
Dietary Selenium Alleviated Mouse Liver Oxidative Stress and NAFLD Induced by Obesity by Regulating the KEAP1/NRF2 Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Studies
2.3. Serum Analysis
2.4. Glucose Tolerance Test
2.5. H&E Staining
2.6. Masson Staining
2.7. Cell Culture
2.8. Quantitative Real Time PCR (qPCR)
2.9. Western Blot
2.10. Annexin V/PI Apoptosis Staining
2.11. Mito Tracker
2.12. JC-1 Staining
2.13. Cell Viability Assay
2.14. Oil Red O Staining
2.15. Immunofluorescence
2.16. Statistical Analysis
3. Results
3.1. Effect of Dietary Selenium on Hepatic Injury and Insulin Resistance In Vivo Models of NAFLD
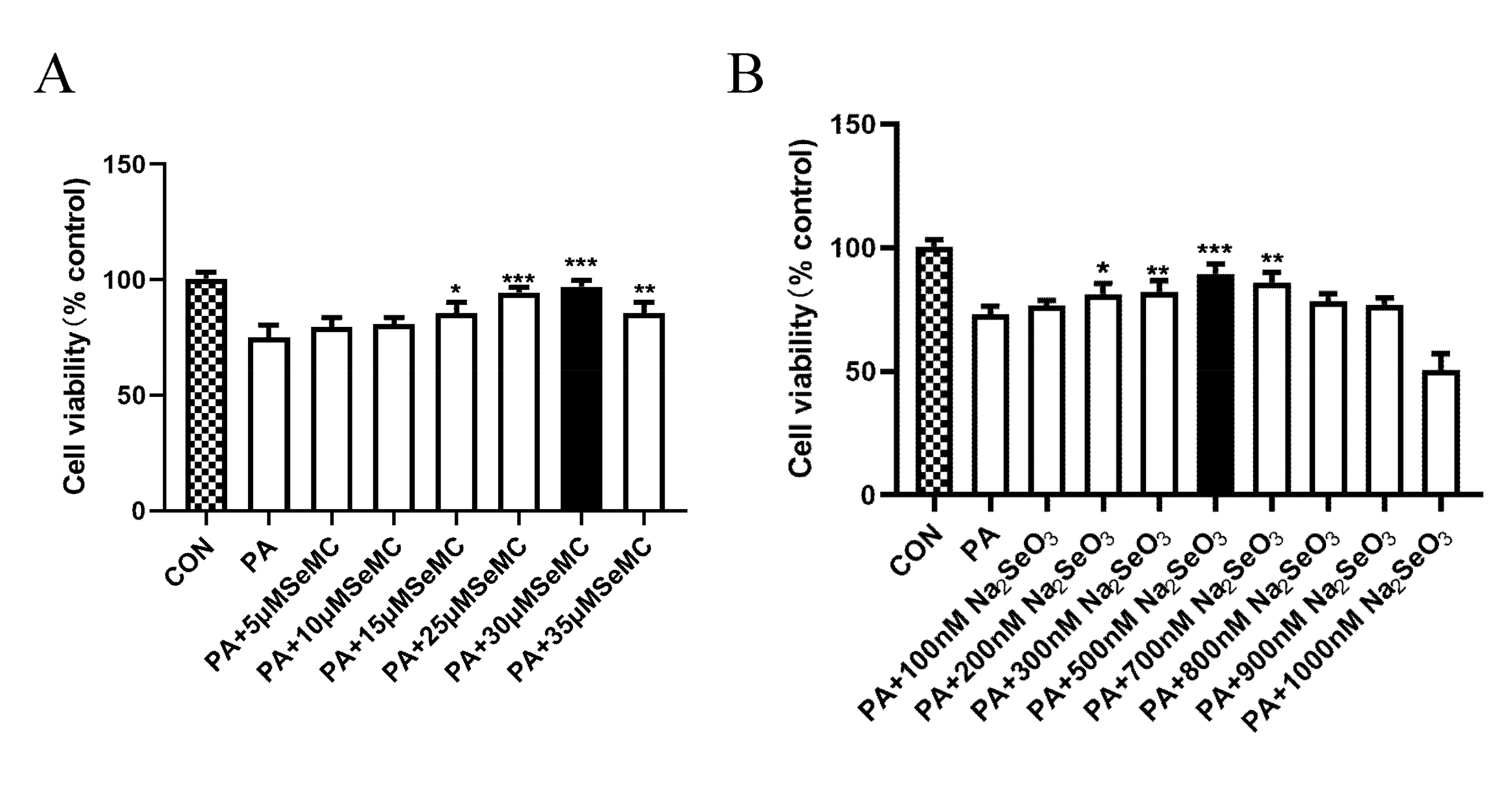
3.2. Selection of the Best Therapeutic Concentration of Na2SeO3 and L-SeMC in Palmitic Acid (PA)-Induced AML-12 Hepatocytes
3.3. Effects of Selenium on Lipid Accumulation In Vitro Models of NAFLD
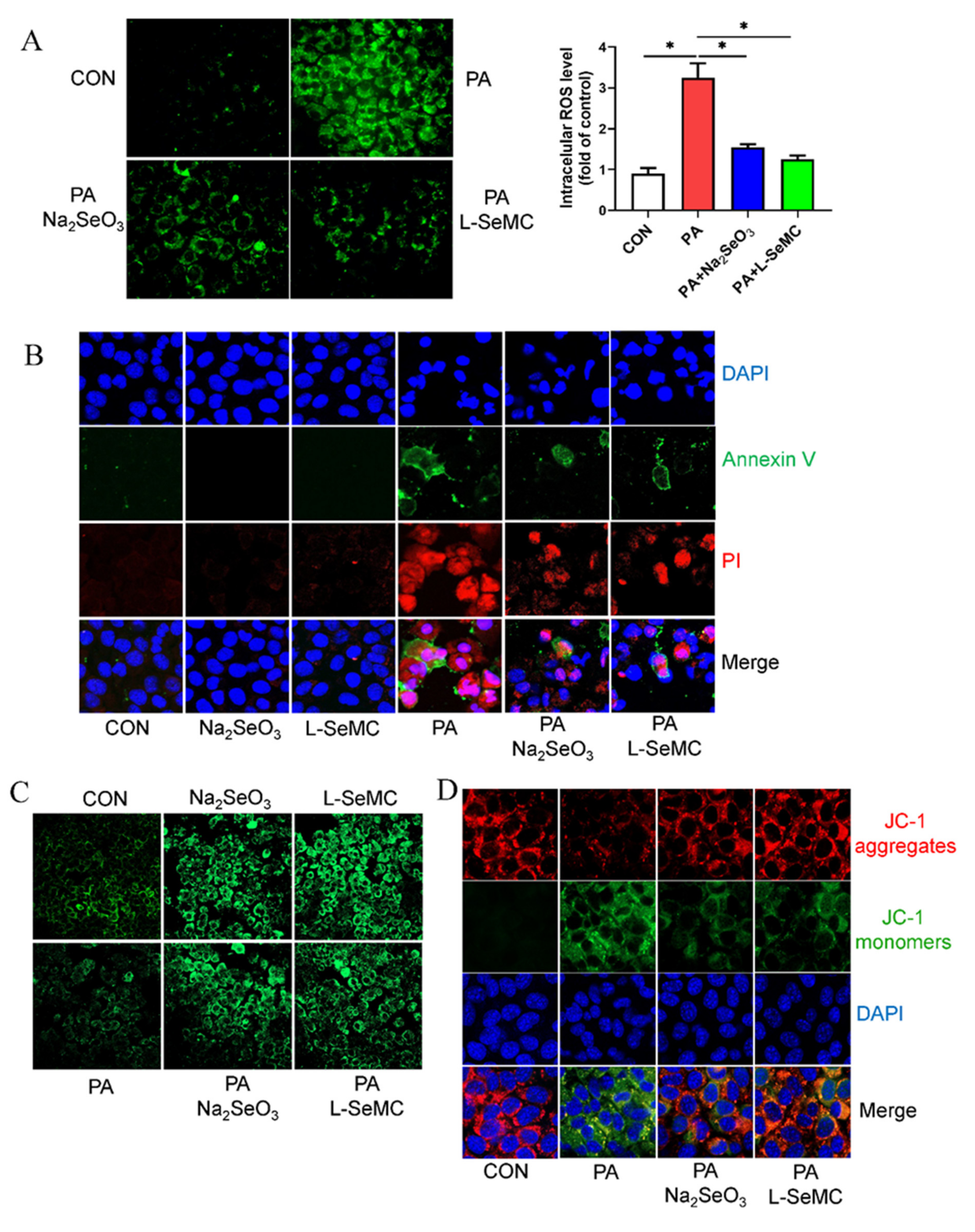
3.4. Effects of Selenium on Oxidative Stress in In Vitro Models of NAFLD
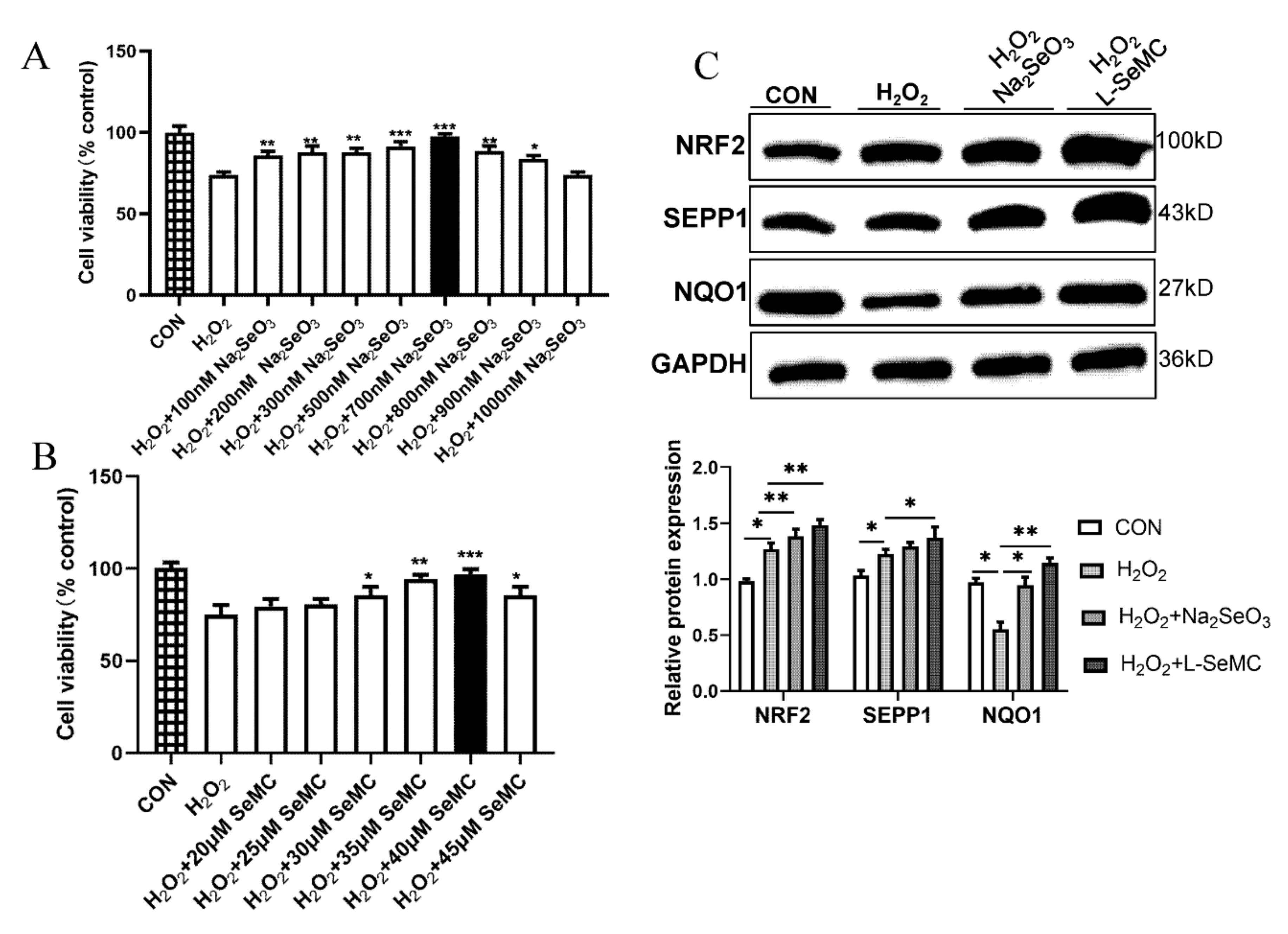
3.5. Effects of Selenium on the KEAP1/NRF2 Pathway in the In Vitro Models of NAFLD
3.6. Effects of Selenium on Oxidative Stress in H2O2-Induced AML-12 Hepatocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Danford, C.J.; Lai, M. NAFLD: A multisystem disease that requires a multidisciplinary approach. Frontline Gastroenterol. 2019, 10, 328–329. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.D.; Behary, J.; Zekry, A. Non-alcoholic fatty liver disease: A review of epidemiology, risk factors, diagnosis and management. Intern Med. J. 2020, 50, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Oates, J.R.; McKell, M.C.; Moreno-Fernandez, M.E.; Damen, M.; Deepe, G.S., Jr.; Qualls, J.E.; Divanovic, S. Macrophage Function in the Pathogenesis of Non-alcoholic Fatty Liver Disease: The Mac Attack. Front. Immunol. 2019, 10, 2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peverill, W.; Powell, L.W.; Skoien, R. Evolving concepts in the pathogenesis of NASH: Beyond steatosis and inflammation. Int. J. Mol. Sci. 2014, 15, 8591–8638. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Farzanegi, P.; Dana, A.; Ebrahimpoor, Z.; Asadi, M.; Azarbayjani, M.A. Mechanisms of beneficial effects of exercise training on non-alcoholic fatty liver disease (NAFLD): Roles of oxidative stress and inflammation. Eur. J. Sport Sci. 2019, 19, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef]
- Colagar, A.H.; Marzony, E.T. Ascorbic Acid in human seminal plasma: Determination and its relationship to sperm quality. J. Clin. Biochem. Nutr. 2009, 45, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.O.; Song, K.H.; Lee, I.S.; Lee, S.J.; Kim, W.I.; Pak, S.W.; Shin, I.S.; Kim, T. Cimicifugae Rhizoma Extract Attenuates Oxidative Stress and Airway Inflammation via the Upregulation of Nrf2/HO-1/NQO1 and Downregulation of NF-kappaB Phosphorylation in Ovalbumin-Induced Asthma. Antioxidants 2021, 10, 1626. [Google Scholar] [CrossRef]
- Wang, C.; Guo, J.; Wu, Z. Combinative treatment of Curdione and docetaxel triggers reactive oxygen species (ROS)-mediated intrinsic apoptosis of triple-negative breast cancer cells. Bioengineered 2021, 12, 10037–10048. [Google Scholar] [CrossRef]
- Li, W.Y.; Guo, M.Y.; Liu, Y.Z.; Mu, W.W.; Deng, G.Z.; Li, C.Y.; Qiu, C.W. Selenium Induces an Anti-tumor Effect Via Inhibiting Intratumoral Angiogenesis in a Mouse Model of Transplanted Canine Mammary Tumor Cells. Biol. Trace Elem. Res. 2016, 171, 371–379. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Gao, X.J.; Cao, Y.G.; Jiang, H.C.; Wang, T.C.; Song, X.J.; Guo, M.Y.; Zhang, N.S. Selenium Deficiency Facilitates Inflammation Through the Regulation of TLR4 and TLR4-Related Signaling Pathways in the Mice Uterus. Inflammation 2015, 38, 1347–1356. [Google Scholar] [CrossRef]
- Leyva-Lopez, N.; Peraza-Arias, M.; Avalos-Soriano, A.; Hernandez, C.; Lizarraga-Velazquez, C.E.; Heredia, J.B. Polyphenolic Extracts from Spent Coffee Grounds Prevent H2O2-Induced Oxidative Stress in Centropomus viridis Brain Cells. Molecules 2021, 26, 6195. [Google Scholar] [CrossRef] [PubMed]
- Strauss, E.; Oszkinis, G.; Staniszewski, R. SEPP1 gene variants and abdominal aortic aneurysm: Gene association in relation to metabolic risk factors and peripheral arterial disease coexistence. Sci. Rep. 2014, 4, 7061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valea, A.; Georgescu, C.E. Selenoproteins in human body: Focus on thyroid pathophysiology. Hormones 2018, 17, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; He, X.J.; Chen, X.Y.; Li, Y.M.; Ke, Y.N. SeP is elevated in NAFLD and participates in NAFLD pathogenesis through AMPK/ACC pathway. J. Cell. Physiol. 2021, 236, 3800–3807. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, P.S.; Raja, S.; Udayakumar, S.H.; Chandrashekar, S.; Nataraj, S.M.; Devegowda, D. Role of NRF2 cascade in determining the differential response of cervical cancer cells to anticancer drugs: An in vitro study. Mol. Biol. Rep. 2021, 49, 109–119. [Google Scholar] [CrossRef]
- Morris, G.; Walker, A.J.; Walder, K.; Berk, M.; Marx, W.; Carvalho, A.F.; Maes, M.; Puri, B.K. Increasing Nrf2 Activity as a Treatment Approach in Neuropsychiatry. Mol. Neurobiol. 2021, 58, 2158–2182. [Google Scholar] [CrossRef]
- Cai, J.; Yang, J.; Chen, X.; Zhang, H.; Zhu, Y.; Liu, Q.; Zhang, Z. Melatonin ameliorates trimethyltin chloride-induced cardiotoxicity: The role of nuclear xenobiotic metabolism and Keap1-Nrf2/ARE axis-mediated pyroptosis. BioFactors 2021. [Google Scholar] [CrossRef]
- Spandidos, A.; Wang, X.; Wang, H.; Seed, B. PrimerBank: A resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2010, 38, D792–D799. [Google Scholar] [CrossRef] [Green Version]
- Yu, A.; Zhou, R.; Xia, B.; Dang, W.; Yang, Z.; Chen, X. NAMPT maintains mitochondria content via NRF2-PPARalpha/AMPKalpha pathway to promote cell survival under oxidative stress. Cell Signal. 2020, 66, 109496. [Google Scholar] [CrossRef]
- Chen, A.; Chen, X.; Cheng, S.; Shu, L.; Yan, M.; Yao, L.; Wang, B.; Huang, S.; Zhou, L.; Yang, Z.; et al. FTO promotes SREBP1c maturation and enhances CIDEC transcription during lipid accumulation in HepG2 cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Huang, J.; Wang, Y.; Yang, Z.; Chen, X. Chemerin causes lipid metabolic imbalance and induces passive lipid accumulation in human hepatoma cell line via the receptor GPR1. Life Sci. 2021, 278, 119530. [Google Scholar] [CrossRef]
- Paglialunga, S.; Dehn, C.A. Clinical assessment of hepatic de novo lipogenesis in non-alcoholic fatty liver disease. Lipids Health Dis. 2016, 15, 159. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Hu, Y.; Luo, Y.; Yang, X. Selenium-containing polysaccharides from Ziyang green tea ameliorate high-fructose diet induced insulin resistance and hepatic oxidative stress in mice. Food Funct. 2015, 6, 3342–3350. [Google Scholar] [CrossRef]
- Zhang, Q.; Qian, Z.Y.; Zhou, P.H.; Zhou, X.L.; Zhang, D.L.; He, N.; Zhang, J.; Liu, Y.H.; Gu, Q. Effects of oral selenium and magnesium co-supplementation on lipid metabolism, antioxidative status, histopathological lesions, and related gene expression in rats fed a high-fat diet. Lipids Health Dis. 2018, 17, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nido, S.A.; Shituleni, S.A.; Mengistu, B.M.; Liu, Y.; Khan, A.Z.; Gan, F.; Kumbhar, S.; Huang, K. Effects of Selenium-Enriched Probiotics on Lipid Metabolism, Antioxidative Status, Histopathological Lesions, and Related Gene Expression in Mice Fed a High-Fat Diet. Biol. Trace Elem. Res. 2016, 171, 399–409. [Google Scholar] [CrossRef]
- Luo, M.; Huang, S.; Zhang, J.; Zhang, L.; Mehmood, K.; Jiang, J.; Zhang, N.; Zhou, D. Effect of selenium nanoparticles against abnormal fatty acid metabolism induced by hexavalent chromium in chicken’s liver. Environ. Sci. Pollut. Res. Int. 2019, 26, 21828–21834. [Google Scholar] [CrossRef]
- Stapleton, S.R.; Garlock, G.L.; Foellmi-Adams, L.; Kletzien, R.F. Selenium: Potent stimulator of tyrosyl phosphorylation and activator of MAP kinase. Biochim. Biophys. Acta 1997, 1355, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Polyzos, S.A.; Kountouras, J.; Zavos, C.; Deretzi, G. Nonalcoholic fatty liver disease: Multimodal treatment options for a pathogenetically multiple-hit disease. J. Clin. Gastroenterol. 2012, 46, 272–284. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Adipokines in nonalcoholic fatty liver disease. Metabolism 2016, 65, 1062–1079. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.; Potter, J.J.; Liu, X.; Torbenson, M.S.; Mezey, E. Selenium supplementation decreases hepatic fibrosis in mice after chronic carbon tetrachloride administration. Biol. Trace Elem. Res. 2010, 133, 83–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Song, G.; Minuk, G.Y. Effects of hepatic stimulator substance, herbal medicine, selenium/vitamin E, and ciprofloxacin on cirrhosis in the rat. Gastroenterology 1996, 110, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.H.; Cheng, W.F.; Li, X.H.; Sun, J.Q.; Li, F.; Ma, L.; Xie, L.M. Effects of dietary supplementation with vitamin E and selenium on rat hepatic stellate cell apoptosis. World J. Gastroenterol. 2005, 11, 4957–4961. [Google Scholar] [CrossRef] [PubMed]
- Reja, M.; Makar, M.; Visaria, A.; Marino, D.; Rustgi, V. Increased serum selenium levels are associated with reduced risk of advanced liver fibrosis and all-cause mortality in NAFLD patients: National Health and Nutrition Examination Survey (NHANES) III. Ann. Hepatol. 2020, 19, 635–640. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, S.; Jiang, H.; Liu, B.; Lv, Z.; Guo, C.; Zhang, H. Effects of selenium on apoptosis and abnormal amino acid metabolism induced by excess fatty acid in isolated rat hepatocytes. Mol. Nutr Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Pittelli, M.; Formentini, L.; Faraco, G.; Lapucci, A.; Rapizzi, E.; Cialdai, F.; Romano, G.; Moneti, G.; Moroni, F.; Chiarugi, A. Inhibition of nicotinamide phosphoribosyltransferase: Cellular bioenergetics reveals a mitochondrial insensitive NAD pool. J. Biol. Chem. 2010, 285, 34106–34114. [Google Scholar] [CrossRef] [Green Version]
- Schafer, K.; Kyriakopoulos, A.; Gessner, H.; Grune, T.; Behne, D. Effects of selenium deficiency on fatty acid metabolism in rats fed fish oil-enriched diets. J. Trace Elem. Med. Biol. 2004, 18, 89–97. [Google Scholar] [CrossRef]
- Renko, K.; Hofmann, P.J.; Stoedter, M.; Hollenbach, B.; Behrends, T.; Kohrle, J.; Schweizer, U.; Schomburg, L. Down-regulation of the hepatic selenoprotein biosynthesis machinery impairs selenium metabolism during the acute phase response in mice. FASEB J. 2009, 23, 1758–1765. [Google Scholar] [CrossRef]
- Murano, K.; Ogino, H.; Okuno, T.; Arakawa, T.; Ueno, H. Role of Supplementary Selenium on the Induction of Insulin Resistance and Oxidative Stress in NSY Mice Fed a High Fat Diet. Biol. Pharm. Bull. 2018, 41, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, X. Reducing selenoprotein P expression suppresses adipocyte differentiation as a result of increased preadipocyte inflammation. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E77–E85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira Ulbrecht, M.O.; Goncalves, D.A.; Zanoni, L.Z.G.; do Nascimento, V.A. Association Between Selenium and Malondialdehyde as an Efficient Biomarker of Oxidative Stress in Infantile Cardiac Surgery. Biol. Trace Elem. Res. 2019, 187, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.N.; Faghihi, A.; Motaghinejad, M.; Shiasi, M.; Imanparast, F.; Amiri, H.L.; Shidfar, F. Zinc and Selenium Co-supplementation Reduces Some Lipid Peroxidation and Angiogenesis Markers in a Rat Model of NAFLD-Fed High Fat Diet. Biol. Trace Elem. Res. 2018, 181, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Hishida, A.; Okada, R.; Naito, M.; Morita, E.; Wakai, K.; Hamajima, N.; Hosono, S.; Nanri, H.; Turin, T.C.; Suzuki, S.; et al. Polymorphisms in genes encoding antioxidant enzymes (SOD2, CAT, GPx, TXNRD, SEPP1, SEP15 and SELS) and risk of chronic kidney disease in Japanese—Cross-sectional data from the J-MICC study. J. Clin. Biochem. Nutr. 2013, 53, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, H.; Nakagawa, Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic. Biol. Med. 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Abramov, A.Y. The emerging role of Nrf2 in mitochondrial function. Free Radic. Biol. Med. 2015, 88, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Farooqui, Z.; Mohammad, R.S.; Lokhandwala, M.F.; Banday, A.A. Nrf2 inhibition induces oxidative stress, renal inflammation and hypertension in mice. Clin. Exp. Hypertens. 2021, 43, 175–180. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene NCBI ID | Forward Primer | Reverse Primer |
|---|---|---|---|
| Mus Acc | 1299249 | CCGAGAAGCAGAAACACGACG | CTACCACATCAAGGCTCCGAAT |
| Mus Fasn | 7988 | ATGGATGAGACCTCCCCAAG | AGAGCTTCTTAAGTAGAGAC |
| Mus β-actin | 10455 | ACAGAGCCTCGCCTTTGCCGA | CATGCCCACCATCACGCCCTGG |
| Mus Srebp1 | 1313979 | ACCCTGGAGGACACGCTGCTAG | GCTTCTGCAAACCTGCGGGAAAC |
| Mus Sepp1 | 10016 | CTGACATTAAGGTGGTTGAC | CACAGCAATCTTCGGTTATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, B.; Wu, P.; Chu, Y.; Gui, S.; Zheng, Y.; Chen, X. Dietary Selenium Alleviated Mouse Liver Oxidative Stress and NAFLD Induced by Obesity by Regulating the KEAP1/NRF2 Pathway. Antioxidants 2022, 11, 349. https://doi.org/10.3390/antiox11020349
Wang Y, Liu B, Wu P, Chu Y, Gui S, Zheng Y, Chen X. Dietary Selenium Alleviated Mouse Liver Oxidative Stress and NAFLD Induced by Obesity by Regulating the KEAP1/NRF2 Pathway. Antioxidants. 2022; 11(2):349. https://doi.org/10.3390/antiox11020349
Chicago/Turabian StyleWang, Yi, Bingbing Liu, Peixuan Wu, Yi Chu, Sisi Gui, Yazhen Zheng, and Xiaodong Chen. 2022. "Dietary Selenium Alleviated Mouse Liver Oxidative Stress and NAFLD Induced by Obesity by Regulating the KEAP1/NRF2 Pathway" Antioxidants 11, no. 2: 349. https://doi.org/10.3390/antiox11020349