
Abiotic Stresses Elicitation Potentiates the Productiveness of Cardoon Calli as Bio-Factories for Specialized Metabolites Production

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Callus Induction
2.2. Experimental Design and Stress Treatments
Chilling and Salt Stress
2.3. Growth Evaluation
2.4. Ultrasound Assisted Extraction of Polyphenols and UHPLC-Q-Orbitrap HRMS Analysis
2.5. RNA Extraction and Expression Analysis
2.6. Antioxidant Activity: ABTS, DPPH, and FRAP Assays
2.7. Lipid Peroxidation Measurement
2.8. Human Cell Cultures
2.9. MTT Assay
2.10. Hydrating Power
2.11. Pro Collagene I Production
2.12. Statistical Analysis
3. Results
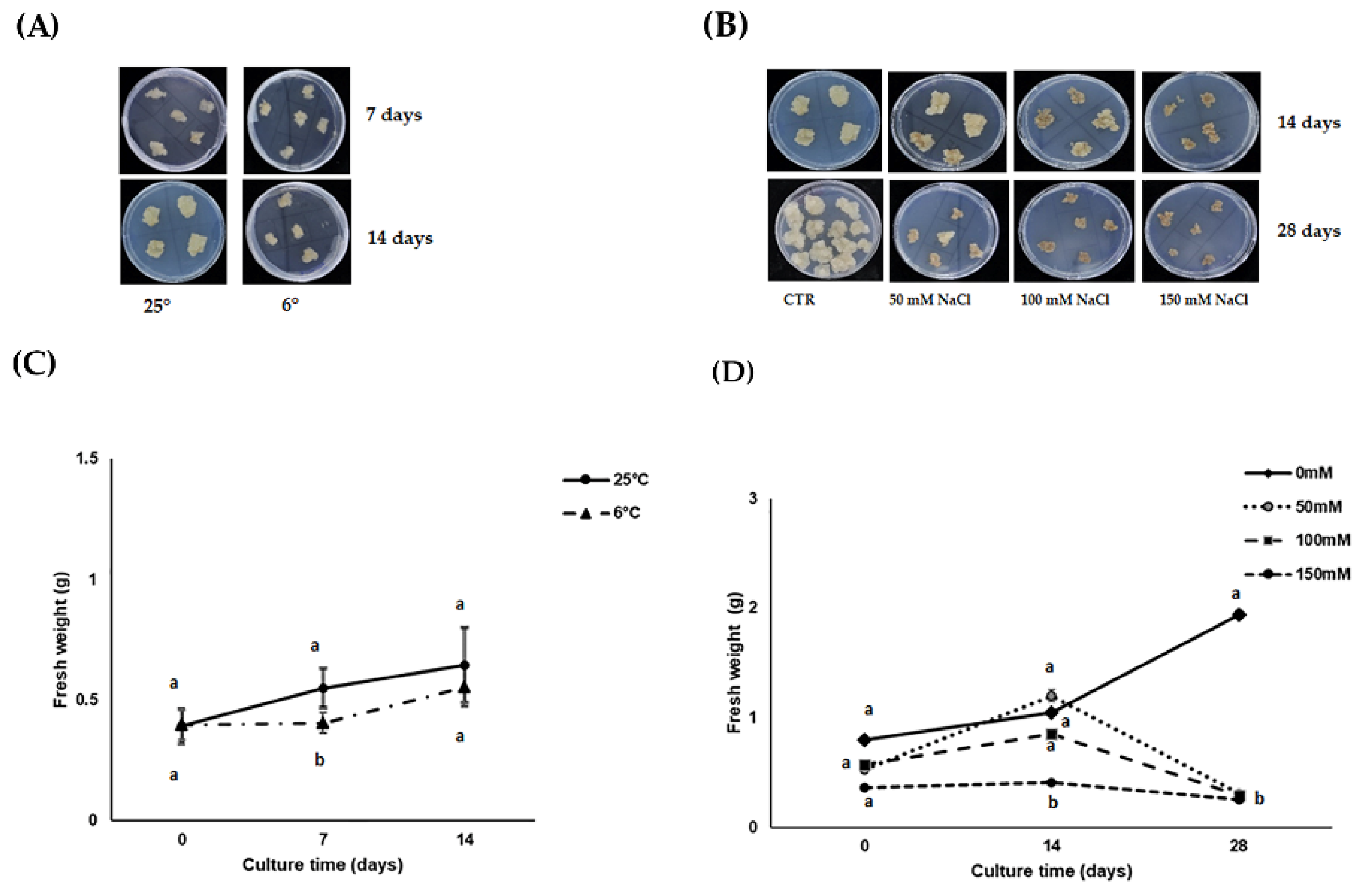
3.1. Calli Growth under Stress Conditions
3.2. Chilling and Salt Stresses Elicit Phenylpropanoid Accumulation
3.2.1. Metabolic Changes
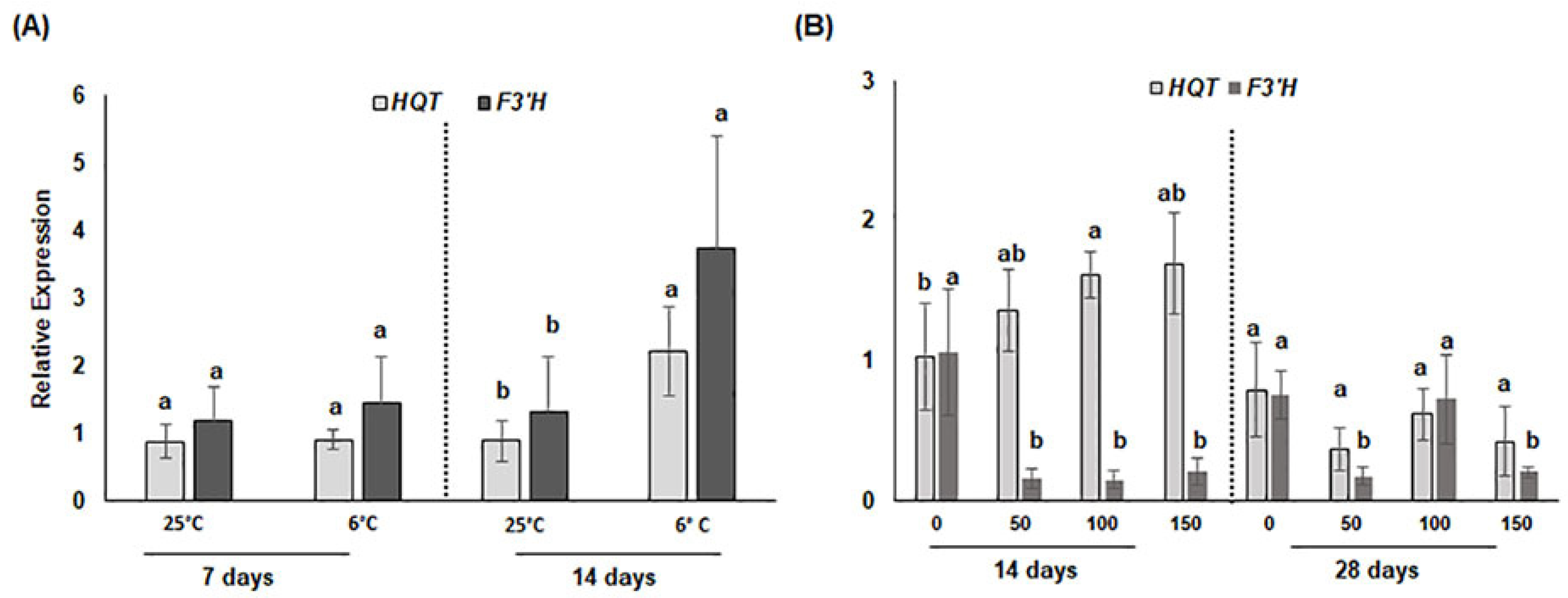
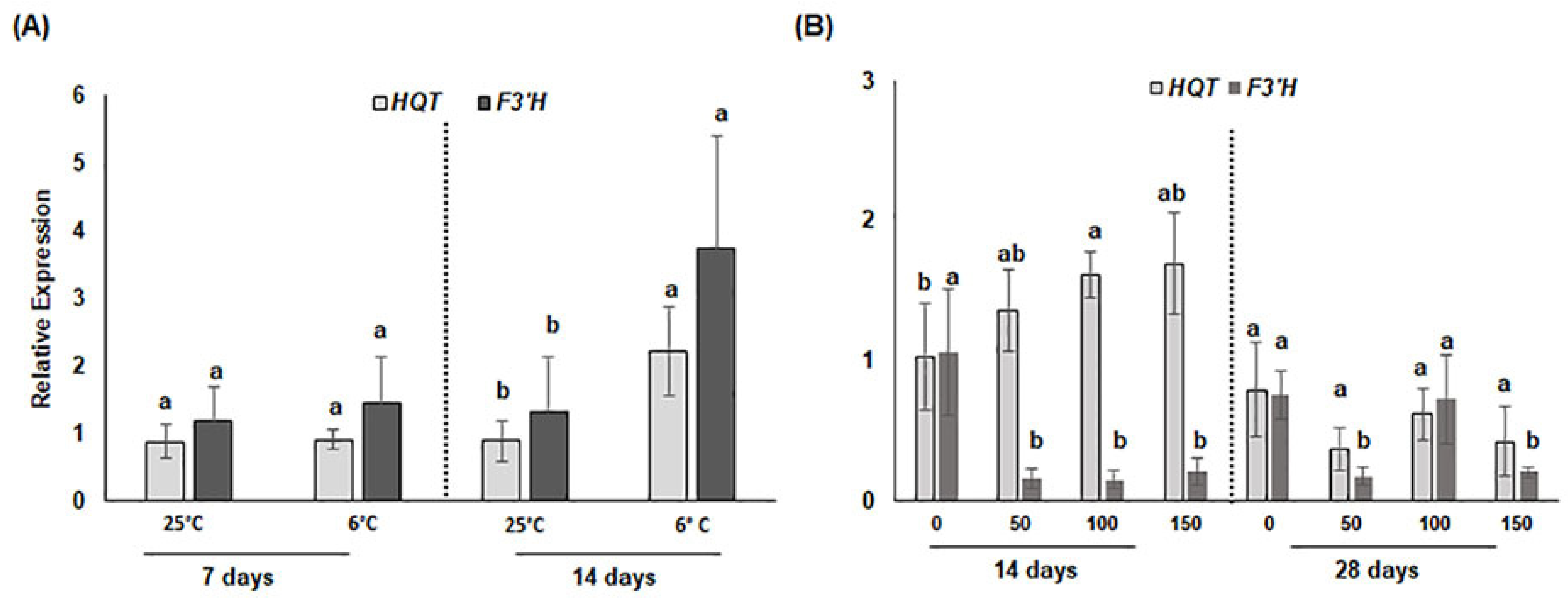
3.2.2. Transcriptional Changes
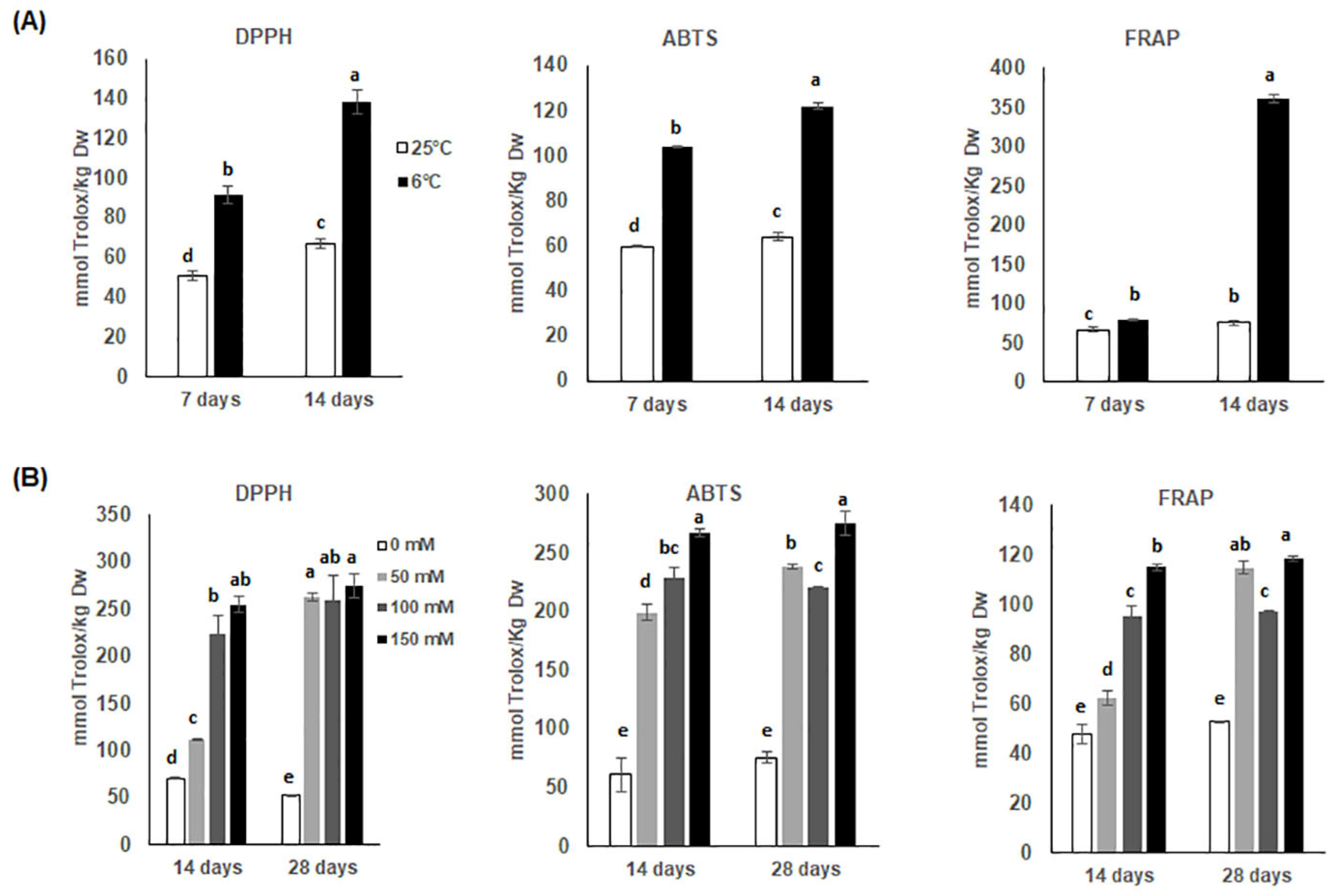
3.3. Changes in Antioxidant Activities
3.4. Effects of Stresses on Lipid Peroxidation of Cardoon Calli
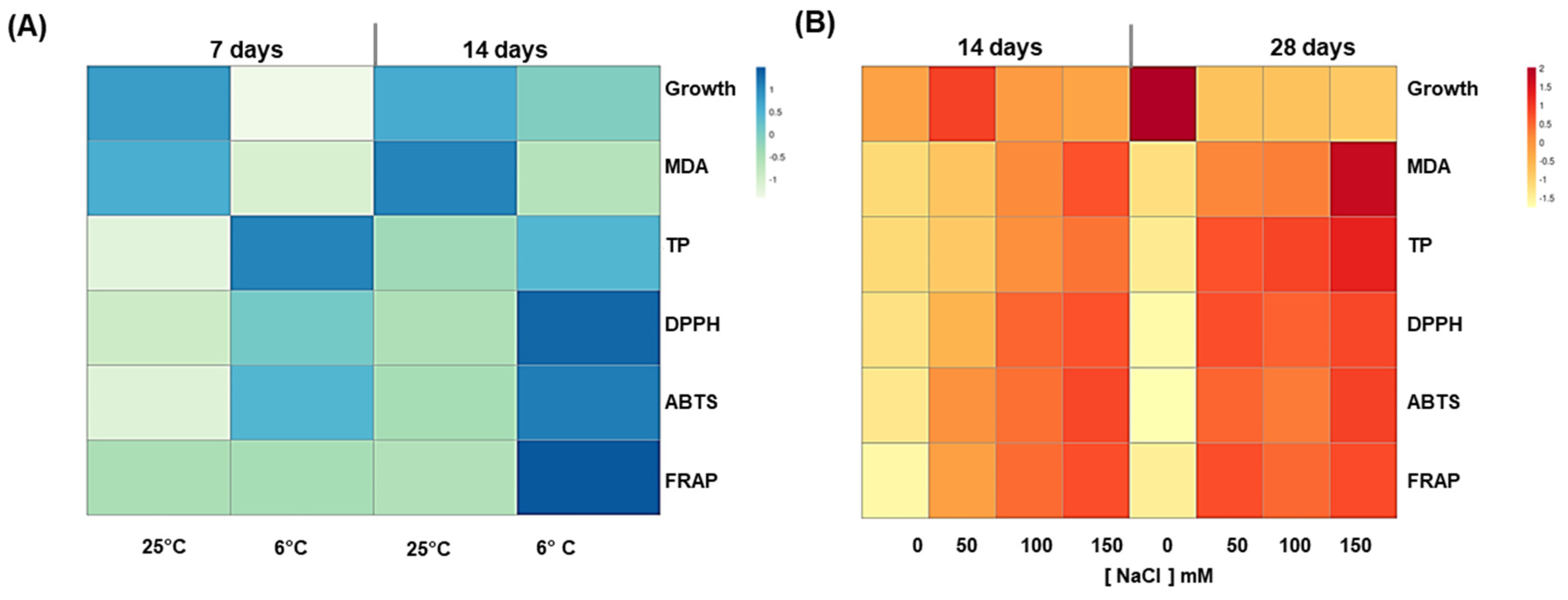
3.5. Evaluation of the Optimal Elicitation Strategy
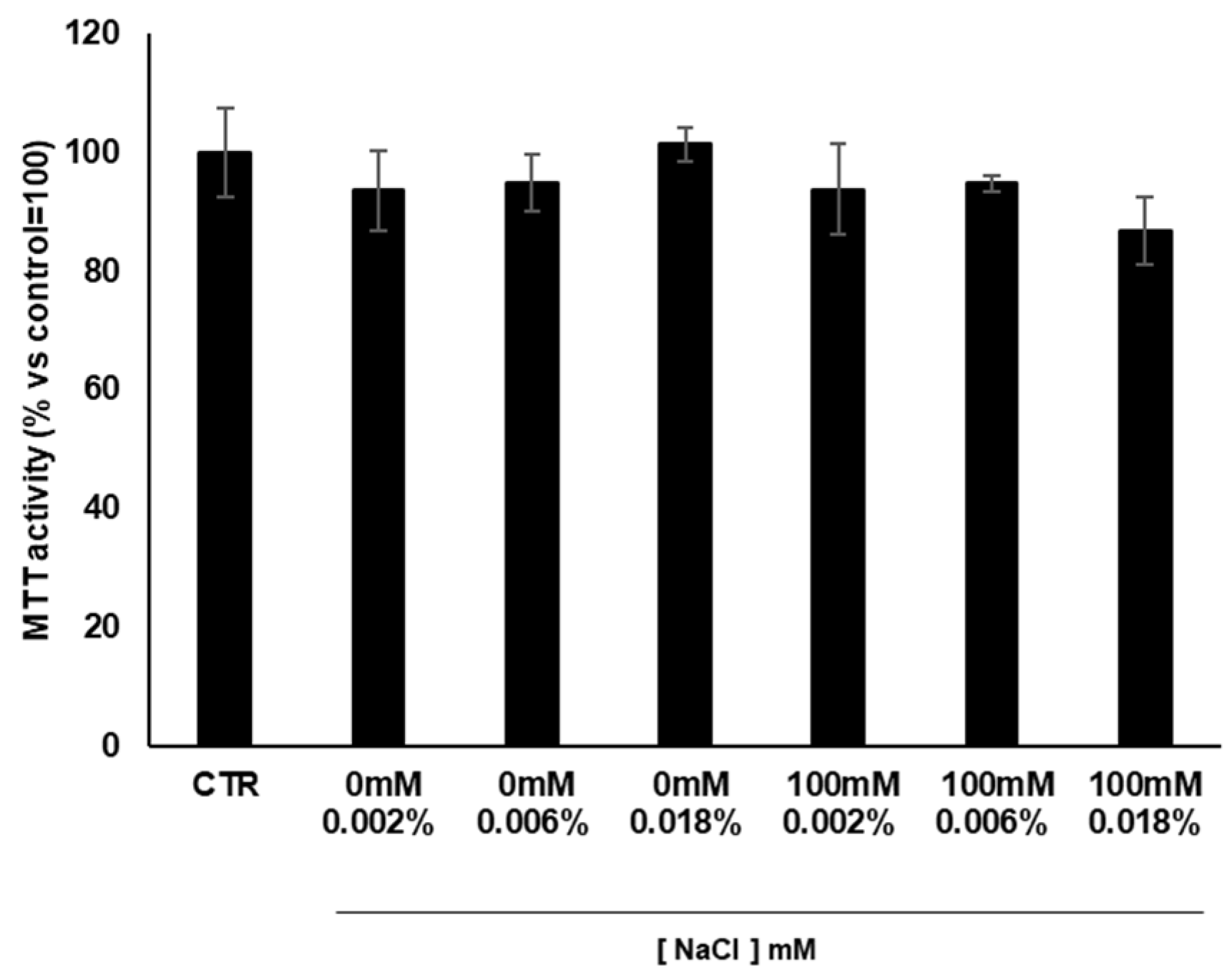
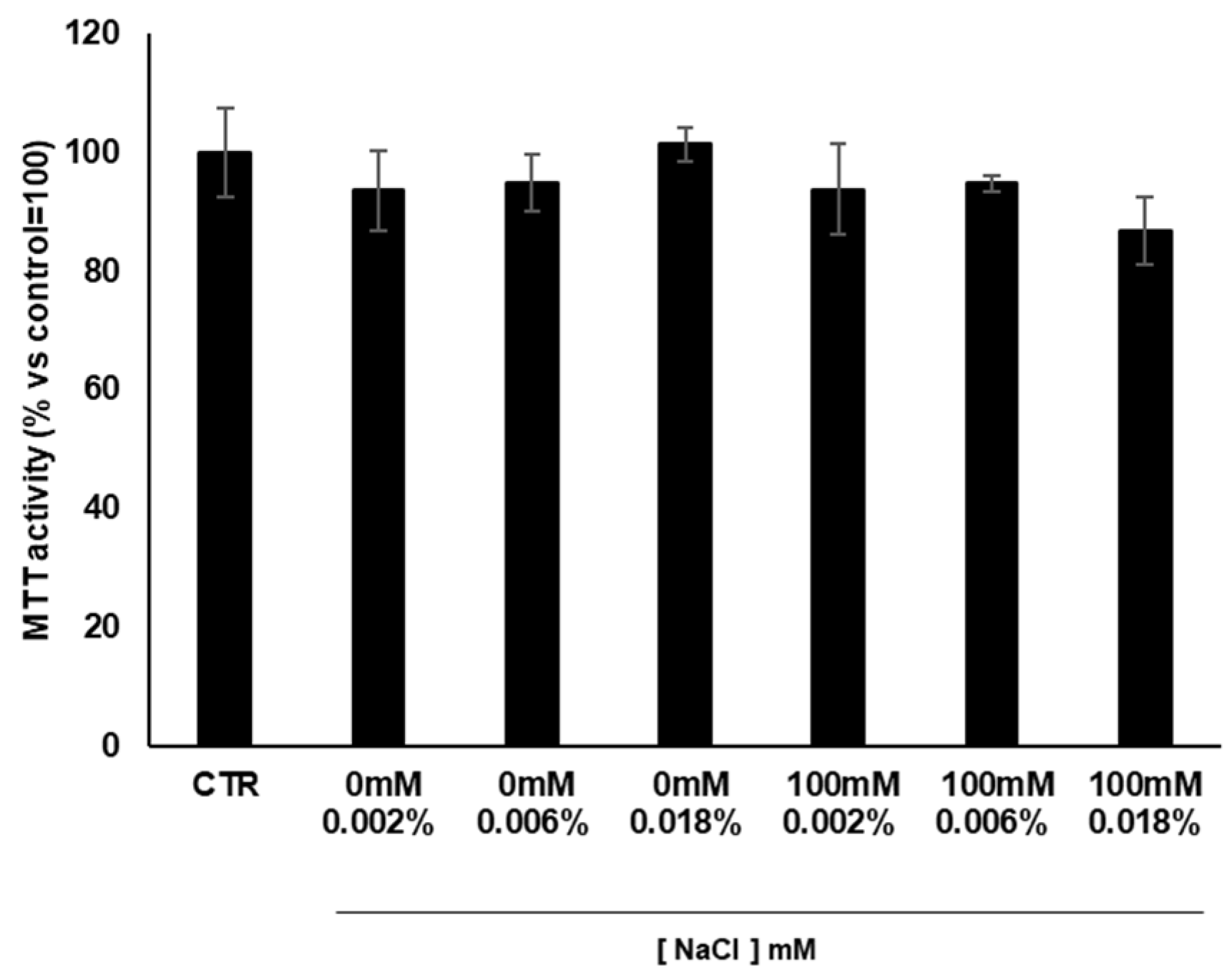
3.6. Cytotoxicity of Cardoon Calli Extracts
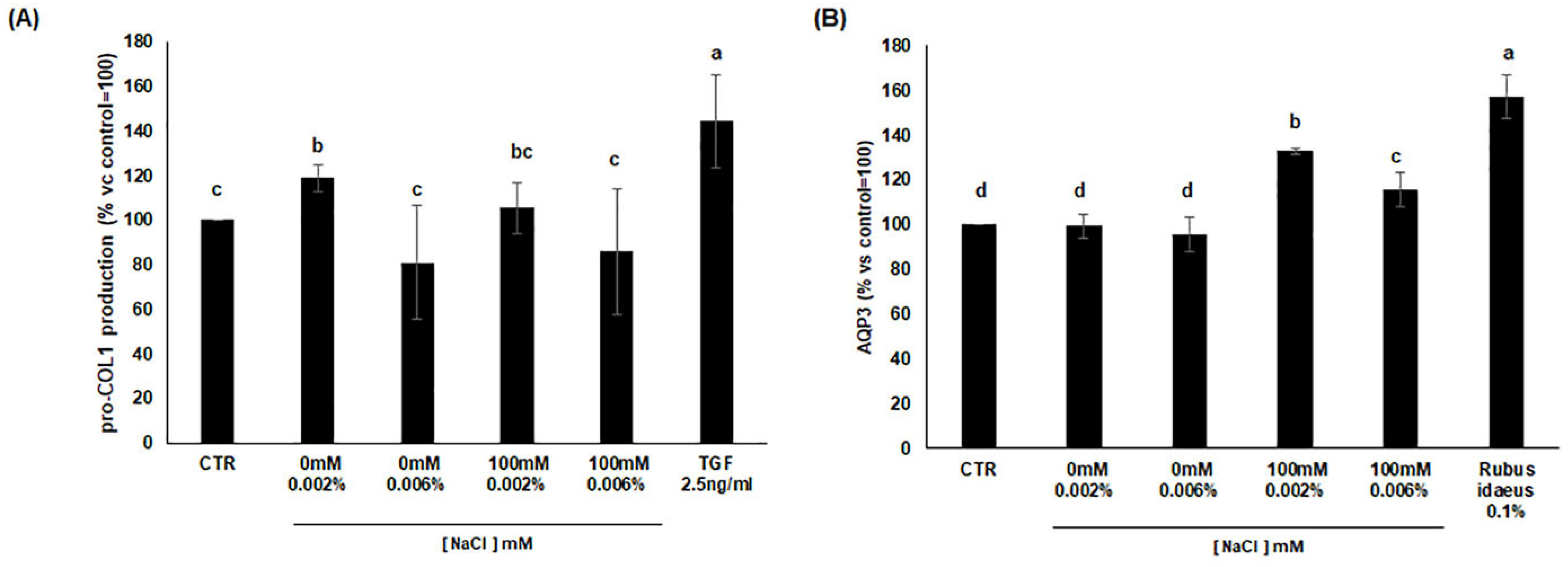
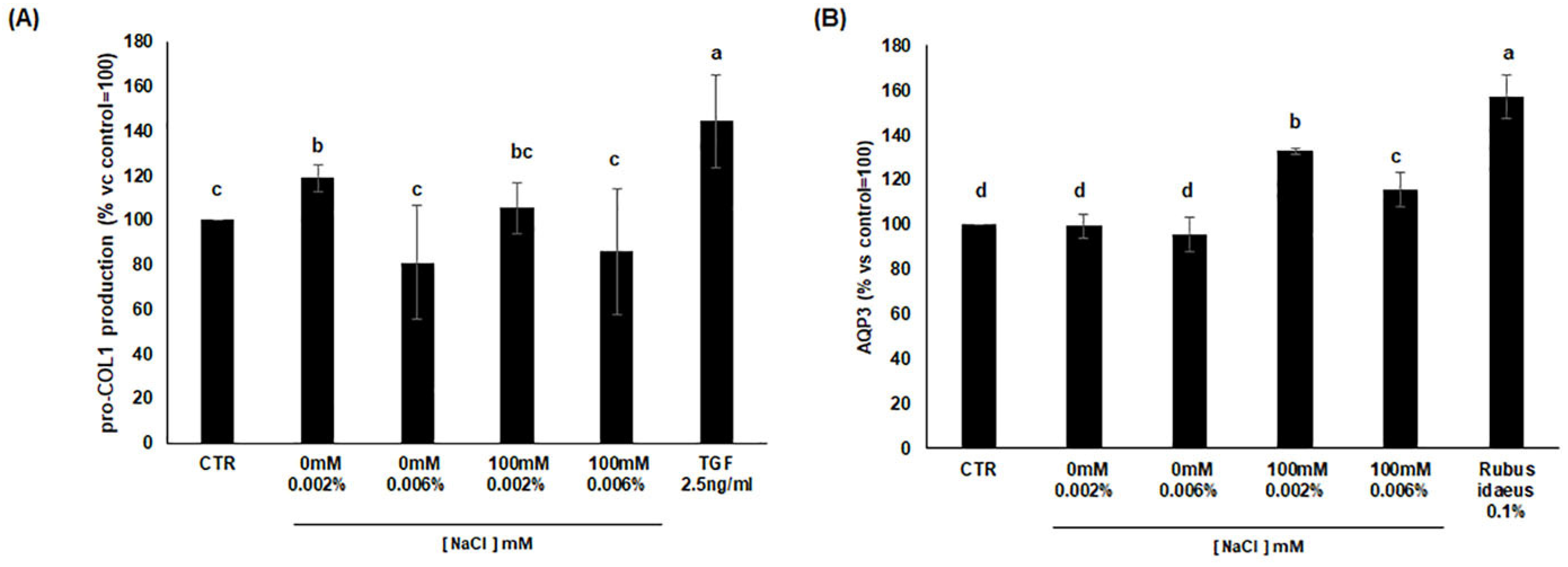
3.7. Analysis of Procollagen I and AQP3 Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balestrini, R.; Brunetti, C.; Cammareri, M.; Caretto, S.; Cavallaro, V.; Cominelli, E.; De Palma, M.; Docimo, T.; Giovinazzo, G.; Grandillo, S.; et al. Strategies to modulate specialized metabolism in mediterranean crops: From molecular aspects to field. Int. J. Mol. Sci. 2021, 22, 2887. [Google Scholar] [CrossRef] [PubMed]
- D’Amelia, V.; Docimo, T.; Crocoll, C.; Rigano, M.M. Specialized metabolites and valuable molecules in crop and medicinal plants: The evolution of their use and strategies for their production. Genes 2021, 12, 936. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Farkya, S.; Srivastava, A.K.; Bisaria, V.S. Bioprocess considerations for production of secondary metabolites by plant cell suspension cultures. Biotechnol. Bioprocess Eng. 2002, 7, 138–149. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Kreis, W. Exploiting plant cell culture for natural product formation. J. Appl. Bot. Food Qual. 2019, 92, 216–225. [Google Scholar] [CrossRef]
- Baenas, N.; García-Viguera, C.; Moreno, D. Elicitation: A tool for enriching the bioactive composition of foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef] [Green Version]
- Linh, T.M.; Mai, N.C.; Hoe, P.T.; Ngoc, N.T.; Thao, P.T.H.; Ban, N.K.; Van, N.T. Development of a cell suspension culture system for promoting alkaloid and vinca alkaloid biosynthesis using endophytic fungi isolated from local Catharanthus roseus. Plants 2021, 10, 672. [Google Scholar] [CrossRef]
- Gabotti, D.; Locatelli, F.; Cusano, E.; Baldoni, E.; Genga, A.; Pucci, L.; Consonni, R.; Mattana, M. Cell suspensions of Cannabis sativa (var. futura): Effect of elicitation on metabolite content and antioxidant activity. Molecules 2019, 24, 4056. [Google Scholar] [CrossRef] [Green Version]
- Wróbel, T.; Dreger, M.; Wielgus, K.; Słomski, R. The application of plant in vitro cultures in cannabinoid production. Biotechnol. Lett. 2018, 40, 445–454. [Google Scholar] [CrossRef]
- Farag, M.A.; Huhman, D.V.; Dixon, R.A.; Sumner, L.W. Metabolomics reveals novel pathways and differential mechanistic and elicitor-specific responses in phenylpropanoid and isoflavonoid biosynthesis in Medicago truncatula cell cultures. Plant Physiol. 2008, 146, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Deavours, B.E.; de Fáltima, Â.; Naoumkina, M.; Dixon, R.A.; Sumner, L.W. Integrated metabolite and transcript profiling identify a biosynthetic mechanism for hispidol in Medicago truncatula cell cultures. Plant Physiol. 2009, 151, 1096–1113. [Google Scholar] [CrossRef] [Green Version]
- Onrubia, M.; Cusido, R.M.; Ramirez, K.; Hernandez-Vazquez, L.; Moyano, E.; Bonfill, M.; Palazon, J. Bioprocessing of plant in vitro systems for the mass production of pharmaceutically important metabolites: Paclitaxel and its derivatives. Curr. Med. Chem. 2013, 20, 880–891. [Google Scholar] [CrossRef]
- Yukimune, Y.; Tabata, H.; Higashi, Y.; Hara, Y. Methyl jasmonate-induced overproduction of paclitaxel and baccatin III in Taxus cell suspension cultures. Nat. Biotechnol. 1996, 14, 1129–1132. [Google Scholar] [CrossRef]
- Graziani, G.; Docimo, T.; De Palma, M.; Sparvoli, F.; Izzo, L.; Tucci, M.; Ritieni, A. Changes in phenolics and fatty acids composition and related gene expression during the development from seed to leaves of three cultivated cardoon genotypes. Antioxidants 2020, 9, 1096. [Google Scholar] [CrossRef]
- Docimo, T.; De Stefano, R.; Cappetta, E.; Piccinelli, A.L.; Celano, R.; De Palma, M.; Tucci, M. Physiological, biochemical, and metabolic responses to short and prolonged saline stress in two cultivated cardoon genotypes. Plants 2020, 9, 554. [Google Scholar] [CrossRef]
- Fratianni, F.; Tucci, M.; De Palma, M.; Pepe, R.; Nazzaro, F. Polyphenolic composition in different parts of some cultivars of globe artichoke (Cynara cardunculus L. var. Scolymus (L.) Fiori). Food Chem. 2007, 104, 1282–1286. [Google Scholar] [CrossRef]
- De Palma, M.; Fratianni, F.; Nazzaro, F.; Tucci, M. Isolation and functional characterization of a novel gene coding for flavonoid 3’-hydroxylase from globe artichoke. Biol. Plant. 2014, 58, 445–455. [Google Scholar] [CrossRef]
- Barbosa, C.H.; Andrade, M.A.; Vilarinho, F.; Castanheira, I.; Fernando, A.L.; Loizzo, M.R.; Sanches Silva, A. A new insight on cardoon: Exploring new uses besides cheese making with a view to zero waste. Foods 2020, 9, 564. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Pereira, C.; Tzortzakis, N.; Barros, L.; Ferreira, I.C.F.R. Nutritional value and bioactive compounds characterization of plant parts from Cynara cardunculus L. (Asteraceae) cultivated in central Greece. Front. Plant Sci. 2018, 1, 459. [Google Scholar] [CrossRef] [Green Version]
- Gominho, J.; Curt, M.D.; Lourenço, A.; Fernández, J.; Pereira, H. Cynara cardunculus L. as a biomass and multi-purpose crop: A review of 30 years of research. Biomass Bioenergy 2018, 109, 257–275. [Google Scholar] [CrossRef]
- Oliet, M.; Gilarranz, M.A.; Domínguez, J.C.; Alonso, M.V.; Rodríguez, F. Ethanol-based pulping from Cynara cardunculus L. J. Chem. Technol. Biotechnol. 2005, 80, 746–753. [Google Scholar] [CrossRef]
- Grammelis, P.; Malliopoulou, A.; Basinas, P.; Nicholas, G. Cultivation and characterization of Cynara cardunculus for solid biofuels production in the mediterranean region. Int. J. Mol. Sci. 2008, 9, 1241–1258. [Google Scholar] [CrossRef] [PubMed]
- Dufour, J.; Arsuaga, J.; Moreno, J.; Torrealba, H.; Camacho, J. Comparative life cycle assessment of biodiesel production from cardoon (Cynara cardunculus) and rapeseed oil obtained under Spanish conditions. Energy Fuels 2013, 27, 5280–5286. [Google Scholar] [CrossRef]
- Paolo, D.; Locatelli, F.; Cominelli, E.; Pirona, R.; Pozzo, S.; Graziani, G.; Ritieni, A.; De Palma, M.; Docimo, T.; Tucci, M.; et al. Towards a cardoon (Cynara cardunculus var. altilis)-based biorefinery: A case study of improved cell cultures via genetic modulation of the phenylpropanoid pathway. Int. J. Mol. Sci. 2021, 22, 1978. [Google Scholar] [CrossRef]
- Golkar, P.; Taghizadeh, M.; Noormohammadi, A. Effects of sodium alginate elicitation on secondary metabolites and antioxidant activity of safflower genotypes under in vitro salinity stress. In Vitro Cell. Dev. Biol. Plant 2019, 55, 527–538. [Google Scholar] [CrossRef]
- Acquadro, A.; Portis, E.; Valentino, D.; Barchi, L.; Lanteri, S. “Mind the Gap”: Hi-C technology boosts contiguity of the globe artichoke genome in low-recombination regions. G3 2020, 10, 3557–3564. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Quantification strategies in real-time PCR. In A-Z of Quantitative PCR.; Bustin, S.A., Ed.; International University Line (IUL): La Jolla, CA, USA, 2004; Volume 3, pp. 87–112. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric reducing ability of plasma (FRAP) as a measure of “Antioxidant Power”: The Frap assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Chen, T.S.; Liou, S.Y.; Wu, H.C.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Chang, Y.L. New analytical method for investigating the antioxidant power of food extracts on the basis of their electron-donating ability: Comparison to the ferric reducing/antioxidant power (FRAP) assay. J. Agric. Food Chem. 2010, 58, 8477–8480. [Google Scholar] [CrossRef]
- Tito, A.; Bimonte, M.; Carola, A.; De Lucia, A.; Barbulova, A.; Tortora, A.; Colucci, G.; Apone, F. An oil-soluble extract of Rubus idaeus cells enhances hydration and water homeostasis in skin cells. Int. J. Cosmet. Sci. 2015, 37, 588–594. [Google Scholar] [CrossRef]
- Eibl, R.; Meier, P.; Stutz, I.; Schildberger, D.; Hühn, T.; Eibl, D. Plant cell culture technology in the cosmetics and food industries: Current state and future trends. Appl. Microbiol. Biotechnol. 2018, 102, 8661–8675. [Google Scholar] [CrossRef] [Green Version]
- Trantas, E.A.; Koffas, M.A.G.; Xu, P.; Ververidis, F. When plants produce not enough or at all: Metabolic engineering of flavonoids in microbial hosts. Front. Plant Sci. 2015, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Mulder-Krieger, T.H.; Verpoorte, R.; Svendsen, A.B.; Scheffer, J.J.C.; Baerheim, A. Production of essential oils and flavours in plant cell and tissue cultures. A review. Plant Cell. Tissue Organ Cult. 1988, 13, 85–154. [Google Scholar] [CrossRef]
- Smetanska, I. Production of secondary metabolites using plant cell cultures. Adv. Biochem. Eng. Biotechnol. 2008, 111, 187–228. [Google Scholar] [CrossRef]
- Wilson, S.A.; Roberts, S.C. Recent advances towards development and commercialization of plant cell culture processes for the synthesis of biomolecules. Plant Biotechnol. J. 2012, 10, 249–268. [Google Scholar] [CrossRef]
- Mulabagal, V.; Tsay, H. Plant Cell Cultures—An alternative and efficient source for the production of biologically important secondary metabolites. Int. J. Appl. Sci. Eng. 2004, 2, 29–48. [Google Scholar]
- Rischer, H.; Szilvay, G.R.; Oksman-Caldentey, K.M. Cellular agriculture—Industrial biotechnology for food and materials. Curr. Opin. Biotechnol. 2020, 61, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, M.; Langellotti, A.L.; Russo, G.L.; Baselice, M.; Donadio, A.; Ritieni, A.; Graziani, G.; Masi, P. Use of different organic carbon sources in Cynara cardunculus cells: Effects on biomass productivity and secondary metabolites. Plants 2022, 11, 701. [Google Scholar] [CrossRef] [PubMed]
- D’Amelia, V.; Villano, C.; Aversano, R. Emerging genetic technologies to improve crop productivity. In Encyclopedia of Food Security and Sustainability; Elsevier: Amsterdam, The Netherlands, 2019; pp. 152–158. [Google Scholar]
- Papazoglou, E.G. Responses of Cynara cardunculus L. to single and combined cadmium and nickel treatment conditions. Ecotoxicol. Environ. Saf. 2011, 74, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, C.; Toscano, V.; Pappalardo, H.; Genovese, C.; Raccuia, S.A. The potential of Cynara cardunculus L. for phytoremediation of heavy metal in contaminated soils. Acta Hortic. 2016, 1147, 127–138. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Svecova, E.; Rea, E.; Lucini, L. Effects of saline stress on mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon genotypes grown in floating system. J. Sci. Food Agric. 2013, 93, 1119–1127. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Lattanzio, V.; Caretto, S.; Linsalata, V.; Colella, G.; Mita, G. Signal transduction in artichoke [Cynara cardunculus L. subsp. Scolymus (L.) Hayek] callus and cell suspension cultures under nutritional stress. Plant Physiol. Biochem. 2018, 127, 97–103. [Google Scholar] [CrossRef]
- Sák, M.; Dokupilová, I.; Kaňuková, Š.; Mrkvová, M.; Mihálik, D.; Hauptvogel, P.; Kraic, J. Biotic and abiotic elicitors of stilbenes production in Vitis vinifera L. cell culture. Plants 2021, 10, 490. [Google Scholar] [CrossRef]
- Zhu, J.-K. Cell signaling under salt, water and cold stresses. Curr. Opin. Plant Biol. 2001, 4, 401–406. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a multifaceted signalling molecule in plant responses to abiotic stress: Understanding the physiological mechanisms. Plant Biol. 2022, 24, 227–239. [Google Scholar] [CrossRef]
- D’Amelia, V.; Aversano, R.; Chiaiese, P.; Carputo, D. The antioxidant properties of plant flavonoids: Their exploitation by molecular plant breeding. Phytochem. Rev. 2018, 17, 611–625. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Fini, A.; Gori, A.; Guidi, L.; Landi, M.; Sebastiani, F.; Tattini, M. Are flavonoids effective antioxidants in plants? twenty years of our investigation. Antioxidants 2020, 9, 1098. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Adem, G.D.; Roy, S.J.; Zhou, M.; Bowman, J.P.; Shabala, S. Evaluating contribution of ionic, osmotic and oxidative stress components towards salinity tolerance in barley. BMC Plant Biol. 2014, 14, 113. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Ahn, M.J.; Hwang, G.S.; An, S.E.; Whang, W.K. Cosmeceutical bioactivities of isolated compounds from Ligularia fischeri Turcz leaves. Appl. Biol. Chem. 2016, 59, 485–494. [Google Scholar] [CrossRef]
- Barroso, M.R.; Barros, L.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Fernandes, I.P.; Barreiro, M.F.; Ferreira, I.C.F.R. Exploring the antioxidant potential of Helichrysum stoechas (L.) Moench phenolic compounds for cosmetic applications: Chemical characterization, microencapsulation and incorporation into a moisturizer. Ind. Crops Prod. 2014, 53, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Dorni, A.I.C.; Amalraj, A.; Gopi, S.; Varma, K.; Anjana, S.N. Novel cosmeceuticals from plants—An industry guided review. J. Appl. Res. Med. Aromat. Plants 2017, 7, 1–26. [Google Scholar]
- Taofiq, O.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C.F.R.; McPhee, D.J. Hydroxycinnamic acids and their derivatives: Cosmeceutical significance, challenges and future perspectives, a review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef]
- Lonati, E.; Carrozzini, T.; Bruni, I.; Mena, P.; Botto, L.; Cazzaniga, E.; Del Rio, D.; Labra, M.; Palestini, P.; Bulbarelli, A. Coffee-derived phenolic compounds activate NRF2 antioxidant pathway in I/R injury in vitro model: A nutritional approach preventing age related-damages. Molecules 2022, 27, 1049. [Google Scholar] [CrossRef] [PubMed]
- Kamarauskaite, J.; Baniene, R.; Raudone, L.; Vilkickyte, G.; Vainoriene, R.; Motiekaityte, V.; Trumbeckaite, S. Antioxidant and mitochondria-targeted activity of caffeoylquinic-acid-rich fractions of wormwood (Artemisia absinthium L.) and silver wormwood (Artemisia ludoviciana nutt.). Antioxidants 2021, 10, 1405. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, L.A.; Zubieta, C.; Lee, S.G.; Wang, Y.; Acajjaoui, S.; Timmins, J.; McSweeney, S.; Jez, J.M.; McCarthy, J.G.; McCarthy, A.A. A structural basis for the biosynthesis of the major chlorogenic acids found in coffee. Plant Physiol. 2012, 160, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef] [PubMed]
- Meneghini, M.; Pandino, G.; Lombardo, S.; Mauromicale, G.; Tavazza, R. Exploration to use of globe artichoke cell cultures as bioactive compounds production. Acta Hortic. 2016, 1147, 413–420. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Fevereiro, P.; Cabral, J.M.S.; Novais, J.M.; Salomé, M.; Pais, S. Callus and suspension cultures for biomass production of Cynara cardunculus (Compositae). Biotechnol. Lett. 1987, 9, 213–218. [Google Scholar] [CrossRef]
- Bimonte, M.; Carola, A.; Tito, A.; Barbulova, A.; Carucci, F.; Apone, F.; Monoli, I.; Cucchiara, M.; Hill, J.; Colucci, G. Coffea bengalensis for antiwrinkle and skin toning applications. Cosmet. Toilet. 2011, 126, 644–650. [Google Scholar]
- Barbulova, A.; Apone, F.; Colucci, G. Plant Cell Cultures as Source of Cosmetic Active Ingredients. Cosmetics 2014, 1, 94–104. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 25 °C | 6 °C | 25 °C | 6 °C | |
|---|---|---|---|---|
| Compounds | 7 Days | 14 Days | ||
| µg g−1 DW | µg g−1 DW | |||
| 3-CQA (CGA) | 1.741 ± 0.141 b | 34.299 ± 1.658 a | 25.805 ± 1.630 b | 33.113 ± 2.662 a |
| 3-FQA | 3.987 ± 0.155 b | 5.621 ± 0.549 a | 5.876 ± 0.628 a | 5.988 ± 0.226 a |
| 5-FQA | 77.541 ± 1.417 b | 112.655 ± 2.513 a | 86.781 ± 3.452 b | 93.098 ± 1.761 a |
| 3,4-DiCQA | 2543.67 ± 12.504 b | 3512.788 ± 11.437 a | 2865.411 ± 144.080 b | 3276.89 ± 68.011 a |
| 1,5-DiCQA | 2346.451 ± 53.713 b | 3562.412 ± 199.066 a | 2755.342 ± 110.307 b | 3145.453 ± 187.963 a |
| 5-iFQA | 17.766 ± 1.274 b | 31.711 ± 0.740 a | 21.432 ± 1.853 b | 29.882 ± 2.208 a |
| p-coumaric acid | 4.785 ± 0.186 b | 16.341 ± 1.989 a | 3.834 ± 0.270 b | 8.999 ± 0.456 a |
| quercetin-glucoside | 1.627 ± 0.278 b | 5.36 ± 0.053 a | 3.521 ± 0.282 a | 1.117 ± 0.289 b |
| myricetin | N.d | N.d | N.d | N.d |
| kaempferolo-glucoside | 1.226 ± 0.005 a | 1.211 ± 0.156 a | 1.226 ± 0.161 a | 1.222 ± 0.157 a |
| naringin | 1.431 ± 0.114 a | 1.185 ± 0.097 b | 0.989 ± 0.032 b | 1.347 ± 0.130 a |
| vitexin | N.d | N.d | 0.026 ± 0.008 | 0.014 ± 0.001 |
| quercetin | 0.277 ± 0.015 b | 0.416 ± 0.006 a | 0.137 ± 0.005 b | 0.277 ± 0.039 a |
| diosmin | N.d | N.d | N.d | N.d |
| luteolin | 1.746 ± 0.106 a | 1.768 ± 0.137 a | 1.741 ± 0.255 a | 1.74 ± 0.133 a |
| kaempferol | 3.666 ± 0.140 a | 3.709 ± 0.379 a | 3.641 ± 0.265 a | 3.65 ± 0.257 a |
| apigenin | 0.011 ± 0.002 b | 0.024 ± 0.001 a | 0.007 ± 0.001 a | 0.007 ± 0.000 a |
| Total phenols | 5005.980 ± 91.680 b | 7289.527 ± 211.430 a | 5775.769 ± 121.320 b | 6602.797 ± 129.770 a |
| 0 mM | 50 mM NaCl | 100 mM NaCl | 150 mM NaCl | 0 mM | 50 mM NaCl | 100 mM NaCl | 150 mM NaCl | |
|---|---|---|---|---|---|---|---|---|
| 14 Days | 28 Days | |||||||
| Compounds | µg g−1 DW | µg g−1 DW | ||||||
| 3-CQA (CGA) | 28.944 ± 1.448 d | 53.423 ± 1.531 c | 81.650 ± 3.256 b | 140.439 ± 2.647 a | 8.315 ± 0.140 d | 11.6015 ± 0.190 c | 165.577 ± 3.163 b | 263.123 ± 2.833 a |
| 3-FQA | 5.760 ± 0.274 b | 5.987 ± 0.249 b | 6.099 ± 0.109 ab | 6.544 ± 0.250 a | 5.154 ± 0.280 d | 5.888 ± 0.00 c | 6.032 ± 0.014 b | 6.987 ± 0.190 a |
| 5-FQA | 85.445 ± 1.276 d | 153.645 ± 4.728 b | 134.987 ± 2.325 c | 165.981 ± 3.746 a | 81.876 ± 0.629 c | 142.660 ± 0.00 b | 145.898 ± 3.643 b | 187.977 ± 0.786 a |
| 3,4-DiCQA | 2653.866 ± 7.959 d | 2867.987 ± 3.425 c | 4087.871 ± 142.984 b | 4545.566 ± 112.699 a | 2438.866 ± 18.321 c | 5127.761 ± 16.971 b | 5523.761 ± 412.001 b | 6287.980 ± 29.602 a |
| 1,5-DiCQA | 2433.451 ± 1.724 d | 2687.342 ± 46.400 c | 3942.544 ± 93.496 b | 4312.232 ± 12.736 a | 2232.341 ± 141.293 c | 5012.544 ± 165.614 b | 5171.671 ± 225.069 b | 6023.561 ± 120.024 a |
| 5-iFQA | 19.566 ± 0.789 c | 21.688 ± 2.056 bc | 24.642 ± 1.840 b | 27.828 ± 0.697 a | 14.761 ± 0.214 d | 19.877 ± 0.502 c | 24.882 ± 2.073 b | 31.761 ± 0.949 a |
| p-coumaric acid | 4.556 ± 0.332 c | 12.559 ± 3.121 b | 9.461 ± 1.372 b | 27.552 ± 1.585 a | 7.747 ± 0.135 d | 9.926 ± 0.561 c | 12.789 ± 0.767 b | 16.500 ± 1.667 a |
| quercetin-glucoside | 2.911 ± 0.537 d | 3.792 ± 0.054 c | 8.174 ± 0.052 b | 9.500 ± 0.253 a | 3.439 ± 0.272 c | 6.641 ± 0.140 b | 8.412 ± 0.129 a | 7.199 ± 1.055 a |
| myricetin | N.d | N.d | N.d | N.d | N.d | N.d | N.d | N.d |
| kaempferolo-glucoside | 1.216 ± 0.306 a | 1.308 ± 0.161 a | 1.219 ± 0.017 a | 1.205 ± 0.023 a | 1.254 ± 0.032 a | 1.222 ± 0.137 a | 1.213 ± 0.143 a | 1.206 ± 0.135 a |
| naringin | 1.655 ± 0.093 a | 1.310 ± 0.009 b | 1.176 ± 0.082 c | 0.806 ± 0.148 d | 0.868 ± 0.075 b | 0.767 ± 0.123 b | 1090 ± 0.215 ab | 1151 ± 0.048 a |
| vitexin | 0.024 ± 0.002 b | 0.030 ± 0.004 a | 0.013 ± 0.001 c | 0.004 ± 0.002 d | 0.022 ± 0.028 a | 0.003 ± 0.000 c | 0.010 ± 0.001 b | 0.005 ± 0.001 c |
| quercetin | 0.225 ± 0.135 ab | 0.272 ± 0.010 a | 0.075 ± 0.011 c | 0.149 ± 0.015 b | 0.240 ± 0.086 a | 0.216 ± 0.049 a | 0.151 ± 0.001 c | 0.182 ± 0.001 b |
| diosmin | N.d | N.d | N.d | N.d | N.d | N.d | N.d | N.d |
| luteolin | 1.737 ± 0.197 | 1.739 ± 0.038 | N.d | N.d | 1.736 ± 0.176 a | 1.735 ± 0.002 a | N.d | N.d |
| kaempferol | 3.626 ± 0.050 a | 3.613 ± 0.281 a | 3.610 ± 0.144 a | 3.604 ± 0.011 a | 3.619 ± 0.032 a | 3.618 ± 0.133 a | 3.616 ± 0.432 a | 3608 ± 0.037 a |
| apigenin | 0.004 ± 0.00 a | 0.004 ± 0.00 a | N.d | 0.001 ± 0.00 b | 0.003 ± 0.00 a | 0.002 ± 0.00 a | 0.002 ± 0.00 a | 0.001 ± 0.00 a |
| Total phenols | 5242.985 ± 120.210 d | 5814.700 ± 162.332 c | 8301.521 ± 234.433 b | 9241.412 ± 231.220 a | 4800.243 ± 132.440 d | 10,448.876 ± 123.331 c | 11,065.104 ± 354.540 b | 12,831.239 ± 121.433 a |
| Treatments | MDA Value μmol g−1 FW | ||
|---|---|---|---|
| 7 days | 14 days | ||
| Control | 25 °C | 4.404 ± 0.357 a | 4.679 ± 1.018 a |
| Chilling Stress | 6 °C | 3.544 ± 0.215 b | 3.785 ± 0.413 a |
| 14 days | 28 days | ||
| Control | 0 mM | 3.578 ± 0.705 b | 3.475 ± 0.677 b |
| Salt stress (NaCl) | 50 mM | 3.939 ± 0.775 ab | 5.012 ± 0.559 a |
| 100 mM | 4.955 ± 0.477 a | 5.058 ± 1.424 ab | |
| 150 mM | 5.5651 ± 0.921 a | 6.664 ± 1332 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Alessandro, R.; Docimo, T.; Graziani, G.; D’Amelia, V.; De Palma, M.; Cappetta, E.; Tucci, M. Abiotic Stresses Elicitation Potentiates the Productiveness of Cardoon Calli as Bio-Factories for Specialized Metabolites Production. Antioxidants 2022, 11, 1041. https://doi.org/10.3390/antiox11061041
D’Alessandro R, Docimo T, Graziani G, D’Amelia V, De Palma M, Cappetta E, Tucci M. Abiotic Stresses Elicitation Potentiates the Productiveness of Cardoon Calli as Bio-Factories for Specialized Metabolites Production. Antioxidants. 2022; 11(6):1041. https://doi.org/10.3390/antiox11061041
Chicago/Turabian StyleD’Alessandro, Rosa, Teresa Docimo, Giulia Graziani, Vincenzo D’Amelia, Monica De Palma, Elisa Cappetta, and Marina Tucci. 2022. "Abiotic Stresses Elicitation Potentiates the Productiveness of Cardoon Calli as Bio-Factories for Specialized Metabolites Production" Antioxidants 11, no. 6: 1041. https://doi.org/10.3390/antiox11061041
APA StyleD’Alessandro, R., Docimo, T., Graziani, G., D’Amelia, V., De Palma, M., Cappetta, E., & Tucci, M. (2022). Abiotic Stresses Elicitation Potentiates the Productiveness of Cardoon Calli as Bio-Factories for Specialized Metabolites Production. Antioxidants, 11(6), 1041. https://doi.org/10.3390/antiox11061041