
Ageing Significantly Alters the Physicochemical Properties and Associated Cytotoxicity Profiles of Ultrafine Particulate Matters towards Macrophages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ageing Process of CB Particles
2.2. Characterization of CB Particles
2.3. Cell Culture
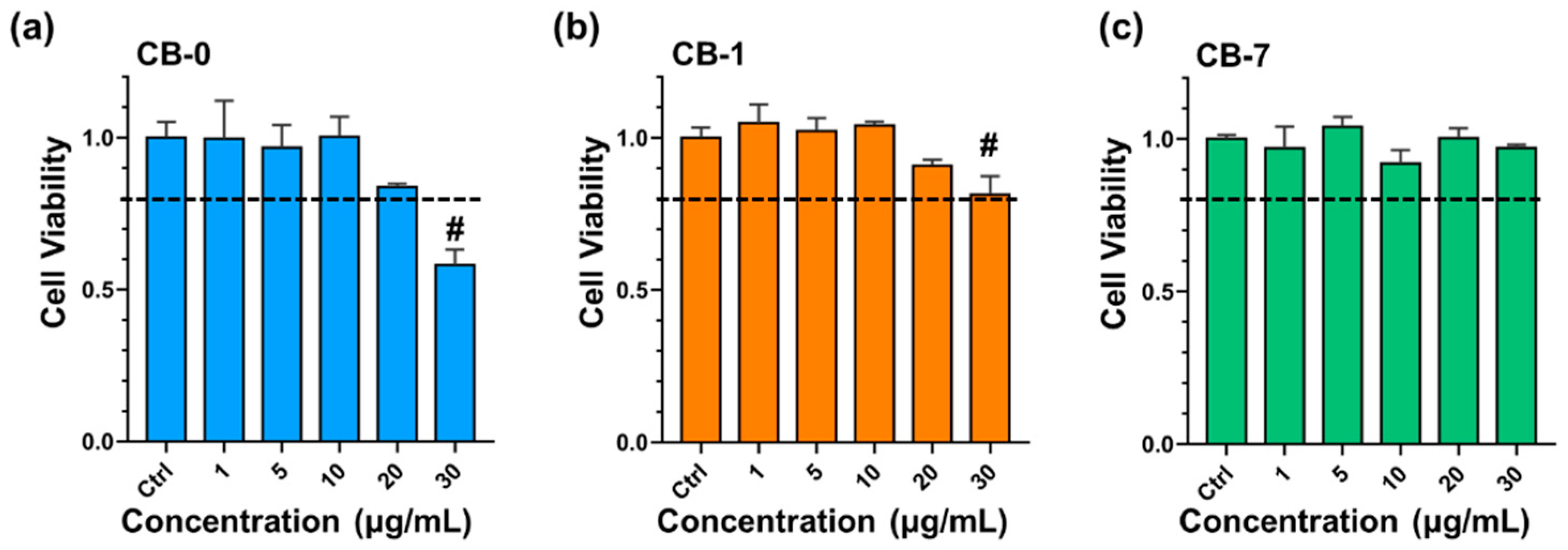
2.4. Cytotoxicity Assessment
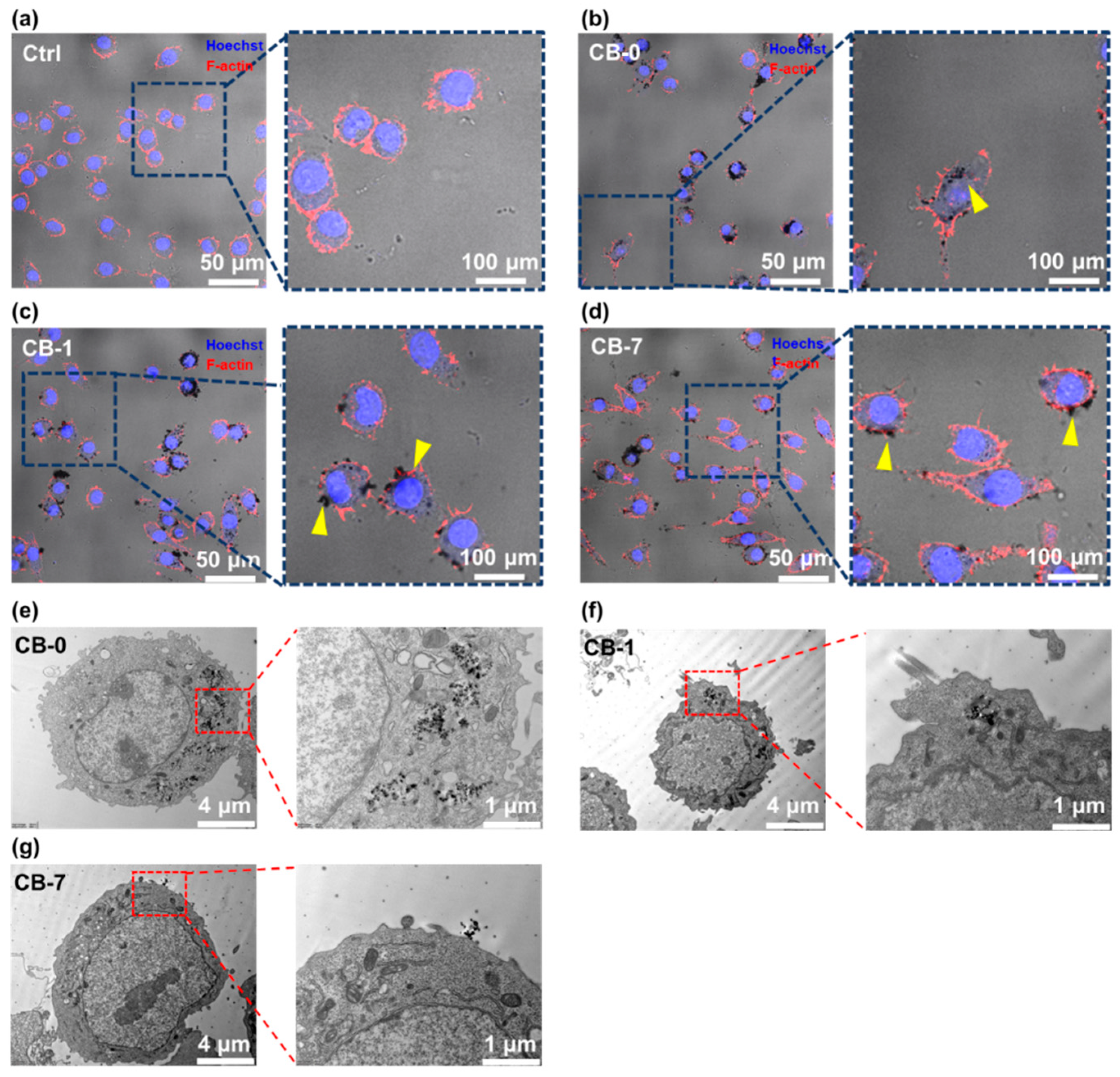
2.5. Characterization of the Cells by Confocal Microscopy
2.6. Characterization of Cells by Transmission Electron Microscope (TEM) for Cellular Structure
2.7. Intracellular ROS Production Assay of CB Particles
2.8. Coomassie Brilliant Blue Staining for Endocytosis Evaluation of CB Particles
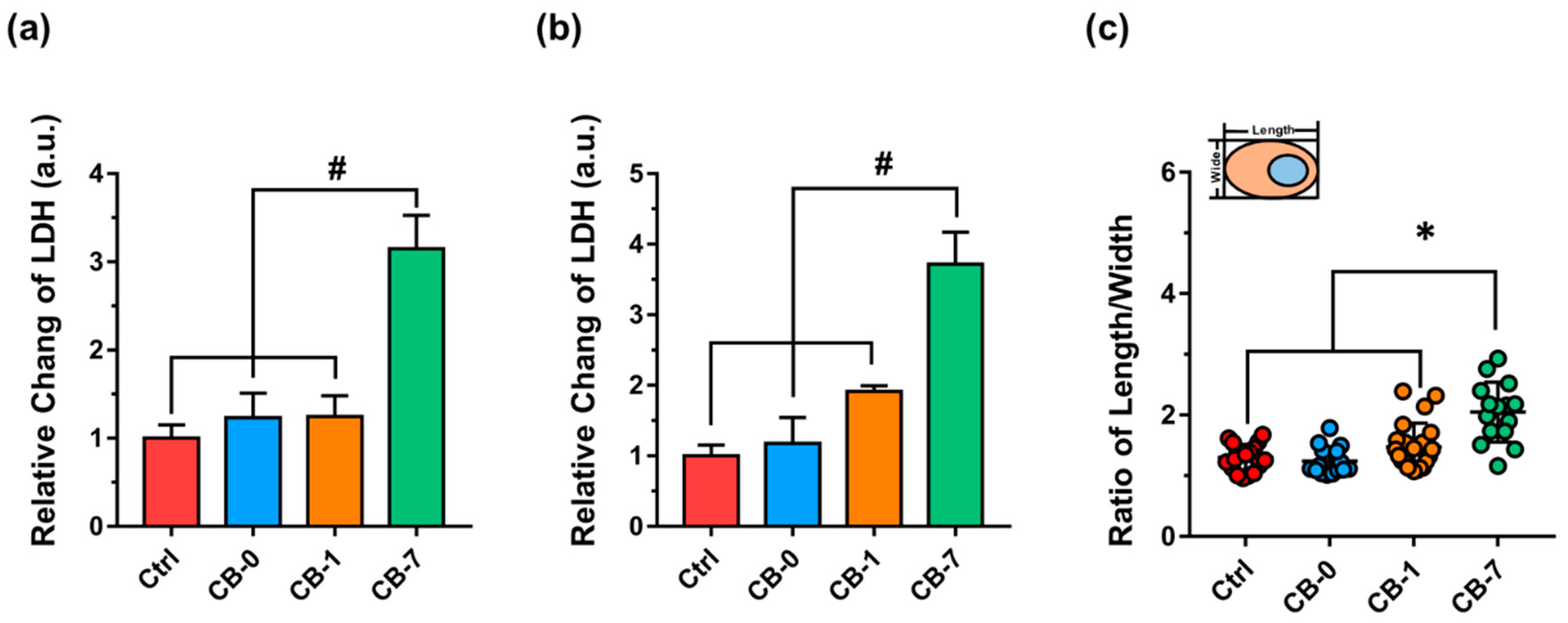
2.9. Determinations of Lactate Dehydrogenase (LDH) Release for Cell Membrane Integrity
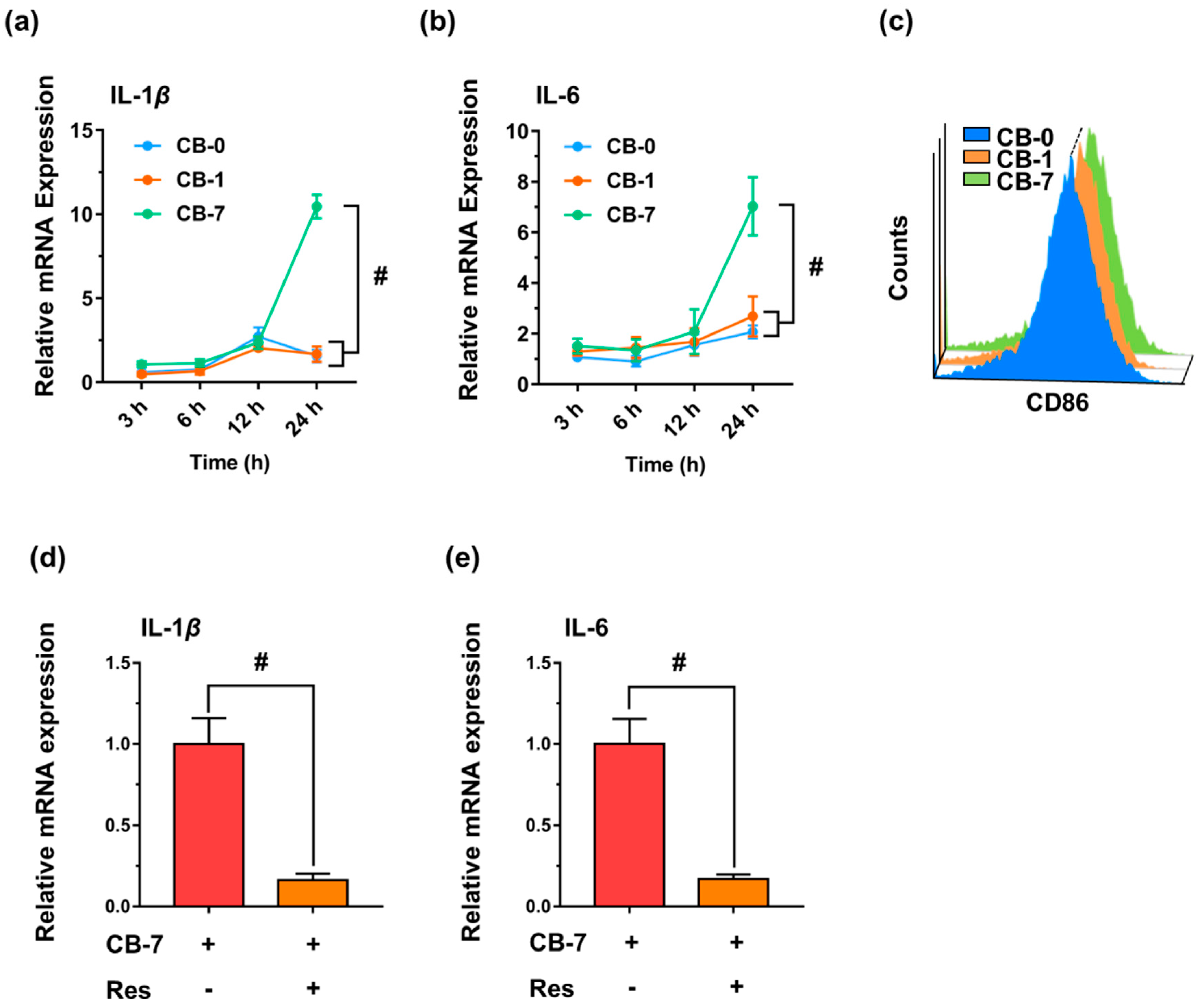
2.10. Real-Time Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Analysis of Gene Expression for Inflammatory Responses
2.11. Flow Cytometry Analysis of Macrophage Polarization
2.12. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Traina, G.; Bolzacchini, E.; Bonini, M.; Contini, D.; Mantecca, P.; Caimmi, S.M.E.; Licari, A.; Chiappini, E. Role of air pollutants mediated oxidative stress in respiratory diseases. Pediatr. Allergy Immunol. 2022, 33, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Lelieveld, J.; Evans, J.S.; Fnais, M.; Giannadaki, D.; Pozzer, A. The contribution of outdoor air pollution sources to premature mortality on a global scale. Nature 2015, 525, 367–371. [Google Scholar] [CrossRef]
- Cheng, J.; Tong, D.; Zhang, Q.; Liu, Y.; Lei, Y.; Yan, G.; Yan, L.; Yu, S.; Cui, R.Y.; Clarke, L.; et al. Pathways of China’s PM2.5 air quality 2015–2060 in the context of carbon neutrality. Natl. Sci. Rev. 2021, 8, nwab078. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Yao, Z.; Dai, Y.; Wang, C.H. A comparison of PM exposure related to emission hotspots in a hot and humid urban environment: Concentrations, compositions, respiratory deposition, and potential health risks. Sci. Total Environ. 2017, 599–600, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Matawle, J.L.; Pervez, S.; Shrivastava, A.; Tiwari, S.; Pant, P.; Deb, M.K.; Bisht, D.S.; Pervez, Y.F. PM2.5 pollution from household solid fuel burning practices in central India: 1. Impact on indoor air quality and associated health risks. Environ. Geochem. Health 2017, 39, 1045–1058. [Google Scholar] [CrossRef]
- Habre, R.; Zhou, H.; Eckel, S.P.; Enebish, T.; Fruin, S.; Bastain, T.; Rappaport, E.; Gilliland, F. Short-term effects of airport-associated ultrafine particle exposure on lung function and inflammation in adults with asthma. Environ. Int. 2018, 118, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Y.; Ma, J.; Ma, Q.; He, H. Heterogeneous reaction of SO2 with soot: The roles of relative humidity and surface composition of soot in surface sulfate formation. Atmos. Environ. 2017, 152, 465–476. [Google Scholar] [CrossRef]
- Kuang, Y.; Shang, J. Changes in light absorption by brown carbon in soot particles due to heterogeneous ozone aging in a smog chamber. Environ. Pollut. 2020, 266, 115273. [Google Scholar] [CrossRef]
- Li, Q.; Shang, J.; Liu, J.; Xu, W.; Feng, X.; Li, R.; Zhu, T. Physicochemical characteristics, oxidative capacities and cytotoxicities of sulfate-coated, 1,4-NQ-coated and ozone-aged black carbon particles. Atmos. Res. 2015, 153, 535–542. [Google Scholar] [CrossRef]
- Jiang, H.; Liu, Y.; Xie, Y.; Liu, J.; Chen, T.; Ma, Q.; He, H. Oxidation potential reduction of carbon nanomaterials during atmospheric-relevant aging: Role of surface coating. Environ. Sci. Technol. 2019, 53, 10454–10461. [Google Scholar] [CrossRef]
- Zhu, J.; Shang, J.; Chen, Y.; Kuang, Y.; Zhu, T. Reactive oxygen species-related inside-to-outside oxidation of soot particles tiggered by visible-light irradiation: Physicochemical property changes and oxidative potential enhancement. Environ. Sci. Technol. 2020, 54, 8558–8567. [Google Scholar] [CrossRef] [PubMed]
- Bates, J.T.; Fang, T.; Verma, V.; Zeng, L.; Weber, R.J.; Tolbert, P.E.; Abrams, J.Y.; Sarnat, S.E.; Klein, M.; Mulholland, J.A.; et al. Review of Acellular Assays of Ambient Particulate Matter Oxidative Potential: Methods and Relationships with Composition, Sources, and Health Effects. Environ. Sci. Technol. 2019, 53, 4003–4019. [Google Scholar] [CrossRef]
- Wei, S.; Qi, Y.; Ma, L.; Liu, Y.; Li, G.; Sang, N.; Liu, S.; Liu, Y. Ageing remarkably alters the toxicity of carbon black particles towards susceptible cells: Determined by differential changes of surface oxygen groups. Environ. Sci: Nano 2020, 7, 1633–1641. [Google Scholar] [CrossRef]
- Yu, Z.; Koppelman, G.H.; Hoek, G.; Kerckhoffs, J.; Vonk, J.M.; Vermeulen, R.; Gehring, U. Ultrafine particles, particle components and lung function at age 16 years: The PIAMA birth cohort study. Environ. Int. 2021, 157, 106792. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, D.; Li, H.; Wu, Y.; Cheng, Y.; Chen, Z.; Yin, G.; Wang, W.; Ge, Y.; Niu, Y.; et al. Exposure to ultrafine particles and oral flora, respiratory function, and biomarkers of inflammation: A panel study in children. Environ. Pollut. 2021, 273, 116489. [Google Scholar] [CrossRef] [PubMed]
- Samoli, E.; Rodopoulou, S.; Schneider, A.; Morawska, L.; Stafoggia, M.; Renzi, M.; Breitner, S.; Lanki, T.; Pickford, R.; Schikowski, T.; et al. Meta-analysis on short-term exposure to ambient ultrafine particles and respiratory morbidity. Eur. Respir. Rev. 2020, 29, 200116. [Google Scholar] [CrossRef]
- Coleman, D.L. Regulation of macrophage phagocytosis. Eur. J. Clin. Microbiol. 1986, 5, 1. [Google Scholar] [CrossRef]
- Michael, S.; Montag, M.; Dott, W. Pro-inflammatory effects and oxidative stress in lung macrophages and epithelial cells induced by ambient particulate matter. Environ. Pollut. 2013, 183, 19–29. [Google Scholar] [CrossRef]
- Colarusso, C.; De Falco, G.; Terlizzi, M.; Roviezzo, F.; Cerqua, I.; Sirignano, M.; Cirino, G.; Aquino, R.P.; Pinto, A.; D’Anna, A.; et al. The inhibition of caspase-1-does not revert particulate matter (PM)-induced lung immunesuppression in mice. Front. Immunol. 2019, 10, 1329. [Google Scholar] [CrossRef] [Green Version]
- Xia, T.; Kovochich, M.; Brant, J.; Hotze, M.; Sempf, J.; Oberley, T.; Sioutas, C.; Yeh, J.I.; Wiesner, M.R.; Nel, A.E. Comparison of the abilities of ambient and manufactured nanoparticles to induce cellular toxicity according to an oxidative stress paradigm. Nano Lett. 2006, 6, 1794–1807. [Google Scholar] [CrossRef]
- Saleh, Y.; Antherieu, S.; Dusautoir, R.; Alleman, L.Y.; Sotty, J.; De Sousa, C.; Platel, A.; Perdrix, E.; Riffault, V.; Fronval, I.; et al. Exposure to atmospheric ultrafine particles induces severe lung inflammatory response and tissue remodeling in mice. Int. J. Environ. Res. Public Health 2019, 16, 1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C. The immune effects of naturally occurring and synthetic nanoparticles. J. Autoimmun. 2010, 34, J234–J246. [Google Scholar] [CrossRef] [PubMed]
- Elsabahy, M.; Wooley, K.L. Cytokines as biomarkers of nanoparticle immunotoxicity. Chem. Soc. Rev. 2013, 42, 5552–5576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotty, J.; Kluza, J.; De Sousa, C.; Tardivel, M.; Antherieu, S.; Alleman, L.Y.; Canivet, L.; Perdrix, E.; Loyens, A.; Marchetti, P.; et al. Mitochondrial alterations triggered by repeated exposure to fine (PM2.5–0.18) and quasi-ultrafine (PM0.18) fractions of ambient particulate matter. Environ. Int. 2020, 142, 105830. [Google Scholar] [CrossRef]
- Facciola, A.; Visalli, G.; Pruiti Ciarello, M.; Di Pietro, A. Newly emerging airborne pollutants: Current knowledge of health impact of micro and nanoplastics. Int. J. Environ. Res. Public Health 2021, 18, 2997. [Google Scholar] [CrossRef]
- Holland, N.A.; Fraiser, C.R.; Sloan, R.C., 3rd; Devlin, R.B.; Brown, D.A.; Wingard, C.J. Ultrafine particulate matter increases cardiac Ischemia/reperfusion injury via mitochondrial permeability transition pore. Cardiovasc. Toxicol. 2017, 17, 441–450. [Google Scholar] [CrossRef]
- Wang, S.; Chen, Y.; Hong, W.; Li, B.; Zhou, Y.; Ran, P. Chronic exposure to biomass ambient particulate matter triggers alveolar macrophage polarization and activation in the rat lung. J. Cell. Mol. Med. 2022, 26, 1156–1168. [Google Scholar] [CrossRef]
- Yuan, X.; Zhang, X.; Sun, L.; Wei, Y.; Wei, X. Cellular toxicity and immunological effects of carbon-based nanomaterials. Part Fibre Toxicol. 2019, 16, 18. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, V.S.; Harding, J.N.; You, D.; Cormier, S.A. Exposure to combustion derived particulate matter exacerbates influenza infection in neonatal mice by inhibiting IL22 production. Part. Fibre Toxicol. 2021, 18, 43. [Google Scholar] [CrossRef]
- Wu, Y.; Guo, Y.; Song, H.; Liu, W.; Yang, Y.; Liu, Y.; Sang, N.; Zuo, Y.Y.; Liu, S. Oxygen content determines the bio-reactivity and toxicity profiles of carbon black particles. Ecotoxicol. Environ. Saf. 2018, 150, 207–214. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Smith, W.; Hao, D.; He, B.; Kong, L. M1 and M2 macrophage polarization and potentially therapeutic naturally occurring compounds. Int. Immunopharmacol. 2019, 70, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef]
- Zhou, D.; Huang, C.; Lin, Z.; Zhan, S.; Kong, L.; Fang, C.; Li, J. Macrophage polarization and function with emphasis on the evolving roles of coordinated regulation of cellular signaling pathways. Cell. Signal. 2014, 26, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Lawal, A.O. Air particulate matter induced oxidative stress and inflammation in cardiovascular disease and atherosclerosis: The role of Nrf2 and AhR-mediated pathways. Toxicol. Lett. 2017, 270, 88–95. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Miethe, S.; Schindler, V.; Alhamdan, F.; Garn, H. Role of airway epithelial cells in the development of different asthma phenotypes. Cell. Signal. 2020, 69, 109523. [Google Scholar] [CrossRef]
- Wu, L.; Li, D.; Qin, L.; Wang, Q.; Saito, Y.; Sara, R.; Fan, J. Growth hormone secretagogue receptor deficiency promotes lung cancer growth by affecting the Th17/Treg balance. Ann. Transl. Med. 2021, 9, 1696. [Google Scholar] [CrossRef]
- Harmon, A.C.; Noel, A.; Subramanian, B.; Perveen, Z.; Dugas, T.R. Inhalation of particulate matter containing free radicals leads to decreased vascular responsiveness associated with an altered pulmonary function. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H667–H683. [Google Scholar] [CrossRef]
- Raaschou-Nielsen, O.; Beelen, R.; Wang, M.; Hoek, G.; Andersen, Z.J.; Hoffmann, B.; Stafoggia, M.; Samoli, E.; Weinmayr, G.; Dimakopoulou, K.; et al. Particulate matter air pollution components and risk for lung cancer. Environ. Int. 2016, 87, 66–73. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Wang, S.; Ma, J.; Xu, M.; Gao, M.; Liu, R.; Chen, W.; Liu, S. Reduction of graphene oxide alters its cyto-compatibility towards primary and immortalized macrophages. Nanoscale 2018, 10, 14637–14650. [Google Scholar] [CrossRef]
- Ma, J.; Guo, A.; Wang, S.; Man, S.; Zhang, Y.; Liu, S.; Liu, Y. From the lung to the knee joint: Toxicity evaluation of carbon black nanoparticles on macrophages and chondrocytes. J. Hazard. Mater. 2018, 353, 329–339. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Ichinose, T.; Yoshida, Y.; Arashidani, K.; Yoshida, S.; Takano, H.; Sun, G.; Shibamoto, T. Urban PM2.5 exacerbates allergic inflammation in the murine lung via a TLR2/TLR4/MyD88-signaling pathway. Sci. Rep. 2017, 7, 11027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.; Zhou, L.; Xie, H.; Pei, Z.; Zhang, M.; Wu, M.; Zhang, S.; Wang, L.; Zhao, C.; Shi, L.; et al. Pulmonary toxicities from a 90-day chronic inhalation study with carbon black nanoparticles in rats related to the systemical immune effects. Int. J. Nanomed. 2019, 14, 2995–3013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liu, C.; Ma, J.; Ma, Q.; He, H. Structural and hygroscopic changes of soot during heterogeneous reaction with O(3). Phys. Chem. Chem. Phys. 2010, 12, 10896–10903. [Google Scholar] [CrossRef]
- Du, T.; Adeleye, A.S.; Zhang, T.; Yang, N.; Hao, R.; Li, Y.; Song, W.; Chen, W. Effects of ozone and produced hydroxyl radicals on the transformation of graphene oxide in aqueous media. Environ. Sci. Nano 2019, 6, 2484–2494. [Google Scholar] [CrossRef]
- Miyata, R.; van Eeden, S.F. The innate and adaptive immune response induced by alveolar macrophages exposed to ambient particulate matter. Toxicol. Appl. Pharmacol. 2011, 257, 209–226. [Google Scholar] [CrossRef]
- Wang, B.; Li, K.; Jin, W.; Lu, Y.; Zhang, Y.; Shen, G.; Wang, R.; Shen, H.; Li, W.; Huang, Y.; et al. Properties and inflammatory effects of various size fractions of ambient particulate matter from Beijing on A549 and J774A.1 cells. Environ. Sci. Technol. 2013, 47, 10583–10590. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Chen, W.; Liu, F.; Liu, J.; Zhang, T.; Chen, W. Aggregation morphology is a key factor determining protein adsorption on graphene oxide and reduced graphene oxide nanomaterials. Environ. Sci. Nano 2019, 6, 1303–1309. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, Y.; Xia, T.; Xu, A.; Liu, S.; Chen, W. The biotransformation of graphene oxide in lung fluids significantly alters its inherent properties and bioactivities toward immune cells. NPG Asia Mater. 2018, 10, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Donadon, M.; Torzilli, G.; Cortese, N.; Soldani, C.; Di Tommaso, L.; Franceschini, B.; Carriero, R.; Barbagallo, M.; Rigamonti, A.; Anselmo, A.; et al. Macrophage morphology correlates with single-cell diversity and prognosis in colorectal liver metastasis. J. Exp. Med. 2020, 217, e20191847. [Google Scholar] [CrossRef]
- Qu, G.; Liu, S.; Zhang, S.; Wang, L.; Wang, X.; Sun, B.; Yin, N.; Gao, X.; Xia, T.; Chen, J.-J.; et al. Graphene oxide induces toll-like receptor 4 (TLR4)-dependent necrosis in macrophages. ACS Nano 2013, 7, 5732–5745. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, Y.; Yang, H.; Lu, Y.; Li, L. Oxidized low-density lipoprotein induces apoptosis in cultured neonatal rat cardiomyocytes by modulating the TLR4/NF-kappaB pathway. Sci. Rep. 2016, 6, 27866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.S.; Ryu, M.H.; Carlsten, C. Ultrafine particles: Unique physicochemical properties relevant to health and disease. Exp. Mol. Med. 2020, 52, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Eapen, M.S.; Hansbro, P.M.; McAlinden, K.; Kim, R.Y.; Ward, C.; Hackett, T.L.; Walters, E.H.; Sohal, S.S. Abnormal M1/M2 macrophage phenotype profiles in the small airway wall and lumen in smokers and chronic obstructive pulmonary disease (COPD). Sci. Rep. 2017, 7, 13392. [Google Scholar] [CrossRef]
- Jiao, Y.; Zhang, T.; Zhang, C.; Ji, H.; Tong, X.; Xia, R.; Wang, W.; Ma, Z.; Shi, X. Exosomal miR-30d-5p of neutrophils induces M1 macrophage polarization and primes macrophage pyroptosis in sepsis-related acute lung injury. Crit. Care 2021, 25, 356. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Qiu, B. The role of immune cells in pulmonary hypertension: Focusing on macrophages. Hum. Immunol. 2022, 83, 153–163. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, X.; Chen, Y.; Ma, L.; Liu, Y.; Qi, Y.; Liu, S. Ageing Significantly Alters the Physicochemical Properties and Associated Cytotoxicity Profiles of Ultrafine Particulate Matters towards Macrophages. Antioxidants 2022, 11, 754. https://doi.org/10.3390/antiox11040754
Yan X, Chen Y, Ma L, Liu Y, Qi Y, Liu S. Ageing Significantly Alters the Physicochemical Properties and Associated Cytotoxicity Profiles of Ultrafine Particulate Matters towards Macrophages. Antioxidants. 2022; 11(4):754. https://doi.org/10.3390/antiox11040754
Chicago/Turabian StyleYan, Xu, Yucai Chen, Li Ma, Yongchun Liu, Yu Qi, and Sijin Liu. 2022. "Ageing Significantly Alters the Physicochemical Properties and Associated Cytotoxicity Profiles of Ultrafine Particulate Matters towards Macrophages" Antioxidants 11, no. 4: 754. https://doi.org/10.3390/antiox11040754