
Exogenous Betaine Enhances the Protrusion Vigor of Rice Seeds under Heat Stress by Regulating Plant Hormone Signal Transduction and Its Interaction Network

Abstract
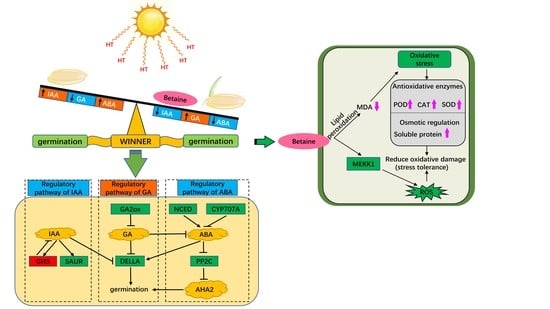
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Share and Cite
Mo, X.; Qian, J.; Liu, P.; Zeng, H.; Chen, G.; Wang, Y. Exogenous Betaine Enhances the Protrusion Vigor of Rice Seeds under Heat Stress by Regulating Plant Hormone Signal Transduction and Its Interaction Network. Antioxidants 2022, 11, 1792. https://doi.org/10.3390/antiox11091792
Mo X, Qian J, Liu P, Zeng H, Chen G, Wang Y. Exogenous Betaine Enhances the Protrusion Vigor of Rice Seeds under Heat Stress by Regulating Plant Hormone Signal Transduction and Its Interaction Network. Antioxidants. 2022; 11(9):1792. https://doi.org/10.3390/antiox11091792
Chicago/Turabian StyleMo, Xu, Jingya Qian, Peng Liu, Hongli Zeng, Guanghui Chen, and Yue Wang. 2022. "Exogenous Betaine Enhances the Protrusion Vigor of Rice Seeds under Heat Stress by Regulating Plant Hormone Signal Transduction and Its Interaction Network" Antioxidants 11, no. 9: 1792. https://doi.org/10.3390/antiox11091792
APA StyleMo, X., Qian, J., Liu, P., Zeng, H., Chen, G., & Wang, Y. (2022). Exogenous Betaine Enhances the Protrusion Vigor of Rice Seeds under Heat Stress by Regulating Plant Hormone Signal Transduction and Its Interaction Network. Antioxidants, 11(9), 1792. https://doi.org/10.3390/antiox11091792