
Resveratrol Downmodulates Neutrophil Extracellular Trap (NET) Generation by Neutrophils in Patients with Severe COVID-19

 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Casuistic
2.2. Flow Cytometry for Cellular Phenotyping
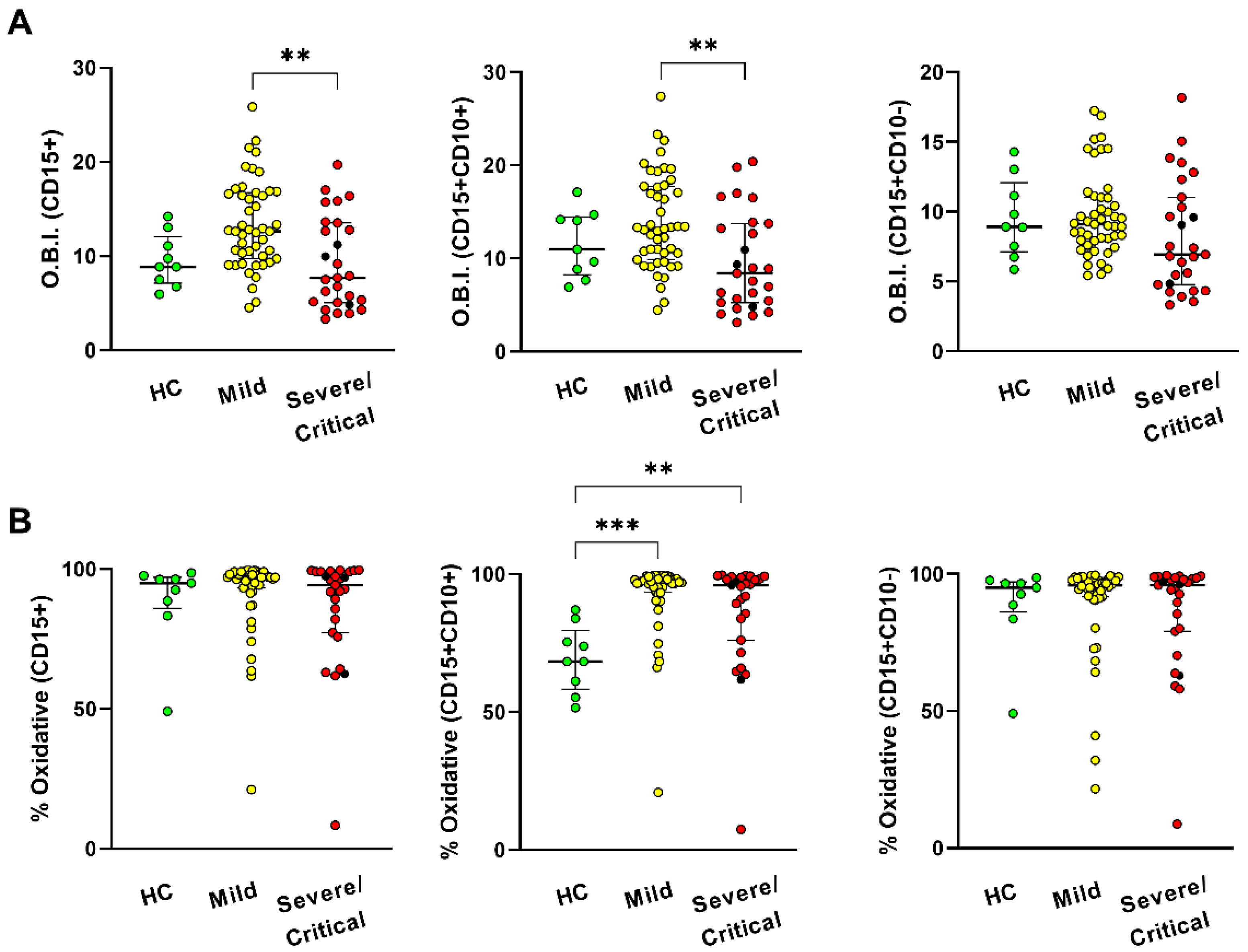
2.3. Measurement of Oxidative Burst
2.4. Neutrophil Enrichment and Culture with Resveratrol
2.5. NET Formation by Immunofluorescence
2.6. Measurements of Cytokines
2.7. NETs Quantification by Quant-iT PicoGreen
2.8. Statistical Analysis
3. Results
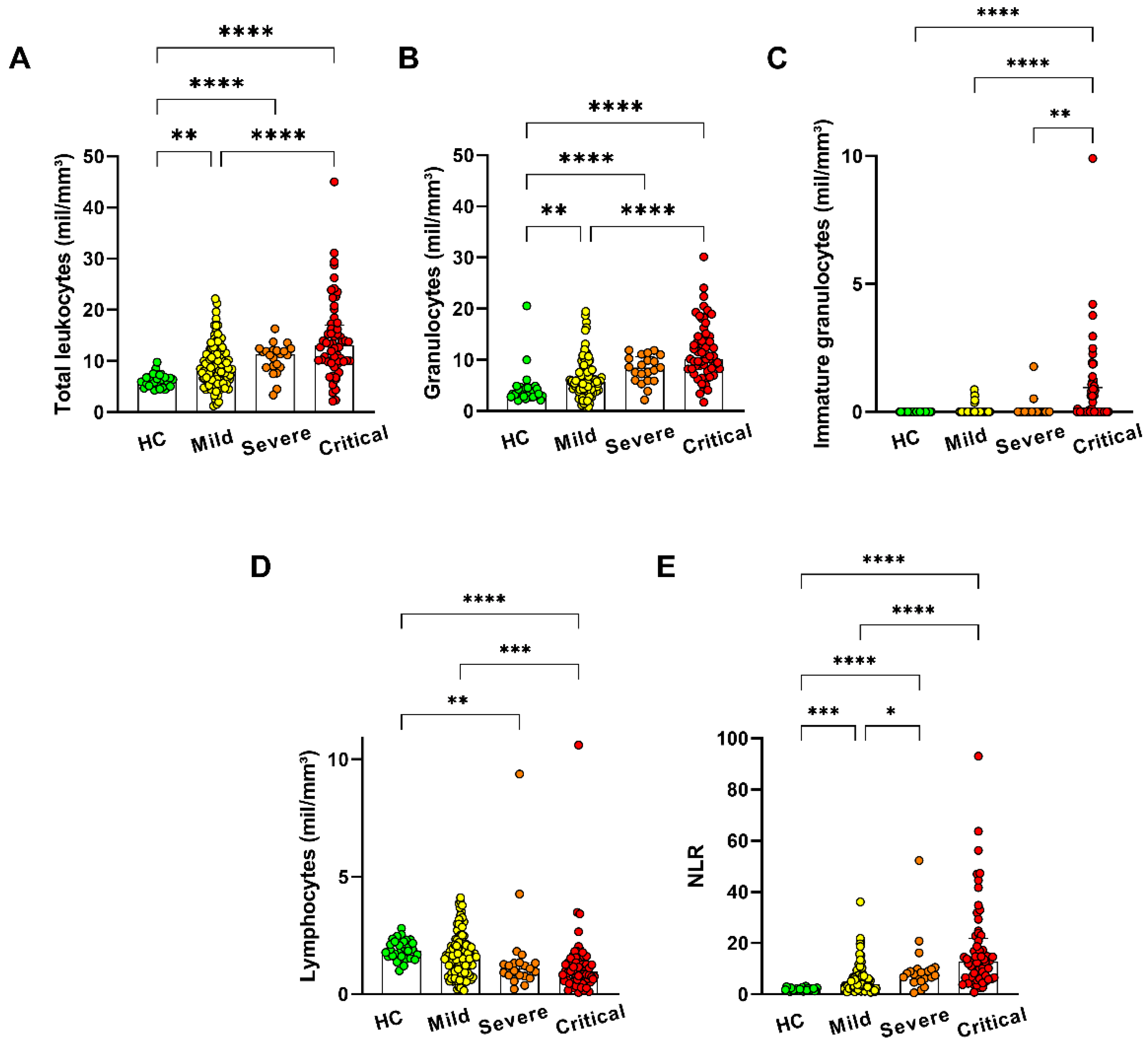
3.1. Characteristics of the COVID-19 Patients and Healthy Controls
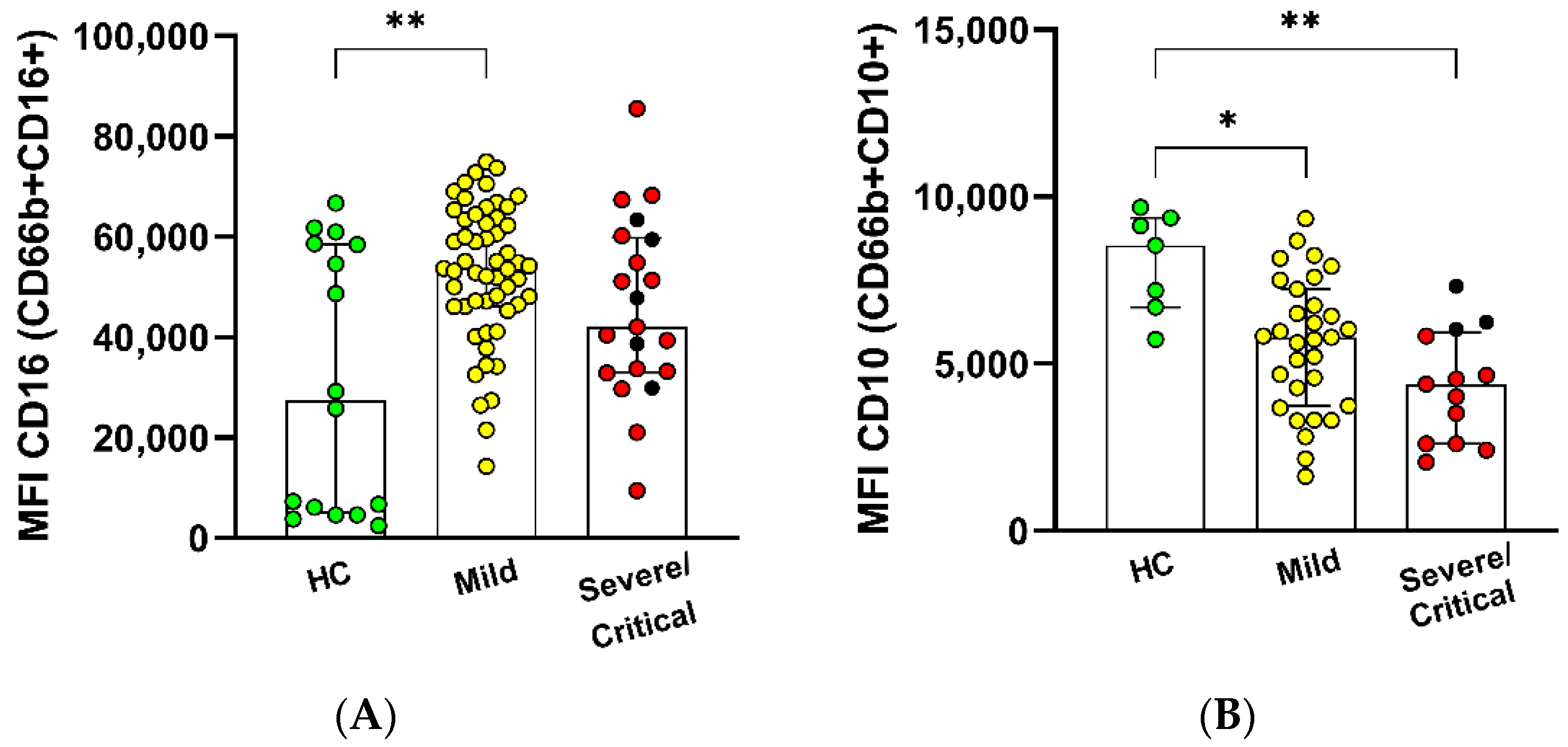
3.2. Cluster Analysis in Granulocyte Populations
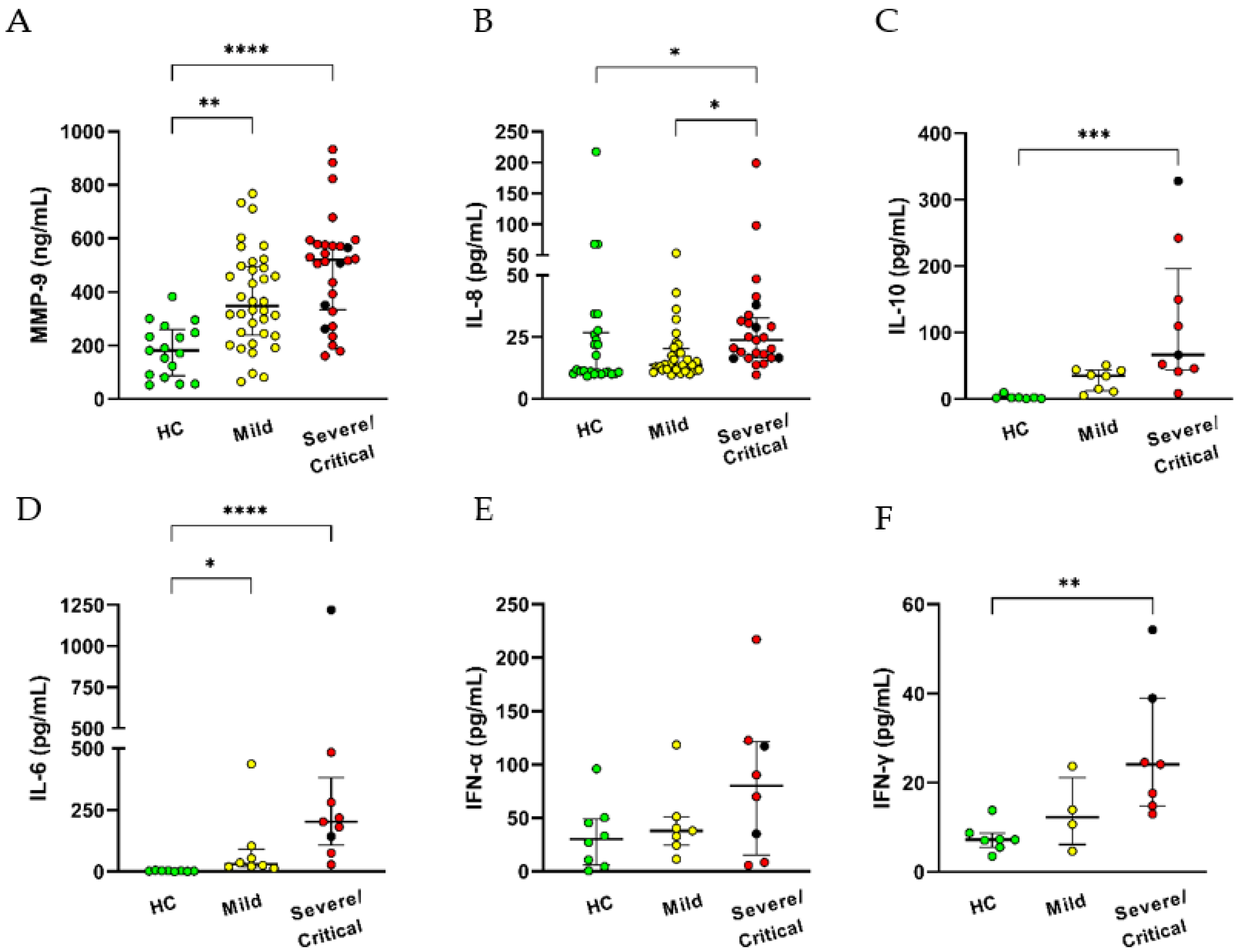
3.3. High Circulants Levels of Cytokines and Increased Neutrophil Oxidative Burst in COVID-19 Patients
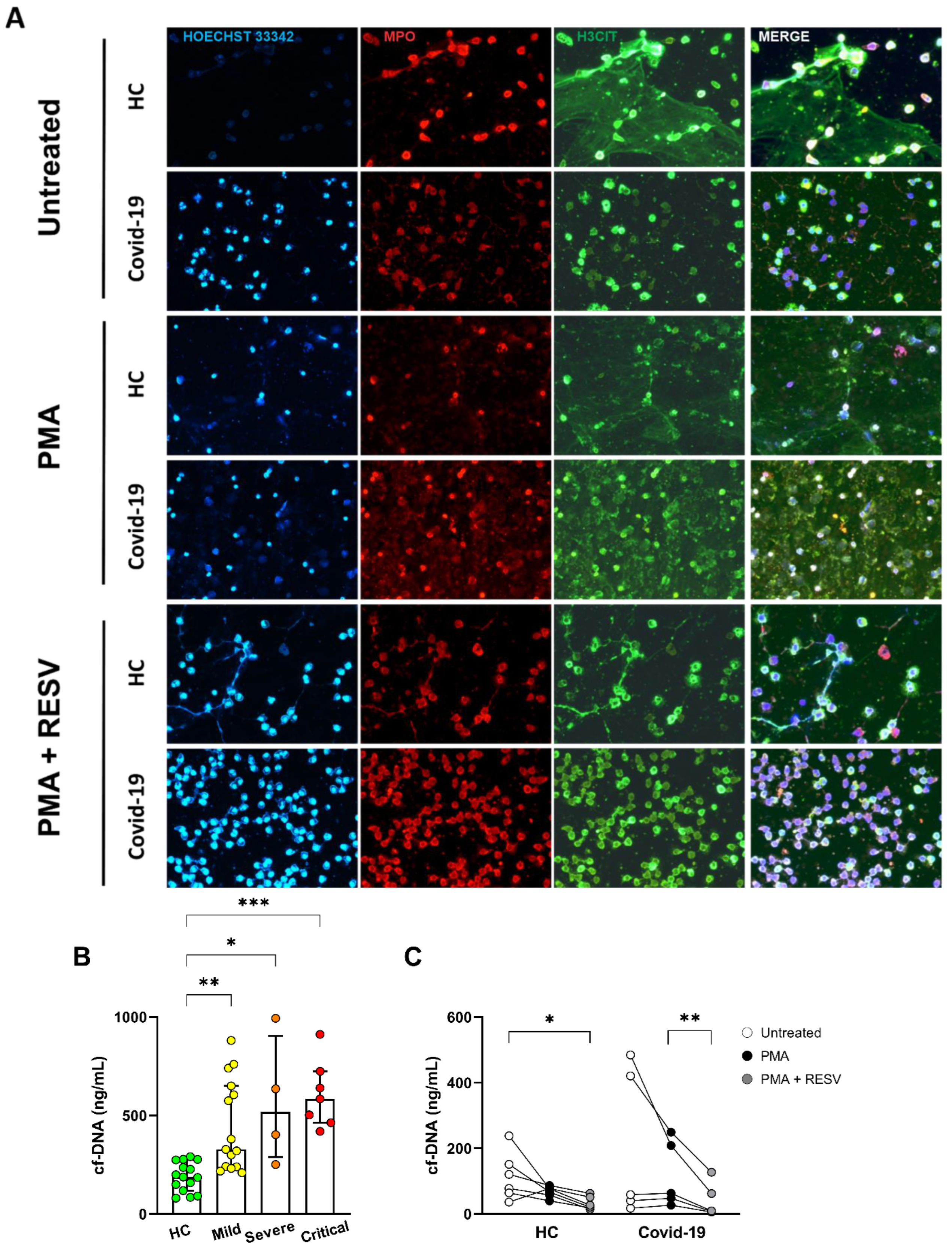
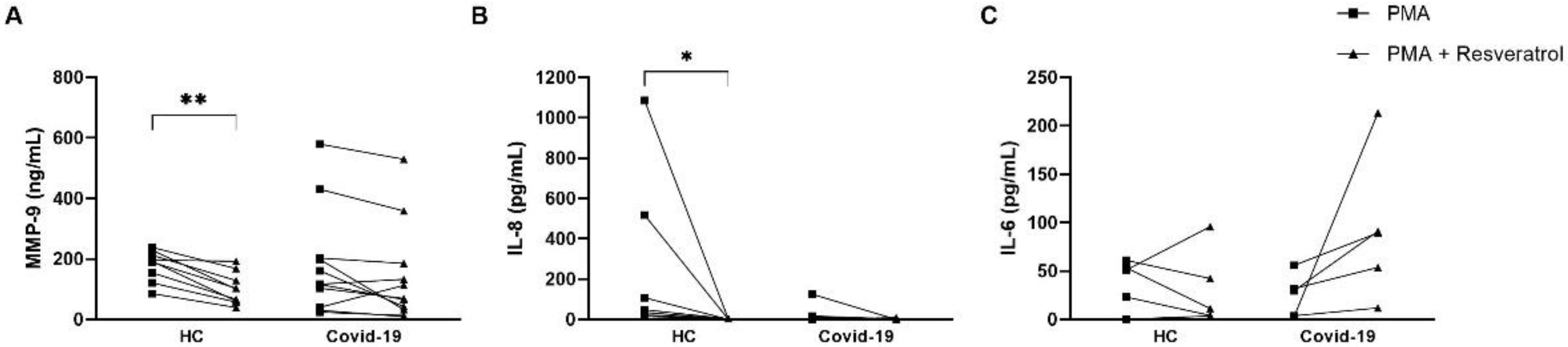
3.4. Resveratrol Downmodulates NET Formation in Neutrophils from COVID-19 Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020. Available online: https://pubmed.ncbi.nlm.nih.gov/32191675/ (accessed on 22 June 2022).
- WHO Coronavirus (COVID-19) Dashboard|WHO Coronavirus (COVID-19) Dashboard with Vaccination Data. Available online: https://COVID-1919.who.int/ (accessed on 22 June 2022).
- MacKman, N.; Antoniak, S.; Wolberg, A.S.; Kasthuri, R.; Key, N.S. Coagulation abnormalities and thrombosis in patients infected with SARS-CoV-2 and other pandemic viruses. Arter. Thromb. Vasc. Biol. 2020, 40, 2033–2044. Available online: https://www.ahajournals.org/doi/abs/10.1161/ATVBAHA.120.314514 (accessed on 22 June 2022). [CrossRef]
- Liu, Y.; Du, X.; Chen, J.; Jin, Y.; Peng, L.; Wang, H.H.; Luo, M.; Chen, L.; Zhao, Y. Neutrophil-to-lymphocyte ratio as an independent risk factor for mortality in hospitalized patients with COVID-19. J. Infect. 2020, 81, e6–e12. Available online: https://pubmed.ncbi.nlm.nih.gov/32283162/ (accessed on 22 June 2022). [CrossRef]
- Ng, H.; Havervall, S.; Rosell, A.; Aguilera, K.; Parv, K.; Von Meijenfeldt, F.A.; Lisman, T.; Mackman, N.; Thålin, C.; Phillipson, M. Circulating markers of neutrophil extracellular traps are of prognostic value in patients with COVID-19. Arter. Thromb. Vasc. Biol. 2021, 41, 988–994. Available online: https://pubmed.ncbi.nlm.nih.gov/33267662/ (accessed on 22 June 2022). [CrossRef]
- Cecchini, R.; Cecchini, A.L. SARS-CoV-2 infection pathogenesis is related to oxidative stress as a response to aggression. Med. Hypotheses 2020, 143, 110102. Available online: https://pubmed.ncbi.nlm.nih.gov/32721799/ (accessed on 22 June 2022). [CrossRef]
- Chernyak, B.V.; Popova, E.N.; Prikhodko, A.S.; Grebenchikov, O.A.; Zinovkina, L.A.; Zinovkin, R.A. COVID-19 and oxidative stress. Biochem. Biokhimiia. 2020, 85, 1543–1553. Available online: https://pubmed.ncbi.nlm.nih.gov/33705292/ (accessed on 22 June 2022). [CrossRef]
- Domi, E.; Hoxha, M.; Kolovani, E.; Tricarico, D.; Zappacosta, B. The importance of nutraceuticals in COVID-19: What’s the role of resveratrol? Molecules 2022, 27, 2376. Available online: https://pubmed.ncbi.nlm.nih.gov/35458574/ (accessed on 22 June 2022). [CrossRef]
- Jannin, B.; Menzel, M.; Berlot, J.P.; Delmas, D.; Lançon, A.; Latruffe, N. Transport of resveratrol, a cancer chemopreventive agent, to cellular targets: Plasmatic protein binding and cell uptake. Biochem. Pharmacol. 2004, 68, 1113–1118. [Google Scholar] [CrossRef]
- van Brummelen, R.; van Brummelen, A.C. The potential role of resveratrol as supportive antiviral in treating conditions such as COVID-19-A formulator’s perspective. Biomed. Pharmacother. 2022, 148, 112767. [Google Scholar] [CrossRef]
- Pasquereau, S.; Nehme, Z.; Haidar Ahmad, S.; Daouad, F.; Van Assche, J.; Wallet, C.; Schwartz, C.; Rohr, O.; Morot-Bizot, S.; Herbein, G. Resveratrol inhibits HCoV-229E and SARS-CoV-2 coronavirus replication in vitro. Viruses 2021, 13, 354. [Google Scholar] [CrossRef]
- Xiao, Z.; Ye, Q.; Duan, X.; Xiang, T. Network pharmacology reveals that resveratrol can alleviate COVID-19-related hyperinflammation. Dis. Markers 2021, 2021. [Google Scholar] [CrossRef]
- Palamara, A.T.; Nencioni, L.; Aquilano, K.; de Chiara, G.; Hernandez, L.; Cozzolino, F.; Ciriolo, M.R.; Garaci, E. Inhibition of influenza A virus replication by resveratrol. J. Infect. Dis. 2005, 191, 1719–1729. Available online: https://pubmed.ncbi.nlm.nih.gov/15838800/ (accessed on 22 June 2022). [CrossRef] [PubMed]
- Lee, S.; Yoon, K.D.; Lee, M.; Cho, Y.; Choi, G.; Jang, H.; Kim, B.S.; Jung, D.-H.; Oh, J.-G.; Kim, G.-W.; et al. Identification of a resveratrol tetramer as a potent inhibitor of hepatitis C virus helicase. Br. J. Pharmacol. 2016, 173, 191–211. Available online: https://pubmed.ncbi.nlm.nih.gov/26445091/ (accessed on 22 June 2022). [CrossRef] [PubMed]
- Cheltsov, A.V.; Aoyagi, M.; Aleshin, A.; Yu, E.C.W.; Gilliland, T.; Zhai, D.; Bobkov, A.A.; Reed, J.C.; Liddington, R.C.; Abagyan, R. Vaccinia virus virulence factor N1L is a novel promising target for antiviral therapeutic intervention. J. Med. Chem. 2010, 53, 3899–3906. Available online: https://pmc/articles/PMC2874095/ (accessed on 22 June 2022). [CrossRef]
- Clouser, C.L.; Chauhan, J.; Bess, M.A.; van Oploo, J.L.; Zhou, D.; Dimick-Gray, S.; Mansky, L.M.; Patterson, S.E. Anti-HIV-1 activity of resveratrol derivatives and synergistic inhibition of HIV-1 by the combination of resveratrol and decitabine. Bioorg. Med. Chem. Lett. 2012, 22, 6642–6646. Available online: https://pubmed.ncbi.nlm.nih.gov/23010273/ (accessed on 22 June 2022). [CrossRef]
- ter Ellen, B.M.; Dinesh Kumar, N.; Bouma, E.M.; Troost, B.; van de Pol, D.P.I.; van der Ende-Metselaar, H.H.; Apperloo, L.; van Gosliga, D.; van den Berge, M.; Nawijn, M.C.; et al. Resveratrol and pterostilbene inhibit SARS-CoV-2 replication in air–liquid interface cultured human primary bronchial epithelial cells. Viruses 2021, 13, 1335. Available online: https://www.mdpi.com/1999-4915/13/7/1335/htm (accessed on 9 August 2022). [CrossRef] [PubMed]
- Rice, C.M.; Lewis, P.; Ponce-Garcia, F.M.; Gibbs, W.; Cela, D.; Hamilton, F.; Arnold, D.; Hyams, C.; Oliver, E.; Barr, R.; et al. Neutrophils in severe COVID-19 are characterized by a hyperactive immature state and maintained CXCR2 expression. medRxiv 2022. Available online: https://www.medrxiv.org/content/10.1101/2022.03.23.22272828v1 (accessed on 22 June 2022). [CrossRef]
- Ssemaganda, A.; Kindinger, L.; Bergin, P.; Nielsen, L.; Mpendo, J.; Ssetaala, A.; Kiwanuka, N.; Munder, M.; Teoh, T.G.; Kropf, P.; et al. Characterization of neutrophil subsets in healthy human pregnancies. PLoS ONE 2014, 9, e85696. [Google Scholar] [CrossRef]
- Morrissey, S.M.; Geller, A.E.; Hu, X.; Tieri, D.; Ding, C.; Klaes, C.K.; Cooke, E.A.; Woeste, M.R.; Martin, Z.C.; Chen, O.; et al. A specific low-density neutrophil population correlates with hypercoagulation and disease severity in hospitalized COVID-19 patients. JCI Insight 2021, 6. Available online: https://pubmed.ncbi.nlm.nih.gov/33986193/ (accessed on 22 June 2022). [CrossRef]
- Silswal, N.; Reddy, N.S.; Qureshi, A.A.; Qureshi, N. Resveratrol Downregulates biomarkers of sepsis via inhibition of proteasome’s proteases. Shock 2018, 50, 579. [Google Scholar] [CrossRef]
- Tsai, Y.F.; Chen, C.Y.; Chang, W.Y.; Syu, Y.T.; Hwang, T.L. Resveratrol suppresses neutrophil activation via inhibition of Src family kinases to attenuate lung injury. Free. Radic. Biol. Med. 2019, 145, 67–77. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0891584919312961 (accessed on 12 November 2020). [CrossRef]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A double-edged sword in health benefits. Biomedicines 2018, 6, 91. Available online: https://pubmed.ncbi.nlm.nih.gov/30205595/ (accessed on 26 June 2022). [CrossRef]
- Roth, S.; Agthe, M.; Eickhoff, S.; Möller, S.; Karsten, C.M.; Borregaard, N.; Solbach, W.; Laskay, T. Secondary necrotic neutrophils release interleukin-16C and macrophage migration inhibitory factor from stores in the cytosol. Cell Death Discov. 2015, 1, 15056. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.E.; Souto, A.A.; Pitrez, P.M.C.; Stein, R.T.; Porto, B.N. Modulatory potential of resveratrol during lung inflammatory disease. Med. Hypotheses 2016, 96, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Mozzini, C.; Girelli, D. The role of Neutrophil Extracellular Traps in COVID-19: Only an hypothesis or a potential new field of research? Thromb. Res. 2020, 191, 26–27. Available online: http://www.thrombosisresearch.com/article/S004938482030147X/fulltext (accessed on 22 June 2022). [CrossRef] [PubMed]
- D′avila-Mesquita, C.; Couto, A.E.; Campos, L.C.; Vasconcelos, T.F.; Michelon-Barbosa, J.; Corsi, C.A.; Mestriner, F.; Petroski-Moraes, B.C.; Garbellini-Diab, M.J.; Couto, D.M.; et al. MMP-2 and MMP-9 levels in plasma are altered and associated with mortality in COVID-19 patients. Biomed. Pharmacother. 2021, 142, 112067. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, R.; Leunig, A.; Pekayvaz, K.; Popp, O.; Joppich, M.; Polewka, V.; Escaig, R.; Anjum, A.; Hoffknecht, M.-L.; Gold, C.; et al. Self-sustaining IL-8 loops drive a prothrombotic neutrophil phenotype in severe COVID-19. JCI Insight 2021, 6, e150862. Available online: https://pubmed.ncbi.nlm.nih.gov/34403366/ (accessed on 22 June 2022). [CrossRef] [PubMed]
- Zhu, Y.; Chen, X.; Liu, X. NETosis and neutrophil extracellular traps in COVID-19: Immunothrombosis and beyond. Front. Immunol. 2022, 13, 739. [Google Scholar] [CrossRef] [PubMed]
- Andargie, T.E.; Tsuji, N.; Seifuddin, F.; Jang, M.K.; Yuen, P.S.; Kong, H.; Tunc, I.; Singh, K.; Charya, A.; Wilkins, K.J.; et al. Cell-free DNA maps COVID-19 tissue injury and risk of death and can cause tissue injury. JCI Insight. 2021, 6, e147610. Available online: https://pubmed.ncbi.nlm.nih.gov/33651717/ (accessed on 22 June 2022). [CrossRef]
- Bradley, L.M.; Douglass, M.F.; Chatterjee, D.; Akira, S.; Baaten, B.J.G. Matrix metalloprotease 9 mediates neutrophil migration into the airways in response to influenza virus-induced toll-like receptor signaling. PLoS Pathog. 2012, 8, e1002641. Available online: https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1002641 (accessed on 22 June 2022). [CrossRef]
- Dhar, S.K.; Damodar, S.; Gujar, S.; Das, M. IL-6 and IL-10 as predictors of disease severity in COVID-19 patients: Results from meta-analysis and regression. Heliyon 2021, 7, e06155. Available online: https://doi.org/10.1016/j.heliyon.2021.e06155 (accessed on 22 June 2022). [CrossRef]
- Kasten, K.R.; Muenzer, J.T.; Caldwell, C.C. Neutrophils are significant producers of IL-10 during sepsis. Biochem. Biophys. Res. Commun. 2010, 393, 28–31. Available online: https://pubmed.ncbi.nlm.nih.gov/20097159/ (accessed on 26 June 2022). [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uninfected | Mild | Severe | Critical | |||||
|---|---|---|---|---|---|---|---|---|
| n = 46 | n = 108 | n = 21 | n = 61 | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Gender Male/Female | 16/30 | 65/43 | 14/7 | 37/24 | ||||
| Age-Male | 47 | 12.1 | 54 | 13.2 | 51 | 12.5 | 56 | 10.6 |
| Age-Female | 39 | 11.5 | 54 | 14.7 | 56 | 11.8 | 54 | 16.2 |
| Evolution | n | % | n | % | n | % | ||
| Discharge | 92 | 85.2 | 18 | 85.7 | 29 | 47.5 | ||
| Death | 6 | 5.6 | 2 | 9.5 | 28 | 45.9 | ||
| Transfer to another institute | 10 | 9.3 | 1 | 4.8 | 4 | 6.6 | ||
| Comorbidities | n | % | n | % | n | % | ||
| HAS | 56 | 51.9 | 11 | 52.4 | 33 | 54.1 | ||
| DM | 45 | 41.7 | 8 | 38.1 | 19 | 31.1 | ||
| Neoplasms | 22 | 20.4 | 2 | 9.5 | 4 | 6.6 | ||
| Vascular diseases | 10 | 9.3 | 1 | 4.8 | 7 | 11.5 | ||
| Obesity | 24 | 22.2 | 6 | 28.6 | 18 | 29.5 | ||
| Use of alcohol and cigarettes | 32 | 29.6 | 5 | 23.8 | 11 | 18.0 | ||
| Transplants | 10 | 9.3 | 0 | 0.0 | 4 | 6.6 | ||
| Kidney diseases | 15 | 13.9 | 0 | 0.0 | 4 | 6.6 | ||
| Gastrointestinal/Liver diseases | 7 | 6.5 | 1 | 4.8 | 4 | 6.6 | ||
| Heart diseases | 28 | 25.9 | 4 | 19.0 | 8 | 13.1 | ||
| Neurological diseases | 9 | 8.3 | 0 | 0.0 | 4 | 6.6 | ||
| Respiratory diseases | 15 | 13.9 | 0 | 0.0 | 6 | 9.8 | ||
| Metabolic/autoimmune diseases | 34 | 31.5 | 5 | 23.8 | 15 | 24.6 | ||
| Other infectious diseases | 9 | 8.3 | 0 | 0.0 | 2 | 3.3 | ||
| Other diseases | 10 | 9.3 | 3 | 14.3 | 9 | 14.8 | ||
| No comorbidities | 5 | 4.6 | 4 | 19.0 | 7 | 11.5 | ||
| 1 | 16 | 14.8 | 4 | 19.0 | 11 | 18.0 | ||
| 2 | 21 | 19.4 | 4 | 19.0 | 20 | 32.8 | ||
| 3 | 25 | 23.1 | 4 | 19.0 | 9 | 14.8 | ||
| ≥4 | 41 | 38.0 | 5 | 23.8 | 14 | 23.0 | ||
| Hospitalization history | Mean | SD | Mean | SD | Mean | SD | ||
| Days of symptoms until collection | 23.4 | 20.7 | 14.7 | 5.0 | 18.7 | 9.4 | ||
| Positive COVID-19 test days until collection | 15.8 | 19.7 | 7.5 | 4.4 | 10.2 | 8.8 | ||
| Hospitalization days until collection | 16.3 | 20.0 | 5.3 | 2.8 | 9.5 | 8.8 | ||
| Days from collection to completion | 11.3 | 16.9 | 15.0 | 10.0 | 27.4 | 25.4 | ||
| Total days of hospitalization | 27.6 | 27.8 | 20.3 | 10.8 | 36.9 | 26.5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza Andrade, M.M.; Leal, V.N.C.; Fernandes, I.G.; Gozzi-Silva, S.C.; Beserra, D.R.; Oliveira, E.A.; Teixeira, F.M.E.; Yendo, T.M.; Sousa, M.d.G.T.; Teodoro, W.R.; et al. Resveratrol Downmodulates Neutrophil Extracellular Trap (NET) Generation by Neutrophils in Patients with Severe COVID-19. Antioxidants 2022, 11, 1690. https://doi.org/10.3390/antiox11091690
de Souza Andrade MM, Leal VNC, Fernandes IG, Gozzi-Silva SC, Beserra DR, Oliveira EA, Teixeira FME, Yendo TM, Sousa MdGT, Teodoro WR, et al. Resveratrol Downmodulates Neutrophil Extracellular Trap (NET) Generation by Neutrophils in Patients with Severe COVID-19. Antioxidants. 2022; 11(9):1690. https://doi.org/10.3390/antiox11091690
Chicago/Turabian Stylede Souza Andrade, Milena M., Vinicius N. C. Leal, Iara G. Fernandes, Sarah C. Gozzi-Silva, Danielle R. Beserra, Emily A. Oliveira, Franciane M. E. Teixeira, Tatiana M. Yendo, Maria da Glória T. Sousa, Walcy R. Teodoro, and et al. 2022. "Resveratrol Downmodulates Neutrophil Extracellular Trap (NET) Generation by Neutrophils in Patients with Severe COVID-19" Antioxidants 11, no. 9: 1690. https://doi.org/10.3390/antiox11091690
APA Stylede Souza Andrade, M. M., Leal, V. N. C., Fernandes, I. G., Gozzi-Silva, S. C., Beserra, D. R., Oliveira, E. A., Teixeira, F. M. E., Yendo, T. M., Sousa, M. d. G. T., Teodoro, W. R., Oliveira, L. d. M., Alberca, R. W., Aoki, V., Duarte, A. J. S., & Sato, M. N. (2022). Resveratrol Downmodulates Neutrophil Extracellular Trap (NET) Generation by Neutrophils in Patients with Severe COVID-19. Antioxidants, 11(9), 1690. https://doi.org/10.3390/antiox11091690