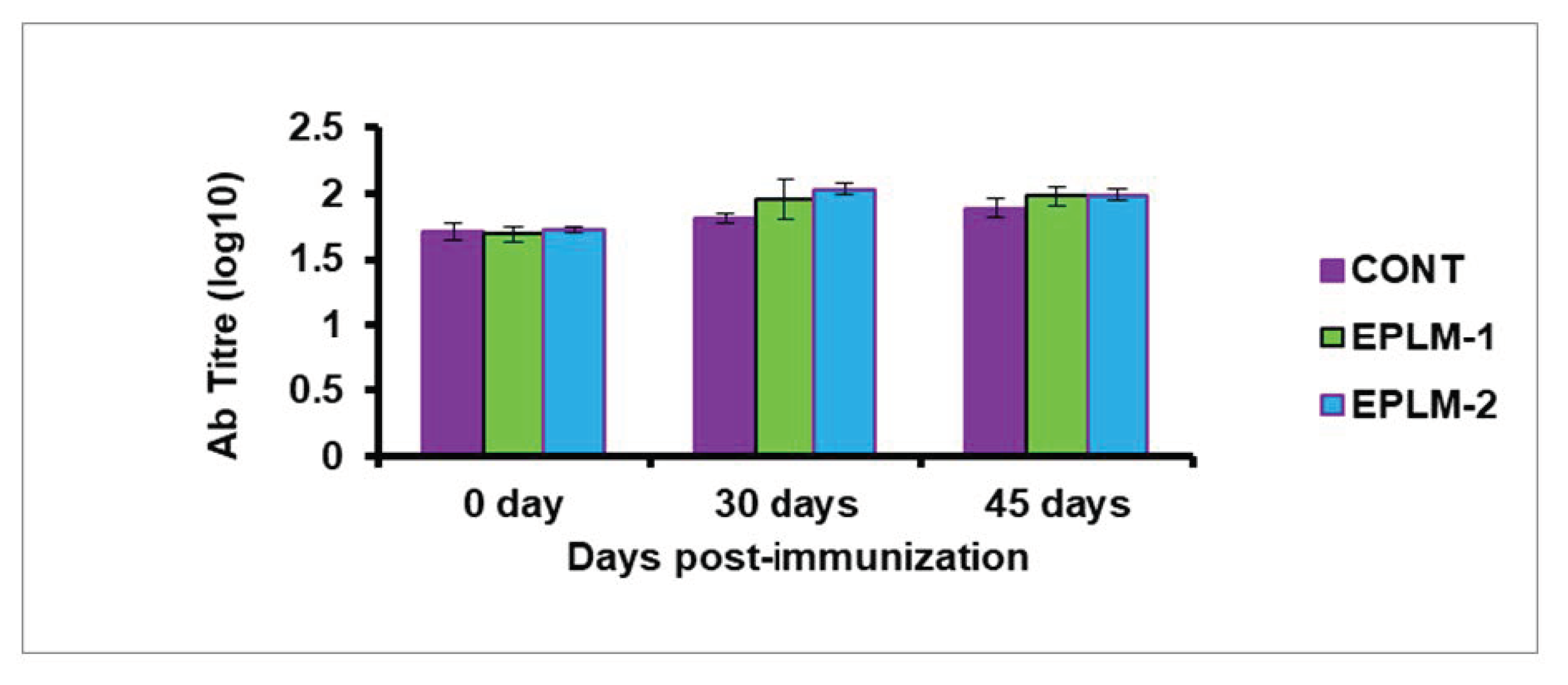
p > 0.05) was detected between the two supplemented groups (EPLM-1 and EPLM-2), suggesting the triviality of EPLM doses in stimulating the immune system. The antibody titer (log
10) of buffalo calves supplemented with EPLM was enhanced (
p < 0.05) at 30 days post-vaccination against
Pasteurella multoicida compared to the control; however, it remained comparable (
p > 0.05) between the supplemented calves (EPLM-1 and EPLM-2). A trend of increased (
p = 0.083) antibody tire was recorded at 45 days post-vaccination in both supplemented groups in comparison to the control (
Table 5).
The erythrocytic reduced glutathione (GSH), catalase (CAT), and superoxide dismutase (SOD) activities were enhanced (
p < 0.01) in both groups of EPLM supplemented buffalo calves in comparison to control. The reduced (
p < 0.01) lipid peroxidation (LPO) and increased (
p < 0.01) total thiol groups (T-SH) in both treatment groups (EPLM-1 and EPLM-2), as compared to control group, were evidenced (
Table 6). However, no difference (
p > 0.05) in any of the indicators of antioxidant status was found between the two treatment groups (EPLM-1 and EPLM-2). The hemoglobin (Hb), packed cell volume (PCV), serum protein (albumin, globulin, and A:G ratio), and serum enzyme (ALT and AST) levels did not differ (
p > 0.05) in experimental buffalo calves, irrespective of feeding regime. However, the serum urea concentration was reduced (
p < 0.05) in both EPLM supplemented groups in comparison to the control, although no difference (
p > 0.05) was recorded between the EPLM-1 and EPLM-2 groups (
Table 7).
The digestibility coefficients of various constituents (dry matter, organic matter, crude protein, and ether extract) were comparable (
p > 0.05) in all the experimental calves, irrespective of feeding regime (
Table 8). The fiber (neutral detergent fiber and acid detergent fiber) digestibility was not affected (
p > 0.05) by the dietary supplementation of phytogenic composite feed additives (EPLM-1 and EPLM-2). The daily intake (g/kg W
0.75) of nutrients, viz., digestible dry matter, digestible organic matter, digestible crude protein, and total digestible nutrients, remained comparable (
p > 0.05) among the treatments. The nutrient density of the total ration (% digestible crude protein and % total digestible nutrients) was also similar (
p > 0.05) in all the experimental buffalo calves. The methane concentration (ppm) in the exhaled air before the dosing of the experimental feed additive did not differ (
p > 0.05) among the calves; however, a trend in the reduction in methane concentration was evident in calves of EPLM-1 (
p = 0.089) and EPLM-2 (
p = 0.061) groups at 3 months post-dosing of phytogenic composite feed additive (
Table 9). However, the reduction was non-significant (
p > 0.05), revealing 55.76 and 61.15% decreases in exhaled air methane concentration in EPLM-1 and EPLM-2 groups, respectively.
4. Discussion
The dietary modulation of the immune system is described by providing substrates at suitable concentrations to the immune cells, the deprivation of specific nutrients to the invading pathogens, or the direct stimulation of immune cells through hormones and metabolites [
42,
43]. Phytonutrients can regulate rumen fermentation by altering protein degradation, ruminal ammonia production, nutrient digestion, and volatile fatty acid proportions by modifying the rumen microbiome [
6,
44]. The reduction in total gas and methane production with supplementation in various blends (
Figure 2) in the present study demonstrated specific antimicrobial and antimethanogenic effects of the blends of eucalyptus and poplar leaves extracts, which are rich in phenolics and essential oils [
21,
22]. The similar in vitro degradability of the substrate in the treatments supplemented with extracts of eucalyptus and poplar leaves with the non-supplemented control signifies that the major fiber degrading microbes were unaffected at the present dose levels. In a study with various phytogenic bioactive compounds on in vitro rumen fermentation, Singh, et al. [
45] reported associative effects of plant extracts in modulating rumen fermentation in a dose dependent manner. The reduction in ammonia nitrogen concentration with supplementation of all the blends are suggestive of reduced protein degradation, owing to the inhibition of hyper-ammonia producing bacteria and other microbes involved in amino acid deamination [
6,
46]. The presence of bioactive compounds in the extract could be associated with the reduced ammonia nitrogen production, as tannins [
47,
48] and essential oils [
49,
50] are reported to reduce protein degradation. The shift in volatile fatty acids production with increased concentration of propionate, and thereby the reduced ratio of acetate to propionate, established an alteration of rumen fermentation towards reduced methanogenesis [
51,
52].
The similar feed intake in all the experimental buffalo calves, irrespective of levels of phytogenic feed additive, is indicative of good palatability of the composite diet. Many researchers [
53,
54] have reported reduced feed intake owing to the addition of phytogenic compounds in the diet. Similar to the present study, no variation in feed intake was reported when eucalyptus leaf-meal powder was added, either in the diet of beef calves [
55] or swamp buffaloes [
56]. The reports on dietary supplementation of poplar leaves or composite with eucalyptus leaf-meal on intake and performance of animals are scanty. The feed intake by animals due to inclusion of plant secondary compounds in the diet depends on the type and concentration of bioactive components [
49]. Although poplar and eucalyptus leaves are rich in essential oils and phenolics [
21,
22], a similar feed intake demonstrated no effects on the gut microbiome or digestive enzymes. The comparable nutrient intake and digestibility (
Table 8) in all the experimental buffalo calves was advocated by other studies [
57,
58] with plant bioactive compounds. However, decreased feed digestibility was reported [
2] due to a reduction in fiber digestion triggered by the inhibition of ruminal cellulolytic bacteria and anaerobic fungi. The supplementation of eucalyptus leaves demonstrated an increased ADG owing to enhanced nutrient digestibility in buffalo calves [
59]; however, reduced feed efficiency and nutrient utilization was reported in other studies with plant compounds [
60]. A similar ADG and feed efficiency of buffalo calves to the present study (
Table 3) is described by comparable feed intake and digestibility in all the experimental animals.
Although phytogenic bioactive compounds modulate structural and functional properties of gut microflora, the molecular events associated with the mucosa cells and habitat microbes that lead to a modulation of local or systemic immune functions is still unclear [
61]. The bioactive plant compounds, viz., tannins, flavonoids, and essential oils, influence the immune systems of animals through different modes of action, such as the binding of proteins to render them unavailable for utilization in rumen and, instead, use quality protein available at lower gut, the interference in active sites of pathogens, or enhancing antioxidant status [
62,
63]. The increased cell-mediated immune response (
Figure 3) and humoral immune response (
Figure 4) in all the supplemented calves could be associated with the selective effects of tannins and terpenoids present in the feed additive on gastrointestinal microbes [
64]. The probiotic effects of tannins and essential oils through the inhibition of proteolytic and hyper-ammonia producing bacteria [
6] with the stimulation of
Bifidobacterium and
Lactobacillus [
65] could indirectly stimulate the immune system.
The increased levels of indicators (
Table 6) of the antioxidant defense system in the supplemented calves are suggestive of a better health status, protecting cells from free radicals through enhanced endogenous antioxidant enzymes [
66]. The bioactive compounds present in the EPLM demonstrated enhanced antioxidant indices, as polyphenolic compounds and essential oils [
63,
67] are reported to enhance these enzymes. The improved antioxidant status also enhanced the immune response of the supplemented buffalo calves. In a study with phytogenic feed additive containing tannins and saponins, Choubey, et al. [
68] also reported enhanced antioxidant status and immune response in goats. The absence of any deviation in blood biochemical parameters described the general wellbeing and health of all experimental animals, describing no deleterious effect on liver or other organs owing to phytogenic feed additive supplementation [
63,
68]; however, a reduced serum urea concentration is indicative of better utilization of dietary protein [
69].
The reduction in methane concentration in the exhaled air of calves in supplemented groups advocated an improvement in rumen functions, owing to bioactive compounds (tannins and essential oils) present in the EPLM. The in vitro study with these plant extract blends also confirmed a reduced methane production (
Table 1). The inhibition of methane was attributed to a modulation of the rumen microbiome, either through direct inhabitation of methanogens in general or methanogenic archaeal diversity by phenolic compounds and terpenoids present in the supplemented feed additive [
6,
70]. The present study corroborated the report of Thao, Wanapat, Kang, and Cherdthong [
56], in which a reduction in methane production was also demonstrated in swamp buffalo supplemented with eucalyptus leaf-meal.








{kind=link}
{kind=link}
{kind=link}
{kind=link}