
Variability of Anthocyanin Concentrations, Total Metabolite Contents and Antioxidant Activities in Adzuki Bean Cultivars

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Seed Materials, Cultivation, and Sample Preparation
2.3. Extraction and Quantification of Anthocyanins
2.4. Extract Preparation for Determination of Total Contents and Antioxidant Activities
2.4.1. Preparation of Defatted Extracts
2.4.2. Preparation of Undefatted Extracts
2.5. Determination of Total Saponin Content (TSC)
2.6. Determination of Total Phenolic Content (TPC)
2.7. Determination of Antioxidant Activities
2.7.1. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) Radical Scavenging Activity
2.7.2. Ferric Reducing Antioxidant Power (FRAP) Assay
2.7.3. Trolox Equivalent Antioxidant Capacity (TEAC)
2.8. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of Adzuki Bean Cultivars
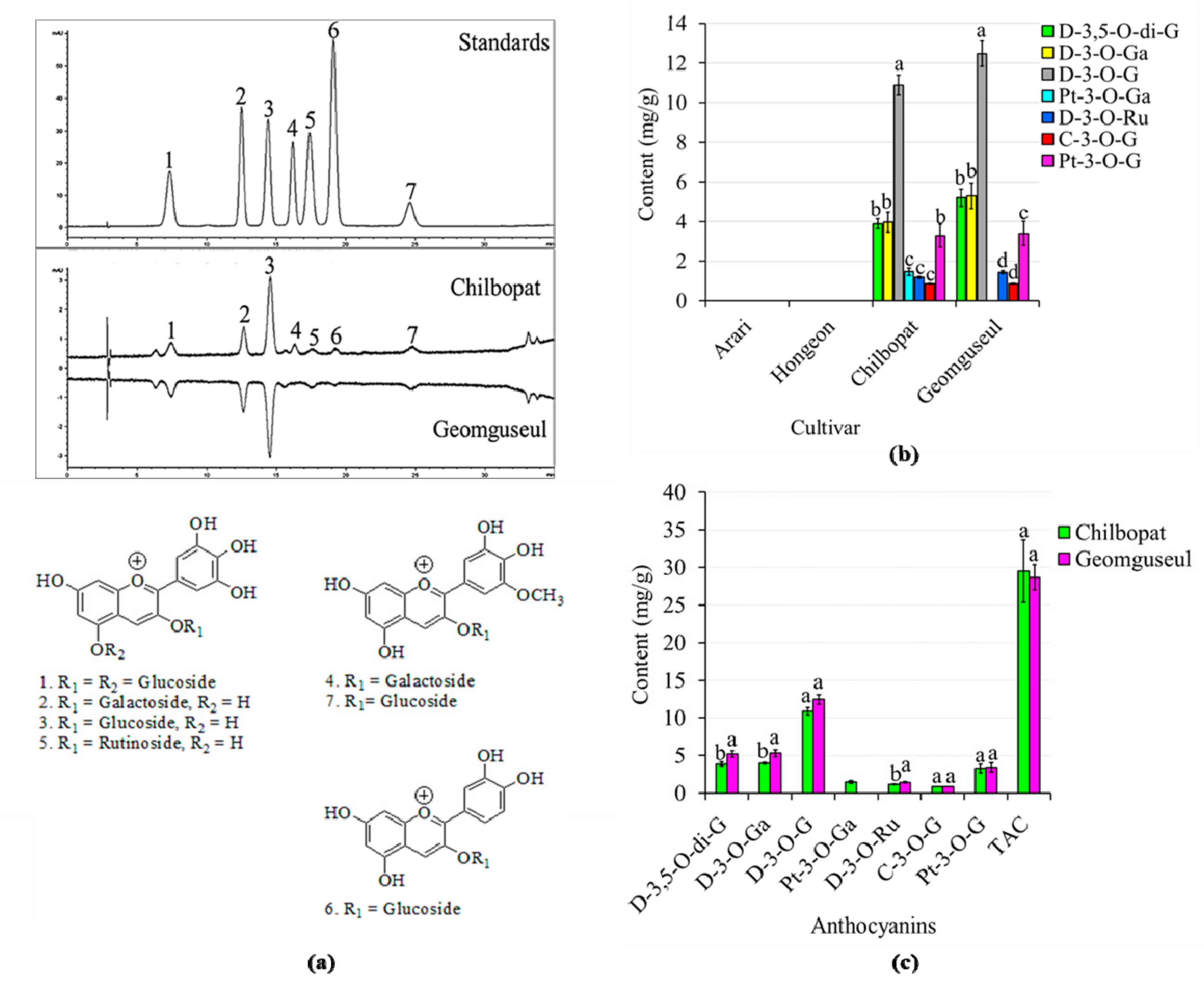
3.2. Variations of Anthocyanin Contents
3.3. TSC, TPC, and Antioxidant Activities
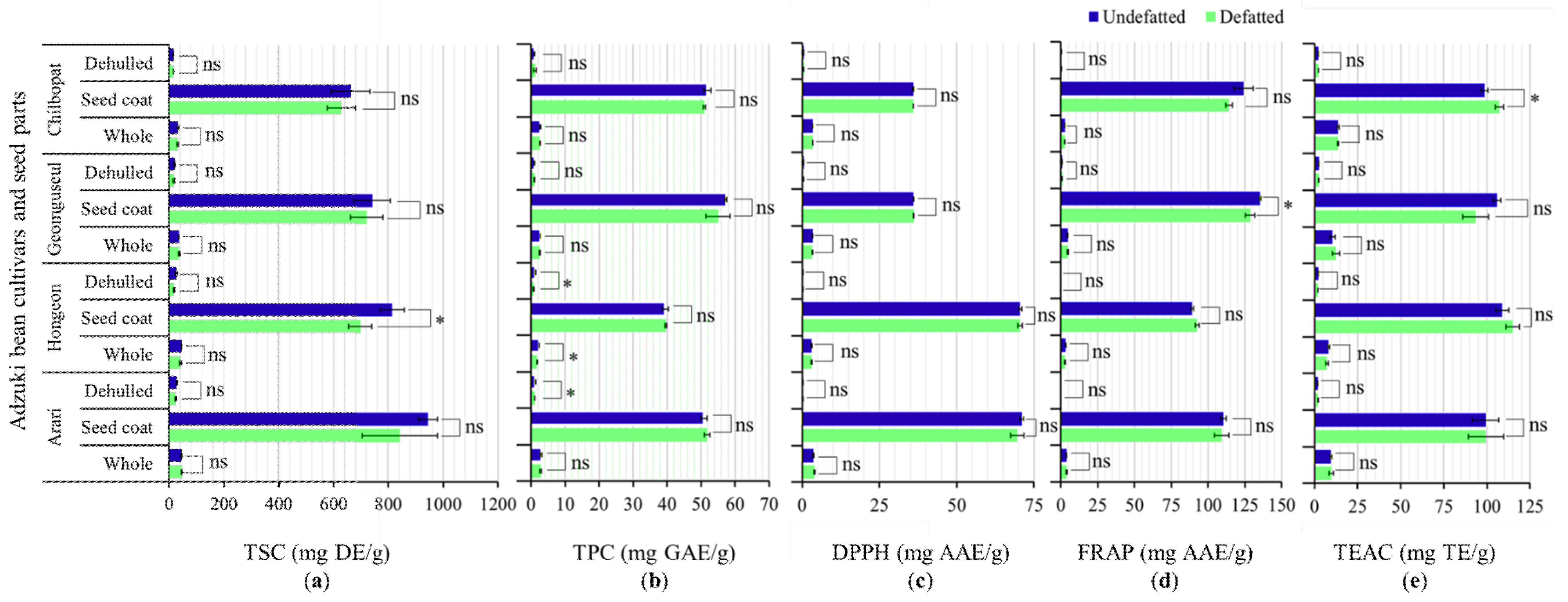
3.3.1. Effect of Defatting on TSC, TPC, and Antioxidant Activities
3.3.2. Variations of TSC, TPC, and Antioxidant Activities across Seed Parts
3.3.3. Variations of TSC, TPC, and Antioxidant Activities between Cultivars
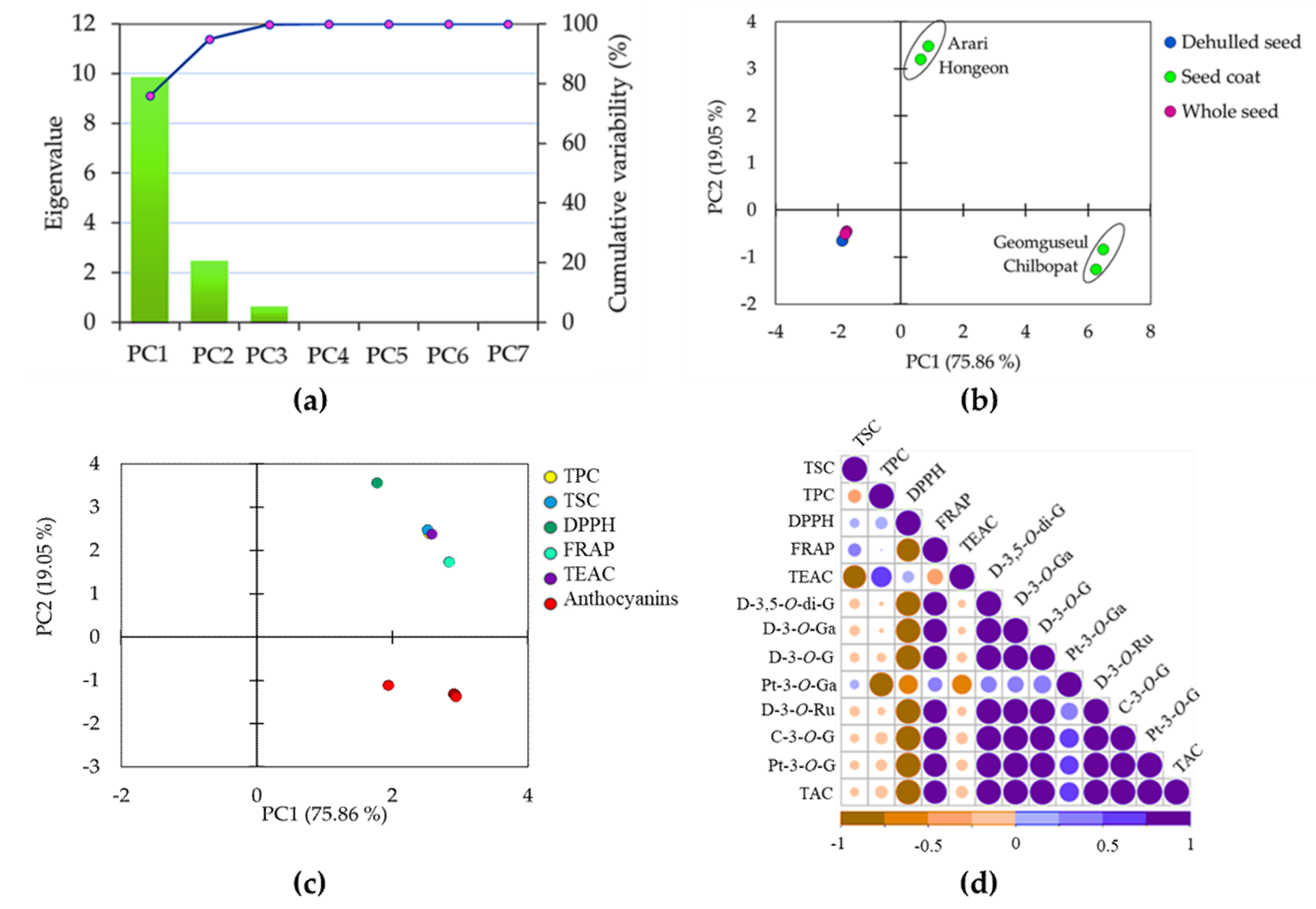
3.4. Principal Component and Correlation Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kawahara, S.-I.; Ishihara, C.; Matsumoto, K.; Senga, S.; Kawaguchi, K.; Yamamoto, A.; Suwannachot, J.; Hamauzu, Y.; Makabe, H.; Fujii, H. Identification and characterization of oligomeric proanthocyanidins with significant anti-cancer activity in adzuki beans (Vigna angularis). Heliyon 2019, 5, e02610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousif, A.; Kato, J.; Deeth, H. Effect of Storage on the Biochemical Structure and Processing Quality of Adzuki Bean (Vigna angularis). Food Rev. Int. 2007, 23, 1–33. [Google Scholar] [CrossRef]
- Horiuchi, Y.; Yamamoto, H.; Ogura, R.; Shimoda, N.; Sato, H.; Kato, K. Genetic analysis and molecular mapping of genes controlling seed coat colour in adzuki bean (Vigna angularis). Euphytica 2015, 206, 609–617. [Google Scholar] [CrossRef]
- Ha, T.J.; Park, J.E.; Lee, K.-S.; Seo, W.D.; Song, S.-B.; Lee, M.-H.; Kim, S.; Kim, J.-I.; Oh, E.; Pae, S.-B.; et al. Identification of anthocyanin compositions in black seed coated Korean adzuki bean (Vigna angularis) by NMR and UPLC-Q-Orbitrap-MS/MS and screening for their antioxidant properties using different solvent systems. Food Chem. 2020, 346, 128882. [Google Scholar] [CrossRef]
- FAO. The Global Economy of Pulses; Rawal, V., Navarro, D.K., Eds.; FAO: Rome, Italy, 2019. [Google Scholar] [CrossRef]
- Bi, S.; Wang, A.; Lao, F.; Shen, Q.; Liao, X.; Zhang, P.; Wu, J. Effects of frying, roasting and boiling on aroma profiles of adzuki beans (Vigna angularis) and potential of adzuki bean and millet flours to improve flavor and sensory characteristics of biscuits. Food Chem. 2020, 339, 127878. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, X.; Wang, P.; Jiang, H.; Li, W. Compositional, morphological, and physicochemical properties of starches from red adzuki bean, chickpea, faba bean, and baiyue bean grown in China. Food Sci. Nutr. 2019, 7, 2485–2494. [Google Scholar] [CrossRef] [Green Version]
- Reddy, C.K.; Luan, F.; Xu, B. Morphology, crystallinity, pasting, thermal and quality characteristics of starches from adzuki bean (Vigna angularis L.) and edible kudzu (Pueraria thomsonii Benth). Int. J. Biol. Macromol. 2017, 105, 354–362. [Google Scholar] [CrossRef]
- Yoshida, H.; Tomiyama, Y.; Yoshida, N.; Mizushina, Y. Characteristics of lipid components, fatty acid distributions and triacylglycerol molecular species of adzuki beans (Vigna angularis). Food Chem. 2009, 115, 1424–1429. [Google Scholar] [CrossRef]
- Takahama, U.; Yamauchi, R.; Hirota, S. Isolation and characterization of a cyanidin-catechin pigment from adzuki bean (Vigna angularis). Food Chem. 2013, 141, 282–288. [Google Scholar] [CrossRef]
- Han, K.H.; Kitano-Okada, T.; Seo, J.M.; Kim, S.J.; Sasaki, K.; Shimada, K.I.; Fukushima, M. Characterization of anthocyanins and proanthocyanidins of adzuki bean extracts and their antioxidant activity. J. Funct. Foods 2015, 14, 692–701. [Google Scholar] [CrossRef]
- Iida, T.; Yoshiki, Y.; Okubo, K.; Ohrui, H.; Kinjo, J.; Nohara, T. Triterpenoid saponins from Vigna angularis. Phytochemistry 1999, 51, 1055–1058. [Google Scholar] [CrossRef]
- Liu, R.; Cai, Z.; Xu, B. Characterization and quantification of flavonoids and saponins in adzuki bean (Vigna angularis L.) by HPLC–DAD–ESI–MSn analysis. Chem. Central J. 2017, 11, 93. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Zheng, Y.; Cai, Z.; Xu, B. Saponins and Flavonoids from Adzuki Bean (Vigna angularis L.) Ameliorate High-Fat Diet-Induced Obesity in ICR Mice. Front. Pharmacol. 2017, 8, 687. [Google Scholar] [CrossRef]
- Luo, J.; Cai, W.; Wu, T.; Xu, B. Phytochemical distribution in hull and cotyledon of adzuki bean (Vigna angularis L.) and mung bean (Vigna radiate L.), and their contribution to antioxidant, anti-inflammatory and anti-diabetic activities. Food Chem. 2016, 201, 350–360. [Google Scholar] [CrossRef]
- Woo, K.S.; Kim, H.-J.; Lee, J.H.; Lee, B.W.; Lee, Y.Y.; Lee, B.K.; Ko, J.Y. Quality Characteristics and Antioxidant Activities of Rice/Adzuki Bean Mixtures Cooked Using Two Different Methods. J. Food Qual. 2018, 2018, 4874795. [Google Scholar] [CrossRef]
- Takahama, U.; Yamauchi, R.; Hirota, S. Antioxidative flavonoids in adzuki-meshi (rice boiled with adzuki bean) react with nitrite under simulated stomach conditions. J. Funct. Foods 2016, 26, 657–666. [Google Scholar] [CrossRef]
- Kang, Y.J.; Satyawan, D.; Shim, S.; Lee, T.; Lee, J.; Hwang, W.J.; Kim, S.K.; Lestari, P.; Laosatit, K.; Kim, K.H.; et al. Draft genome sequence of adzuki bean, Vigna angularis. Sci. Rep. 2015, 5, srep08069. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Fan, B.; Cao, Z.; Su, Q.; Wang, Y.; Zhang, Z.; Tian, J. Development of a high-density genetic linkage map and identification of flowering time QTLs in adzuki bean (Vigna angularis). Sci. Rep. 2016, 6, 39523. [Google Scholar] [CrossRef] [Green Version]
- Banni, K.; Moe, K.T.; Park, Y.-J. Assessing genetic diversity, population structure and gene flow in the Korean red bean [Vigna angularis (Willd.) Ohwi & Ohashi] using SSR markers. Plant Genet. Resour. 2012, 10, 74–82. [Google Scholar] [CrossRef]
- Yao, Y.; Cheng, X.; Wang, S.; Wang, L.; Ren, G. Influence of altitudinal variation on the antioxidant and antidiabetic potential of azuki bean (Vigna angularis). Int. J. Food Sci. Nutr. 2011, 63, 117–124. [Google Scholar] [CrossRef]
- Kato, J.; Meguro, T.; Suzuki, M.M.; Deeth, H.C. Variations in the Seed Coat Colour of Adzuki Beans in the Aspects of Varieties, Harvest Years and Growing Locations, using Two-Dimensional Colour Mapping. Plant Prod. Sci. 2000, 3, 61–66. [Google Scholar] [CrossRef]
- Hori, Y.; Sato, S.; Hatai, A. Antibacterial activity of plant extracts from azuki beans (Vigna angularis) in vitro. Phytother. Res. 2006, 20, 162–164. [Google Scholar] [CrossRef]
- Kim, E.-H.; Song, H.-K.; Park, Y.-J.; Lee, J.-R.; Kim, M.-Y.; Chung, I.-M. Determination of Phenolic Compounds in Adzuki bean (Vigna angularis) Germplasm. Korean J. Crop Sci. 2011, 56, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Engquist, A.; Swanson, B.G. Microstructural differences among adzuki bean (Vigna-angularis) cultivars. Food Struct. 1992, 11, 171–179. [Google Scholar]
- Jia, R.; Ge, S.; Ren, S.; Luo, Y.; Xiu, L.; Liu, H.; Cai, D. Antibacterial mechanism of adzuki bean seed coat polyphenols and their potential application in preservation of fresh raw beef. Int. J. Food Sci. Technol. 2021, 56, 5025–5039. [Google Scholar] [CrossRef]
- Kawakami, W.; Oshima, A.; Yanase, E. Structural characterization of proanthocyanidins from adzuki seed coat. Food Chem. 2018, 239, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Yamate, J.; Hori, Y.; Hatai, A.; Nozawa, M.; Sagai, M. Protective effect of polyphenol-containing azuki bean (Vigna angularis) seed coats on the renal cortex in streptozotocin-induced diabetic rats. J. Nutr. Biochem. 2005, 16, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Siddique, K.H.M.; Johansen, C.; Turner, N.C.; Jeuffroy, M.-H.; Hashem, A.; Sakar, D.; Gan, Y.; Alghamdi, S.S. Innovations in agronomy for food legumes. A review. Agron. Sustain. Dev. 2011, 32, 45–64. [Google Scholar] [CrossRef] [Green Version]
- Chun, H.C.; Jung, K.Y.; Choi, Y.D.; Lee, S.; Song, S.; Ko, J.Y.; Choi, J.M. Differences in yields, antioxidant compounds, and antioxidant activity of ethanolic extracts among 11 adzuki bean cultivars (Vigna angularis L.) cultivated on a somewhat poorly drained paddy field. Korean J. Crop Sci. 2017, 62, 203–213. (In Korean) [Google Scholar]
- Jeong, E.-W.; Park, S.-Y.; Yang, Y.-S.; Baek, Y.-J.; Yun, D.-M.; Kim, H.-J.; Go, G.-W.; Lee, H.-G. Black Soybean and Adzuki Bean Extracts Lower Blood Pressure by Modulating the Renin-Angiotensin System in Spontaneously Hypertensive Rats. Foods 2021, 10, 1571. [Google Scholar] [CrossRef]
- Kim, T.J.; Park, J.G.; Kil Ahn, S.; Kim, K.W.; Choi, J.; Kim, H.Y.; Ha, S.-H.; Seo, W.D.; Kim, J.K. Discrimination of Adzuki Bean (Vigna angularis) Geographical Origin by Targeted and Non-Targeted Metabolite Profiling with Gas Chromatography Time-of-Flight Mass Spectrometry. Metabolites 2020, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.F.; Zhang, F.X.; Zhang, M.W.; Wei, Z.C.; Yang, C.Y.; Zhang, Y.; Tang, X.J.; Deng, Y.Y.; Chi, J.W. Phenolic Composition and Antioxidant Activity in Seed Coats of 60 Chinese Black Soybean (Glycine max L. Merr.) Varieties. J. Agric. Food Chem. 2011, 59, 5935–5944. [Google Scholar] [CrossRef]
- Wu, H.-J.; Deng, J.-C.; Yang, C.-Q.; Zhang, J.; Wang, X.-C.; Yang, F.; Yang, W.-Y.; Liu, J. Metabolite profiling of isoflavones and anthocyanins in black soybean [Glycine max (L.) Merr.] seeds by HPLC-MS and geographical differentiation analysis in Southwest China. Anal. Methods 2016, 9, 792–802. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K.C. Phytochemical Profiles and Health-Promoting Effects of Cool-Season Food Legumes as Influenced by Thermal Processing. J. Agric. Food Chem. 2009, 57, 10718–10731. [Google Scholar] [CrossRef]
- Roque, N.L.; Aguilar, S.; Quispe-Neyra, J.; Navarro, W.M.; Alfaro-Cruz, S.; Condezo-Hoyos, L. A green analytical assay for the quantitation of the total saponins in quinoa (Chenopodium quinoa Willd.) based on macro lens-coupled smartphone. Talanta 2019, 204, 576–585. [Google Scholar] [CrossRef]
- Choi, Y.M.; Yoon, H.; Shin, M.J.; Lee, Y.; Hur, O.S.; Lee, B.C.; Ha, B.K.; Wang, X.; Desta, K.T. Metabolite contents and antioxidant activities of soybean (Glycine max (L.) Merrill) seeds of different seed coat colors. Antioxidants 2021, 10, 1210. [Google Scholar] [CrossRef]
- Li, Y.; Yang, K.; Yang, W.; Chu, L.; Chen, C.; Zhao, B.; Li, Y.; Jian, J.; Yin, Z.; Wang, T.; et al. Identification of QTL and Qualitative Trait Loci for Agronomic Traits Using SNP Markers in the Adzuki Bean. Front. Plant Sci. 2017, 8, 840. [Google Scholar] [CrossRef] [Green Version]
- Choung, M.-G.; Choi, B.-R.; An, Y.-N.; Chu, Y.-H.; Cho, Y.-S. Anthocyanin Profile of Korean Cultivated Kidney Bean (Phaseolus vulgaris L.). J. Agric. Food Chem. 2003, 51, 7040–7043. [Google Scholar] [CrossRef]
- Harlen, W.C.; Jati, I.R.A. Antioxidant activity of anthocyanins in common legume grains. In Polyphenols: Mechanisms of Action in Human Health and Disease, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 81–92. [Google Scholar]
- Chu, L.; Zhao, P.; Wang, K.; Zhao, B.; Li, Y.; Yang, K.; Wan, P. VaSDC1 Is Involved in Modulation of Flavonoid Metabolic Pathways in Black and Red Seed Coats in Adzuki Bean (Vigna angularis L.). Front. Plant Sci. 2021, 12, 1464. [Google Scholar] [CrossRef]
- Díaz, A.M.; Caldas, G.V.; Blair, M.W. Concentrations of condensed tannins and anthocyanins in common bean seed coats. Food Res. Int. 2010, 43, 595–601. [Google Scholar] [CrossRef]
- Kan, L.; Nie, S.; Hu, J.; Liu, Z.; Xie, M. Antioxidant activities and anthocyanins composition of seed coats from twenty-six kidney bean cultivars. J. Funct. Foods 2016, 26, 622–631. [Google Scholar] [CrossRef]
- Kuang, Q.; Yu, Y.; Attree, R.; Xu, B. A comparative study on anthocyanin, saponin, and oil profiles of black and red seed coat peanut (Arachis hypogacea) grown in China. Int. J. Food Prop. 2017, 20, S131–S140. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Chu, L.; Wang, K.; Zhao, B.; Li, Y.; Yang, K.; Wan, P. Analyses on the pigment composition of different seed coat colors in adzuki bean. Food Sci. Nutr. 2022, 1–9. [Google Scholar] [CrossRef]
- Yoshida, K.; Sato, Y.; Okuno, R.; Kameda, K.; Isobe, M.; Kondo, T. Structural analysis and measurement of anthocyanins from colored seed coats of vigna, phaseolus, and glycine legumes. Biosci. Biotechnol. Biochem. 1996, 60, 589–593. [Google Scholar] [CrossRef]
- Zhang, A.; Jiang, X.; Ge, Y.; Xu, Q.; Li, Z.; Tang, H.; Cao, D.; Zhang, D. The Effects of GABA-Rich Adzuki Beans on Glycolipid Metabolism, as Well as Intestinal Flora, in Type 2 Diabetic Mice. Front. Nutr. 2022, 9, 849529. [Google Scholar] [CrossRef]
- Lee, J.H.; Jeon, J.K.; Kim, S.G.; Kim, S.H.; Chun, T.; Imm, J.Y. Comparative analyses of total phenols, flavonoids, saponins and antioxidant activity in yellow soy beans and mung beans. Int. J. Food Sci. Technol. 2011, 46, 2513–2519. [Google Scholar] [CrossRef]
- Sung, J.S.; Song, S.B.; Kim, J.Y.; An, Y.J.; Park, J.E.; Choe, M.E.; Chu, J.H.; Ha, T.J.; Han, S.I. Variation in physicochemical characteristics and antioxidant activities of small redbean cultivars. Korean J. Crop Sci. 2020, 65, 231–240. (In Korean) [Google Scholar]
- Bello, M.; Jiddah-Kazeem, B.; Fatoki, T.H.; Ibukun, E.O.; Akinmoladun, A.C. Antioxidant property of Eucalyptus globulus Labill. Extracts and inhibitory activities on carbohydrate metabolizing enzymes related to type-2 diabetes. Biocatal. Agric. Biotechnol. 2021, 36, 102111. [Google Scholar] [CrossRef]
- Yue, J.; Gu, Z.; Zhu, Z.; Yi, J.; Ohm, J.-B.; Chen, B.; Rao, J. Impact of defatting treatment and oat varieties on structural, functional properties, and aromatic profile of oat protein. Food Hydrocoll. 2020, 112, 106368. [Google Scholar] [CrossRef]
- Galili, S.; Hovav, R. Determination of polyphenols, flavonoids, and antioxidant capacity in dry seeds. In Polyphenols in Plants: Isolation, Purification and Extract Preparation, 1st ed.; Watson, R.R., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 305–323. [Google Scholar] [CrossRef]
- Huang, R.; Huang, K.; Guan, X.; Li, S.; Cao, H.; Zhang, Y.; Lao, X.; Bao, Y.; Wang, J. Effect of defatting and extruding treatment on the physicochemical and storage properties of quinoa (Chenopodium quinoa Wild) flour. LWT 2021, 147, 111612. [Google Scholar] [CrossRef]
- Mondor, M.; Aksay, S.; Drolet, H.; Roufik, S.; Farnworth, E.; Boye, J.I. Influence of processing on composition and antinutritional factors of chickpea protein concentrates produced by isoelectric precipitation and ultrafiltration. Innov. Food Sci. Emerg. Technol. 2009, 10, 342–347. [Google Scholar] [CrossRef]
- Buitimea-Cantúa, N.E.; Torres-Chávez, P.I.; Ledesma-Osuna, A.I.; Ramírez-Wong, B.; Robles-Sánchez, R.M.; Serna-Saldívar, S.O. Effect of defatting and decortication on distribution of fatty acids, phenolic and antioxidant compounds in sorghum (Sorghum bicolor) bran fractions. Int. J. Food Sci. Technol. 2013, 48, 2166–2175. [Google Scholar] [CrossRef]
- Nepote, V.; Grosso, N.R.; Guzmán, C.A. Optimization of extraction of phenolic antioxidants from peanut skins. J. Sci. Food Agric. 2004, 85, 33–38. [Google Scholar] [CrossRef]
- Kanatt, S.R.; Arjun, K.; Sharma, A. Antioxidant and antimicrobial activity of legume hulls. Food Res. Int. 2011, 44, 3182–3187. [Google Scholar] [CrossRef]
- Zilić, S.; Akillioǧlu, H.G.; Serpen, A.; Perić, V.; Gökmen, V. Comparisons of phenolic compounds, isoflavones, antioxidant capacity and oxidative enzymes in yellow and black soybeans seed coat and dehulled bean. Eur. Food Res. Technol. 2013, 237, 409–418. [Google Scholar] [CrossRef]
- Butkutė, B.; Lemežienė, N.; Dagilytė, A.; Cesevičienė, J.; Benetis, R.; Mikaliūnienė, J.; Rodovičius, H. Mineral element and total phenolic composition and antioxidant capacity of seeds and aerial plant parts of perennial legumes. Commun. Soil Sci. Plant Anal. 2016, 47, 36–45. [Google Scholar] [CrossRef]
- Dueñas, M.; Hernández, T.; Estrella, I. Assessment of in vitro antioxidant capacity of the seed coat and the cotyledon of legumes in relation to their phenolic contents. Food Chem. 2006, 98, 95–103. [Google Scholar] [CrossRef]
- Gohara, A.K.; De Souza, A.H.P.; Gomes, S.T.M.; De Souza, N.E.; Visentainer, J.V.; Matsushita, M. Nutritional and bioactive compounds of adzuki beans cultivars using chemometric approach. Cienc. Agrotec. 2016, 40, 104–113. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Gan, R.Y.; Ge, Y.Y.; Zhang, D.; Corke, H. Polyphenols in common beans (Phaseolus vulgaris L.): Chemistry, analysis, and factors affecting composition. Compr. Rev. Food Sci. F 2018, 17, 1518–1539. [Google Scholar] [CrossRef] [Green Version]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental abiotic factors on the content of saponins in plants. Phytochem. Rev. 2010, 10, 471–491. [Google Scholar] [CrossRef]
- Boudjou, S.; Oomah, B.D.; Zaidi, F.; Hosseinian, F. Phenolics content and antioxidant and anti-inflammatory activities of legume fractions. Food Chem. 2013, 138, 1543–1550. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Adzuki Bean Cultivar | |||
|---|---|---|---|---|
| Chilbopat | Geomguseul | Hongeon | Arari | |
| Qualitative | ||||
| Hypocotyledon color | Green | Green | Green | Green |
| Growth type | Determinate | Determinate | Determinate | Determinate |
| Flower color | Light-yellow | Yellow | Yellow | Yellow |
| Pod color | Gray | Gray | Gray | Gray |
| Quantitative | ||||
| DF (days) | 60 | 49 | 32 | 49 |
| DM (days) | 95 | 90 | 59 | 90 |
| PPP (number, n = 3) | 47.7 ± 16.4 ab | 61.0 ± 6.5 a | 31.3 ± 10.1 b | 24.0 ± 10.4 b |
| SPP (number, n = 10) | 6.1 ± 0.7 c | 6.9 ± 0.7 b | 6.6 ± 0.5 bc | 7.9 ± 0.7 a |
| HSW (g, n = 3) | 17.2 ± 0.1 a | 13.1 ± 0.0 d | 14.0 ± 0.1 c | 14.3 ± 0.1 b |
| Metabolite Content | Extract | Seed Parts | Adzuki Bean Cultivars | Range | CV (%) | |||
|---|---|---|---|---|---|---|---|---|
| Chilbopat | Geomguseul | Hongeon | Arari | |||||
| TSC (mg DE/g) | Defatted | Whole seed | 32.44 ± 2.50 b,z | 37.93 ± 2.03 b,y | 42.38 ± 3.61 b,xy | 46.85 ± 0.55 b,x | 32.44–46.85 | 13.38 |
| Seed coat | 629.20 ± 51.81 a,y | 721.12 ± 59.22 a,xy | 698.01 ± 41.90 a,xy | 842.63 ± 136.74 a,x | 629.20–842.63 | 10.66 | ||
| Dehulled seed | 16.79 ± 0.74 b,y | 19.00 ± 3.09 b,y | 19.74 ± 1.78 b,y | 25.11 ± 2.74 b,x | 16.79–25.11 | 15.16 | ||
| Undefatted | Whole seed | 33.86 ± 3.24 b,y | 36.92 ± 1.16 b,y | 45.44 ± 0.43 b,x | 45.12 ± 1.94 b,x | 33.86–45.44 | 12.56 | |
| Seed coat | 663.13 ± 70.47 a,z | 741.74 ± 66.66 a,yz | 814.20 ± 44.08 a,y | 944.78 ± 34.18 a,x | 663.13–944.78 | 13.10 | ||
| Dehulled seed | 16.20 ± 1.81 b,y | 20.27 ± 2.95 b,y | 26.50 ± 4.13 b,x | 29.12 ± 1.58 b,x | 16.20–29.19 | 22.09 | ||
| TPC (mg GAE/g) | Defatted | Whole seed | 2.58 ± 0.07 b,x | 2.51 ± 0.15 b,x | 1.82 ± 0.13 b,y | 2.82 ± 0.29 b,x | 1.82–2.82 | 15.34 |
| Seed coat | 51.01 ± 0.33 a,y | 55.08 ± 3.53 a,x | 39.70 ± 0.24 a,z | 51.82 ± 0.86 a,xy | 39.70–55.08 | 11.75 | ||
| Dehulled seed | 1.19 ± 0.40 c,x | 1.05 ± 0.06 c,x | 0.80 ± 0.15 c,x | 1.07 ± 0.02 c,x | 0.80–1.19 | 14.12 | ||
| Undefatted | Whole seed | 2.72 ± 0.07 b,xy | 2.66 ± 0.15 b,yz | 2.41 ± 0.13 b,z | 2.98 ± 0.29 b,x | 2.41–2.98 | 7.53 | |
| Seed coat | 51.85 ± 0.33 a,y | 57.35 ± 3.53 a,x | 39.48 ± 0.24 a,z | 50.85 ± 0.86 a,y | 39.48–57.35 | 13.02 | ||
| Dehulled seed | 1.01 ± 0.40 c,y | 0.99 ± 0.06 b,y | 1.26 ± 0.15 c,x | 1.31 ± 0.02 c,x | 0.99–1.31 | 12.49 | ||
| Activity | Extracts | Seed Parts | Adzuki Bean Cultivars | Range | CV (%) | |||
|---|---|---|---|---|---|---|---|---|
| Chilbopat | Geomguseul | Hongeon | Arari | |||||
| DPPH (mg AAE/g) | Defatted | Whole seed | 3.43 ± 0.02 b,x | 3.33 ± 0.07 b,xy | 3.08 ± 0.23 b,y | 3.98 ± 0.20 b,w | 3.08–3.98 | 9.46 |
| Seed coat | 35.93 ± 0.00 a,x | 36.09 ± 0.06 a,x | 70.50 ± 0.74 a,w | 69.61 ± 2.18 a,w | 35.93–70.50 | 32.10 | ||
| Dehulled seed | 0.48 ± 0.05 c,w | 0.45 ± 0.06 c,w | 0.23 ± 0.05 c,x | 0.26 ± 0.01 c,x | 0.23–0.48 | 31.26 | ||
| Undefatted | Whole seed | 3.41 ± 0.01 b,x | 3.37 ± 0.01 b,x | 3.05 ± 1.04 b,y | 3.67 ± 2.00 b,w | 3.05–3.67 | 6.48 | |
| Seed coat | 35.26 ± 0.26 a,x | 36.05 ± 0.06 a,x | 70.52 ± 4.07 a,w | 71.07 ± 5.05 a,w | 35.26–71.07 | 32.63 | ||
| Dehulled seed | 0.48 ± 0.01 c,w | 0.51 ± 0.04 c,w | 0.31 ± 0.00 c,x | 0.24 ± 0.03 c,y | 0.24–0.51 | 29.38 | ||
| FRAP (mg AAE/g) | Defatted | Whole seed | 2.77 ± 0.21 b,y | 4.88 ± 0.37 b,w | 2.86 ± 0.28 b,y | 3.97 ± 0.26 b,x | 2.77–4.88 | 22.36 |
| Seed coat | 114.28 ± 2.31 a,x | 128.63 ± 3.23 a,w | 92.55 ± 1.30 a,y | 109.23 ± 4.85 a,x | 92.55–128.63 | 11.60 | ||
| Dehulled seed | 0.46 ± 0.03 b,x | 1.06 ± 0.01 c,w | 0.22 ± 0.06 c,y | 0.25 ± 0.04 b,y | 0.22–1.06 | 67.88 | ||
| Undefatted | Whole seed | 2.77 ± 0.12 b,z | 4.55 ± 0.26 b,w | 3.32 ± 0.17 b,y | 3.90 ± 0.23 b,x | 2.77–4.55 | 17.36 | |
| Seed coat | 124.16 ± 6.61 a,x | 135.30 ± 0.57 a,w | 89.25 ± 0.81 a,z | 110.44 ± 2.07 a,y | 89.25–135.30 | 14.96 | ||
| Dehulled seed | 0.46 ± 0.03 b,x | 1.07 ± 0.02 c,w | 0.26 ± 0.01 c,y | 0.20 ± 0.05 c,z | 0.20–1.07 | 69.54 | ||
| TEAC (mg TE/g) | Defatted | Whole seed | 13.72 ± 0.28 b,w | 12.44 ± 2.15 b,w | 7.21 ± 0.86 b,y | 9.83 ± 1.30 b,x | 7.21–13.72 | 23.18 |
| Seed coat | 107.24 ± 2.52 a,wx | 93.46 ± 7.40 a,y | 114.91 ± 3.80 a,w | 99.33 ± 10.27 a,xy | 93.46–114.91 | 7.81 | ||
| Dehulled seed | 2.36 ± 0.05 c,w | 2.51 ± 0.12 c,w | 1.97 ± 0.09 c,x | 2.08 ± 0.14 b,x | 1.97–2.51 | 9.60 | ||
| Undefatted | Whole seed | 13.77 ± 0.56 b,w | 10.39 ± 2.52 b,x | 8.18 ± 0.44 b,z | 9.47 ± 1.53 b,xy | 8.18–13.77 | 19.78 | |
| Seed coat | 98.55 ± 2.07 a,x | 105.82 ± 7.35 a,wx | 108.81 ± 3.74 a,w | 99.29 ± 10.64 a,x | 98.55–108.81 | 4.21 | ||
| Dehulled seed | 2.33 ± 0.04 c,x | 2.62 ± 0.05 c,w | 2.15 ± 0.01 c,y | 1.98 ± 0.04 b,z | 1.98–2.62 | 10.28 | ||
| Source of Variation | Df | SS | F-Value |
|---|---|---|---|
| Total phenolic content | |||
| Cultivar (C) | 3 | 324.97 | 156.35 **** |
| Extract (E) | 1 | 1.44 | 2.08 |
| Seed part (S) | 2 | 36,612.03 | 26,421.48 **** |
| C × E | 3 | 2.11 | 1.02 |
| C × S | 6 | 579.20 | 139.33 **** |
| E × S | 2 | 0.41 | 0.30 |
| C × E × S | 6 | 7.35 | 1.77 |
| Total saponin content | |||
| Cultivar | 3 | 75,940.45 | 15.48 **** |
| Extract | 1 | 10,229.88 | 6.25 * |
| Seed part | 2 | 8,437,242.12 | 2579.05 **** |
| C × E | 3 | 3984.82 | 0.81 |
| C × S | 6 | 114,827.42 | 11.70 **** |
| E × S | 2 | 17,748.72 | 5.43 ** |
| C × E × S | 6 | 6425.22 | 0.66 |
| DPPH | |||
| Cultivar | 3 | 2350.67 | 3100.43 **** |
| Extract | 1 | 0.18 | 0.72 |
| Seed part | 2 | 42,244.48 | 83,578.14 **** |
| C × E | 3 | 0.47 | 0.63 |
| C × S | 6 | 4767.76 | 3144.24 **** |
| E × S | 2 | 0.58 | 1.15 |
| C × E × S | 6 | 2.13 | 1.40 |
| TEAC | |||
| Cultivar | 3 | 141.74 | 4.15 * |
| Extract | 1 | 1.70 | 0.15 |
| Seed part | 2 | 151,354.40 | 6641.79 **** |
| C × E | 3 | 102.54 | 3.00 |
| C × S | 6 | 595.03 | 8.70 **** |
| E × S | 2 | 1.31 | 0.06 |
| C × E × S | 6 | 300.85 | 4.40 *** |
| FRAP | |||
| Cultivar | 3 | 1974.74 | 174.77 **** |
| Extract | 1 | 26.16 | 6.95 * |
| Seed part | 2 | 196,956.24 | 26,146.63 **** |
| C × E | 3 | 47.07 | 4.17* |
| C × S | 6 | 3422.82 | 151.46 **** |
| E × S | 2 | 52.10 | 6.92 ** |
| C × E × S | 6 | 106.69 | 4.72 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desta, K.T.; Yoon, H.; Shin, M.-J.; Lee, S.; Wang, X.-H.; Choi, Y.-M.; Yi, J.-Y. Variability of Anthocyanin Concentrations, Total Metabolite Contents and Antioxidant Activities in Adzuki Bean Cultivars. Antioxidants 2022, 11, 1134. https://doi.org/10.3390/antiox11061134
Desta KT, Yoon H, Shin M-J, Lee S, Wang X-H, Choi Y-M, Yi J-Y. Variability of Anthocyanin Concentrations, Total Metabolite Contents and Antioxidant Activities in Adzuki Bean Cultivars. Antioxidants. 2022; 11(6):1134. https://doi.org/10.3390/antiox11061134
Chicago/Turabian StyleDesta, Kebede Taye, Hyemyeong Yoon, Myoung-Jae Shin, Sukyeung Lee, Xiao-Han Wang, Yu-Mi Choi, and Jung-Yoon Yi. 2022. "Variability of Anthocyanin Concentrations, Total Metabolite Contents and Antioxidant Activities in Adzuki Bean Cultivars" Antioxidants 11, no. 6: 1134. https://doi.org/10.3390/antiox11061134
APA StyleDesta, K. T., Yoon, H., Shin, M.-J., Lee, S., Wang, X.-H., Choi, Y.-M., & Yi, J.-Y. (2022). Variability of Anthocyanin Concentrations, Total Metabolite Contents and Antioxidant Activities in Adzuki Bean Cultivars. Antioxidants, 11(6), 1134. https://doi.org/10.3390/antiox11061134