
The Effect of Holder Pasteurization and Different Variants on Breast Milk Antioxidants

 , ,
, ,  ,
, 

Abstract
1. Introduction
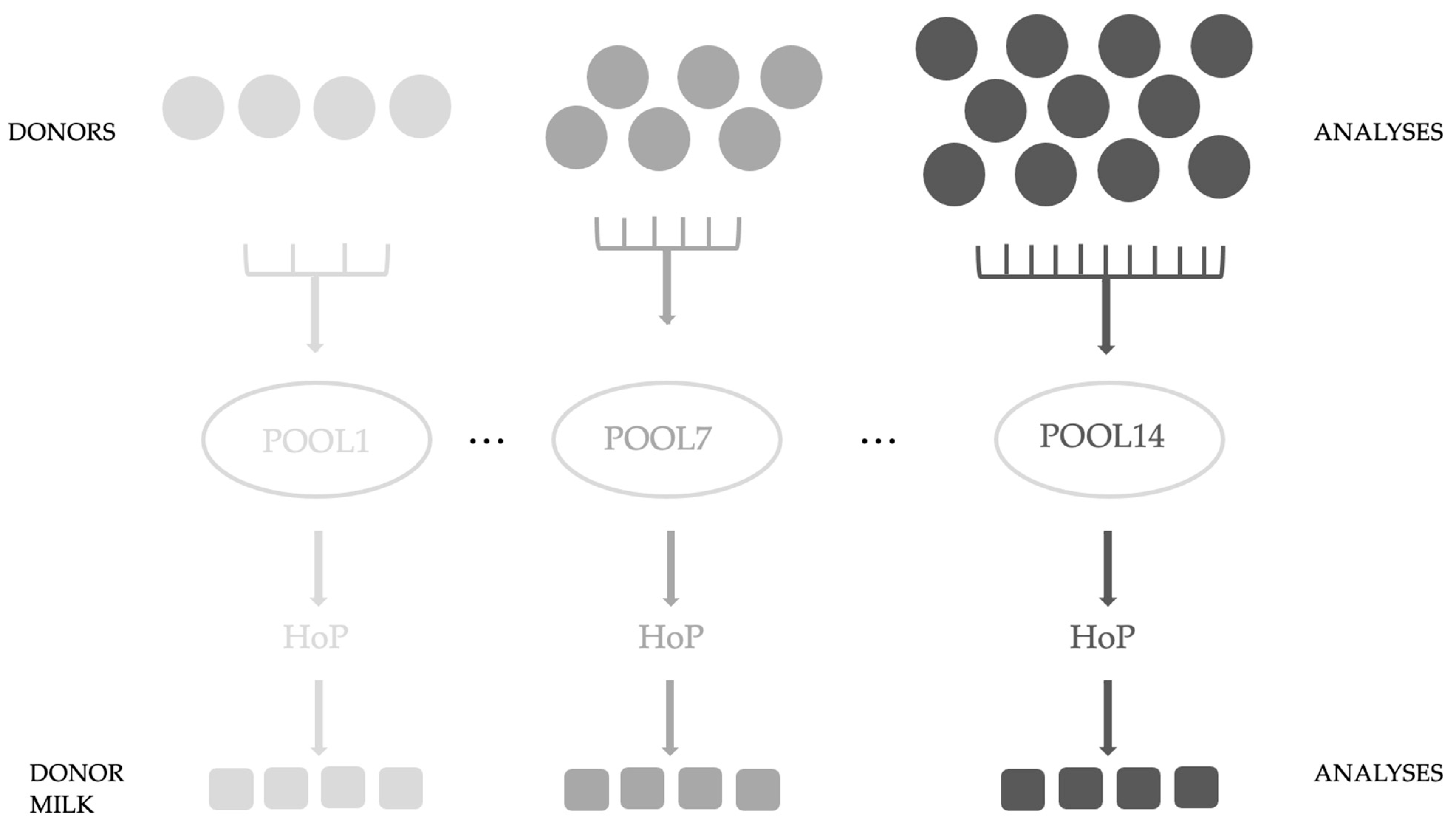
2. Materials and Methods
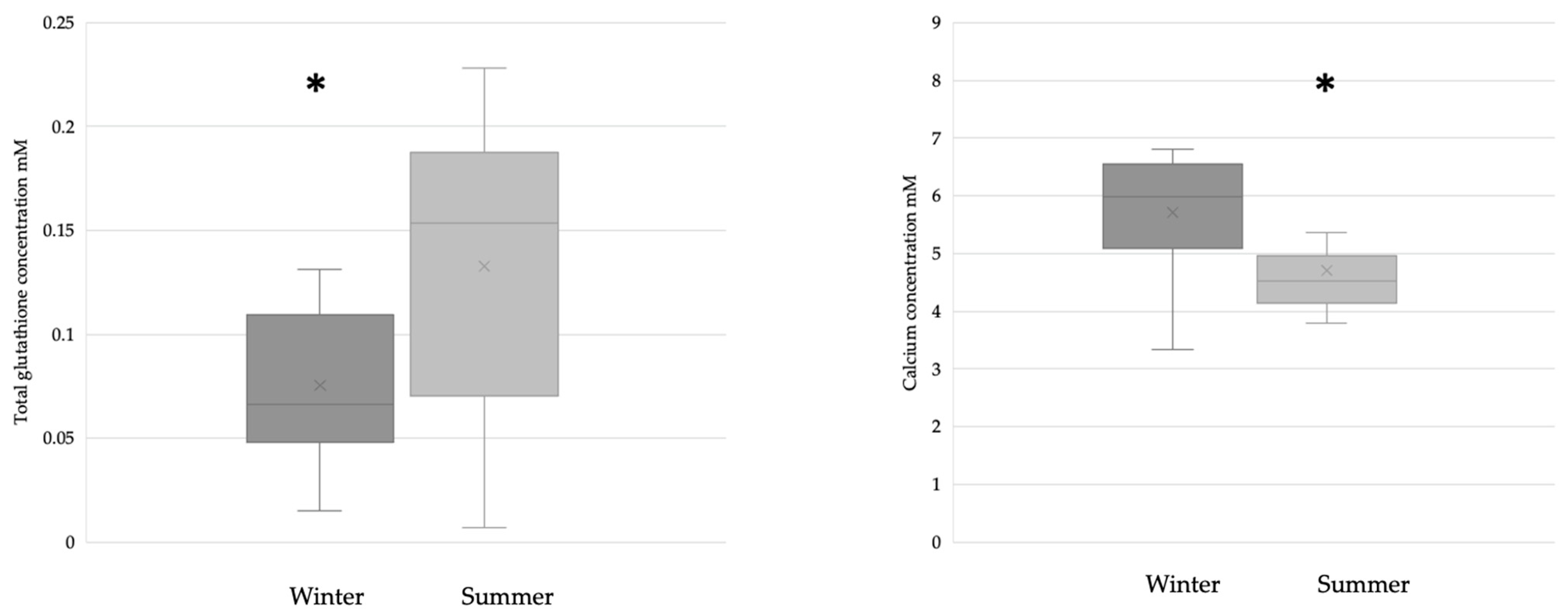
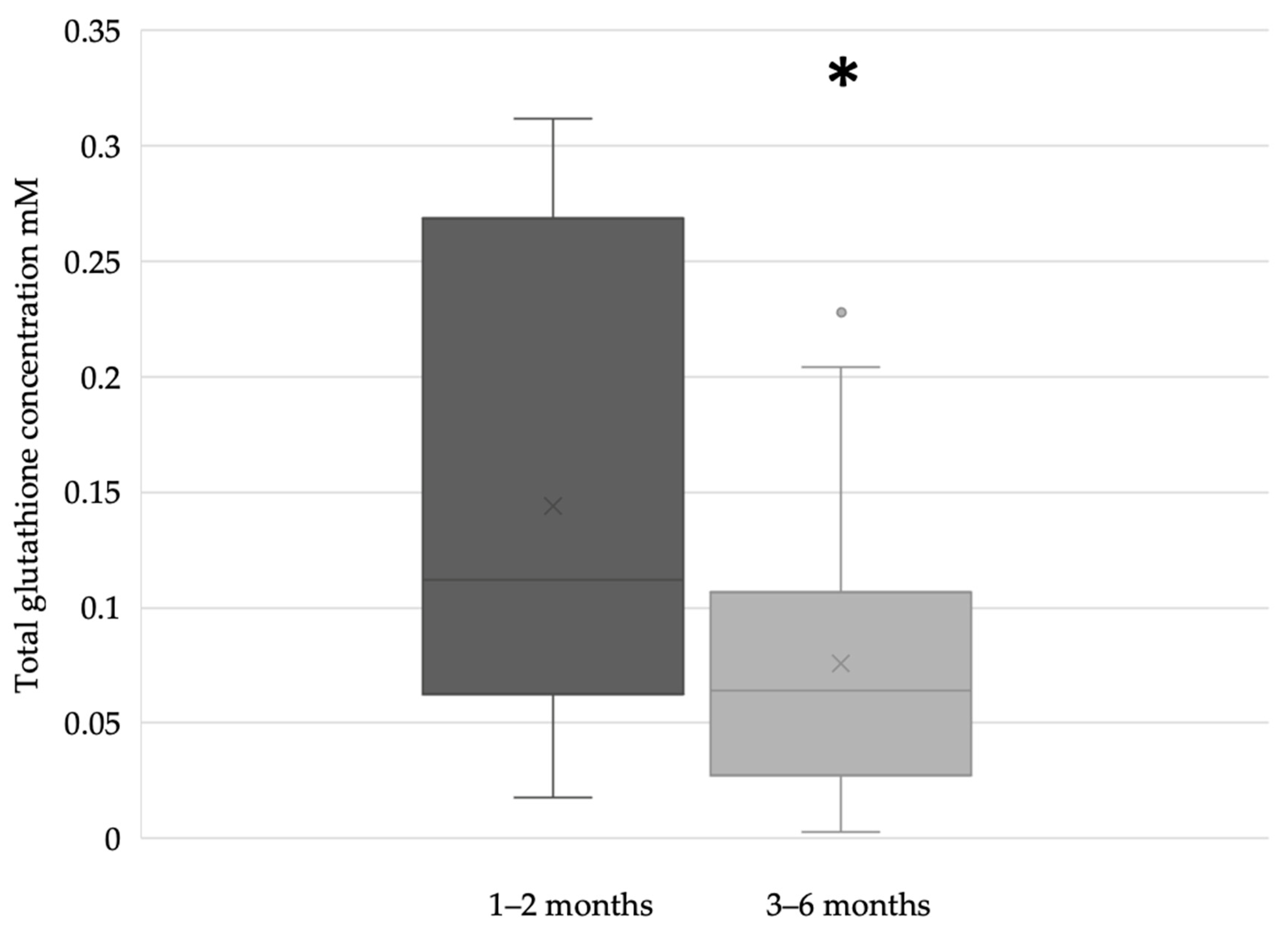
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vass, R.A.; Kiss, G.; Bell, E.F.; Roghair, R.D.; Miseta, A.; Bódis, J.; Funke, S.; Ertl, T. Breast milk for term and preterm infants-own mother’s milk or donor milk? Nutrients 2021, 13, 424. [Google Scholar] [CrossRef]
- Moore, T.A.; Ahmad, I.M.; Zimmerman, M.C. Oxidative stress and preterm birth: An integrative review. Biol. Res. Nurs. 2018, 20, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Mauchart, P.; Vass, R.A.; Nagy, B.; Sulyok, E.; Bódis, J.; Kovács, K. Oxidative stress in assisted reproductive techniques, with a focus on an underestimated risk factor. Curr. Issues Mol. Biol. 2023, 45, 1272–1286. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Lee, E.H.; Kim, E.A. Retinopathy of Prematurity among very-low-birth-weight infants in Korea: Incidence, treatment, and risk factors. J. Korean Med. Sci. 2015, 30, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Tataranno, M.L.; Buonocore, G. Oxidative stress and bronchopulmonary dysplasia. J. Clin. Neonatol. 2012, 1, 109–114. [Google Scholar] [PubMed]
- Silvers, K.M.; Gibson, A.T.; Russell, J.M.; Powers, H.J. Antioxidant activity, packed cell transfusions, and outcome in premature infants. Arch. Dis. Child. Fetal Neonatal Ed. 1998, 78, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Juncker, H.G.; Ruhé, E.J.M.; Burchell, G.L.; van den Akker, C.H.P.; Korosi, A.; van Goudoever, J.B.; van Keulen, B.J. The effect of pasteurization on the antioxidant properties of human milk: A literature review. Antioxidants 2021, 10, 1737. [Google Scholar] [CrossRef]
- Churchill, M.; Zawawi, H.; Elisia, I.; Seider, M.; Noseworthy, R.; Thompson, A.; Glenn, A.J.; Ramdath, D.D.; O’Connor, D.; Darling, P.; et al. The antioxidant capacity of breast milk and plasma of women with or without gestational diabetes mellitus. Antioxidants 2023, 12, 842. [Google Scholar] [CrossRef]
- Cloetens, L.; Panee, J.; Åkesson, B. The antioxidant capacity of milk--the application of different methods in vitro and in vivo. Cell Mol. Biol. 2013, 59, 43–57. [Google Scholar]
- Kőszegi, T.; Sali, N.; Raknić, M.; Horváth-Szalai, Z.; Csepregi, R.; Končić, M.Z.; Papp, N.; Poór, M. A novel luminol-based enhanced chemiluminescence antioxidant capacity microplate assay for use in different biological matrices. J. Pharmacol. Toxicol. Methods 2017, 88, 153–159. [Google Scholar] [CrossRef]
- Golubkova, A.; Leiva, T.; Snyder, K.; Schlegel, C.; Bonvicino, S.M.; Agbaga, M.P.; Brush, R.S.; Hansen, J.M.; Vitiello, P.F.; Hunter, C.J. Response of the glutathione (GSH) antioxidant defense system to oxidative injury in necrotizing enterocolitis. Antioxidants 2023, 12, 1385. [Google Scholar] [CrossRef] [PubMed]
- Di Giacomo, C.; Malfa, G.A.; Tomasello, B.; Bianchi, S.; Acquaviva, R. Natural compounds and glutathione: Beyond mere antioxidants. Antioxidants 2023, 12, 1445. [Google Scholar] [CrossRef] [PubMed]
- Gomme, P.T.; McCann, K.B.; Bertolini, J. Transferrin: Structure, function and potential therapeutic actions. Drug Discov. Today 2005, 10, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Blanc, B.; Isliker, H. Isolation and characterization of the red siderophilic protein from maternal milk: Lactotransferrin. Bull. Soc. Chim. Biol. 1961, 43, 929–943. [Google Scholar]
- Hao, L.; Shan, Q.; Wei, J.; Ma, F.; Sun, P. Lactoferrin: Major physiological functions and applications. Curr. Protein Pept. Sci. 2019, 20, 139–144. [Google Scholar] [CrossRef]
- EFSA NDA. Scientific Opinion on the essential composition of infant and follow-on formulae. EFSA J. 2014, 12, 3760. [Google Scholar] [CrossRef]
- Rios-Leyvraz, M.; Yao, Q. Calcium, zinc, and vitamin D in breast milk: A systematic review and meta-analysis. Int. Breastfeed. J. 2023, 18, 27. [Google Scholar] [CrossRef]
- Vass, R.A.; Kemeny, A.; Dergez, T.; Ertl, T.; Reglodi, D.; Jungling, A.; Tamas, A. Distribution of bioactive factors in human milk samples. Int. Breastfeed. J. 2019, 14, 9. [Google Scholar] [CrossRef]
- Vass, R.A.; Bell, E.F.; Colaizy, T.T.; Schmelzel, M.L.; Johnson, K.J.; Walker, J.R.; Ertl, T.; Roghair, R.D. Hormone levels in preterm and donor human milk before and after Holder pasteurization. Pediatr. Res. 2020, 88, 612–617. [Google Scholar] [CrossRef]
- Vass, R.A.; Roghair, R.D.; Bell, E.F.; Colaizy, T.T.; Schmelzel, M.L.; Johnson, K.J.; Walker, J.R.; Ertl, T. Pituitary glycoprotein hormones in human milk before and after pasteurization or refrigeration. Nutrients 2020, 12, 687. [Google Scholar] [CrossRef]
- Shoji, H.; Shimizu, T. Effect of human breast milk on biological metabolism in infants. Pediatr. Int. 2019, 61, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Vass, R.A.; Kiss, G.; Bell, E.F.; Miseta, A.; Bódis, J.; Funke, S.; Bokor, S.; Molnár, D.; Kósa, B.; Kiss, A.A.; et al. Thyroxine and thyroid-stimulating hormone in own mother’s milk, donor milk, and infant formula. Life 2022, 12, 584. [Google Scholar] [CrossRef] [PubMed]
- Vass, R.A.; Bell, E.F.; Roghair, R.D.; Kiss, G.; Funke, S.; Bokor, S.; Molnár, D.; Miseta, A.; Bódis, J.; Kovács, K.; et al. Insulin, testosterone, and albumin in term and preterm breast milk, donor milk, and infant formula. Nutrients 2023, 15, 1476. [Google Scholar] [CrossRef]
- Montjaux-Regis, N.; Cristini, C.; Arnaud, C.; Glorieux, I.; Vanpee, M.; Casper, C. Improved growth of preterm infants receiving mother’s own raw milk compared with pasteurized donor milk. Acta Paediatr. 2011, 100, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Hard, A.L.; Nilsson, A.K.; Lund, A.M.; Hansen-Pupp, I.; Smith, L.E.H.; Hellstrom, A. Review shows that donor milk does not promote the growth and development of preterm infants as well as maternal milk. Acta Paediatr. 2019, 108, 998–1007. [Google Scholar] [CrossRef]
- Tsopmo, A.; Friel, J.K. Human milk has anti-oxidant properties to protect premature infants. Curr. Pediatr. Rev. 2007, 3, 45–51. [Google Scholar] [CrossRef]
- Friel, J.K.; Martin, S.M.; Langdon, M.; Herzberg, G.R.; Buettner, G.R. Milk from mothers of both premature and full-term infants provides better antioxidant protection than does infant formula. Pediatr. Res. 2002, 51, 612–618. [Google Scholar] [CrossRef]
- Aycicek, A.; Erel, O. Total oxidant/antioxidant status in jaundiced newborns before and after phototherapy. J. Pediatr. 2007, 83, 319–322. [Google Scholar] [CrossRef]
- Ribeiro, V.P.D.; Tinoco, R.B.; Chamon, A.L.B.; Pessoa, I.S.; Santos, T.C.D.; Silva, R.S.; Fronza, M. The influence of time and temperature on human milk storage antioxidant properties, oxidative stress, and total protein. J. Hum. Lact. 2023, 39, 308–314. [Google Scholar] [CrossRef]
- Ankrah, N.A.; Appiah-Opong, R.; Dzokoto, C. Human breastmilk storage and the glutathione content. J. Trop. Pediatr. 2000, 46, 111–113. [Google Scholar] [CrossRef][Green Version]
- Silvestre, D.; Miranda, M.; Muriach, M.; Almansa, I.; Jareño, E.; Romero, F.J. Antioxidant capacity of human milk: Effect of thermal conditions for the pasteurization. Acta Paediatr. 2008, 97, 1070–1074. [Google Scholar] [CrossRef]
- Zarban, A.; Taheri, F.; Chahkandi, T.; Sharifzadeh, G.; Khorashadizadeh, M. Antioxidant and radical scavenging activity of human colostrum, transitional and mature milk. J. Clin. Biochem. Nutr. 2009, 45, 150–154. [Google Scholar] [CrossRef]
- Kusano, C.; Ferrari, B. Total antioxidant capacity: A biomarker in biomedical and nutritional studies. J. Cell. Mol. Biol. 2008, 7, 1–15. [Google Scholar]
- Elisia, I.; Kitts, D.D. Quantification of hexanal as an index of lipid oxidation in human milk and association with antioxidant components. J. Clin. Biochem. Nutr. 2011, 49, 147–152. [Google Scholar] [CrossRef]
- Kitts, D.D.; Hu, C. Biological and chemical assessment of antioxidant activity of sugar-lysine model Maillard reaction products. Ann. N.Y. Acad. Sci. 2005, 1043, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Gila-Díaz, A.; Herranz Carrillo, G.; Cañas, S.; Saenz de Pipaón, M.; Martínez-Orgado, J.A.; Rodríguez-Rodríguez, P.; López de Pablo, Á.L.; Martin-Cabrejas, M.A.; Ramiro-Cortijo, D.; Arribas, S.M. Influence of maternal age and gestational age on breast milk antioxidants during the first month of lactation. Nutrients 2020, 12, 2569. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jackson, R.T.; Khan, S.A.; Ahuja, J.; Pehrsson, P.R. Human milk nutrient composition in the United States: Current knowledge, challenges, and research needs. Curr. Dev. Nutr. 2018, 2, nzy025. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, L.; Bao, W.; Rong, S. Nutritional composition of breast milk in Chinese women: A systematic review. Asia Pac. J. Clin. Nutr. 2018, 27, 491–502. [Google Scholar]
- Peila, C.; Moro, G.E.; Bertino, E.; Cavallarin, L.; Giribaldi, M.; Giuliani, F.; Cresi, F.; Coscia, A. The effect of Holder Pasteurization on nutrients and biologically-active components in donor human milk: A review. Nutrients 2016, 8, 477. [Google Scholar] [CrossRef]
- Gidrewicz, D.A.; Fenton, T.R. A systematic review and meta-analysis of the nutrient content of preterm and term breast milk. BMC Pediatr. 2014, 14, 216. [Google Scholar] [CrossRef]
- Dorea, J.G. Calcium and phosphorus in human milk. Nutr. Res. 1999, 19, 709–739. [Google Scholar] [CrossRef]
- Hinde, K.; Foster, A.B.; Landis, L.M.; Rendina, D.; Oftedal, O.T.; Power, M.L. Daughter dearest: Sex-biased calcium in mother’s milk among rhesus macaques. Am. J. Phys. Anthropol. 2013, 151, 144–150. [Google Scholar] [CrossRef]
- Zhang, M.; Zhai, R.; Liu, J.; Guang, H.; Li, B.; Zhang, S. Seasonal variation of blood calcium levels in children aged 1–10. J. Clin. Lab. Anal. 2016, 30, 741–744. [Google Scholar] [CrossRef]
- Peacock, M. Calcium metabolism in health and disease. Clin. J. Am. Soc. Nephrol. 2010, 1, S23–S30. [Google Scholar] [CrossRef] [PubMed]
- Khoo, A.L.; Koenen, H.J.; Chai, L.Y.; Sweep, F.C.; Netea, M.G.; van der Ven, A.J.; Joosten, I. Seasonal variation in vitamin D(3) levels is paralleled by changes in the peripheral blood human T cell compartment. PLoS ONE 2012, 7, 29250. [Google Scholar] [CrossRef] [PubMed]
- Binder, C.; Baumgartner-Parzer, S.; Gard, L.-I.; Berger, A.; Thajer, A. Human Milk Processing and Its Effect on Protein and Leptin Concentrations. Nutrients 2023, 15, 347. [Google Scholar] [CrossRef]
- Ley, S.H.; Hanley, A.J.; Stone, D.; O’Connor, D.L. Effects of pasteurization on adiponectin and insulin concentrations in donor human milk. Pediatr. Res. 2011, 70, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, U.M.; Siimes, M.A.; Dallman, P.R. Iron absorption in infants: High bioavailability of breast milk iron as indicated by the extrinsic tag method of iron absorption and by the concentration of serum ferritin. J. Pediatr. 1977, 91, 36–39. [Google Scholar] [CrossRef]
- Bullen, J.J.; Rogers, H.J.; Leigh, L. Iron-binding proteins in milk and resistance to Escherichia coli infection in infants. Br. Med. J. 1972, 1, 69–75. [Google Scholar] [CrossRef]
- Wieringa, F.T.; Berger, J.; Dijkhuizen, M.A.; Hidayat, A.; Ninh, N.X.; Utomo, B.; Wasantwisut, E.; Winichagoon, P. Sex differences in prevalence of anaemia and iron deficiency in infancy in a large multi-country trial in South-East Asia. Br. J. Nutr. 2007, 98, 1070–1076. [Google Scholar] [CrossRef]
- Alam, F.; Memon, A.S.; Fatima, S.S. Increased body mass index may lead to hyperferritinemia irrespective of body iron stores. Pak. J. Med. Sci. 2015, 31, 1521–1526. [Google Scholar] [PubMed]
- Keikha, M.; Shayan-Moghadam, R.; Bahreynian, M.; Kelishadi, R. Nutritional supplements and mother’s milk composition: A systematic review of interventional studies. Int. Breastfeed. J. 2021, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Cortijo, D.; Herranz Carrillo, G.; Singh, P.; Rebollo-Hernanz, M.; Rodríguez-Rodríguez, P.; Ruvira, S.; Martín-Trueba, M.; Martin, C.R.; Arribas, S.M. Maternal and neonatal factors modulating breast milk cytokines in the first month of lactation. Antioxidants 2023, 12, 996. [Google Scholar] [CrossRef]
- Bermejo-Haro, M.Y.; Camacho-Pacheco, R.T.; Brito-Pérez, Y.; Mancilla-Herrera, I. The hormonal physiology of immune components in breast milk and their impact on the infant immune response. Mol. Cell Endocrinol. 2023, 572, 111956. [Google Scholar] [CrossRef] [PubMed]
- Petrullo, L.; Hinde, K.; Lu, A. Steroid hormone concentrations in milk predict sex-specific offspring growth in a nonhuman primate. Am. J. Hum. Biol. 2019, 31, e23315. [Google Scholar] [CrossRef]
- Czlapka-Matyasik, M.; Gramza-Michalowska, A. The total dietary antioxidant capacity, its seasonal variability, and dietary sources in cardiovascular patients. Antioxidants 2023, 12, 292. [Google Scholar] [CrossRef]
- Simon Sarkadi, L.; Zhang, M.; Muránszky, G.; Vass, R.A.; Matsyura, O.; Benes, E.; Vari, S.G. Fatty acid composition of milk from mothers with normal weight, obesity, or gestational diabetes. Life 2022, 12, 1093. [Google Scholar] [CrossRef]
- Ramiro-Cortijo, D.; Singh, P.; Liu, Y.; Medina-Morales, E.; Yakah, W.; Freedman, S.D.; Martin, C.R. Breast milk lipids and fatty acids in regulating neonatal intestinal development and protecting against intestinal injury. Nutrients 2020, 12, 534. [Google Scholar] [CrossRef]
- Bokor, S.; Vass, R.A.; Funke, S.; Ertl, T.; Molnár, D. Epigenetic effect of maternal methyl-group donor intake on offspring’s health and disease. Life 2022, 12, 609. [Google Scholar] [CrossRef]
- Dallas, D.C.; Underwood, M.A.; Zivkovic, A.M.; German, J.B. Digestion of protein in premature and term Infants. J. Nutr. Disord. Ther. 2012, 2, 112. [Google Scholar] [CrossRef]
- Pozzo, L.; Cirrincione, S.; Russo, R.; Karamać, M.; Amarowicz, R.; Coscia, A.; Antoniazzi, S.; Cavallarin, L.; Giribaldi, M. Comparison of oxidative status of human milk, human milk fortifiers and preterm infant formulas. Foods 2019, 8, 458. [Google Scholar] [CrossRef]
- Turai, R.; Schandl, M.F.; Dergez, T.; Vass, R.A.; Kvárik, T.; Horányi, E.; Balika, D.; Mammel, B.; Gyarmati, J.; Fónai, F.; et al. Early and late complications of hyperglycemic extremely low birth-weight infants. Orv. Hetil. 2019, 160, 1270–1278. [Google Scholar] [CrossRef]
- Roghair, R.D.; Colaizy, T.T.; Steinbrekera, B.; Vass, R.A.; Hsu, E.; Dagle, D.; Chatmethakul, T. Neonatal leptin levels predict the early childhood developmental assessment scores of preterm infants. Nutrients 2023, 15, 1967. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.M.; Turner, M.A.; Weindling, A.M. Neurodevelopmental outcomes at 42 months after thyroxine supplementation in infants below 28 weeks’ gestation: A randomized controlled trial. Thyroid 2020, 30, 948–954. [Google Scholar] [CrossRef]
- Sulyok, E.; Farkas, B.; Bodis, J. Pathomechanisms of prenatally programmed adult diseases. Antioxidants 2023, 12, 1354. [Google Scholar] [CrossRef]
- Barker, D.J.P. In utero programming of chronic disease. Clin. Sci. 1998, 95, 115–128. [Google Scholar] [CrossRef]
- Barker, D.J.; Osmond, C.; Forsén, T.J.; Kajantie, E.; Eriksson, J.G. Trajectories of growth among children who have coronary events as adults. N. Engl. J. Med. 2005, 353, 1802–1809. [Google Scholar] [CrossRef]
- Kuzawa, C.W. Fetal origins of developmental plasticity: Are fetal cues reliable predictors of future nutritional environment? Am. J. Hum. Biol. 2005, 17, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.G.A.; Yin, Y.; Yang, H.; Chughtai, M.I.; et al. The role of oxidative stress and antioxidant balance in pregnancy. Mediat. Inflamm. 2021, 2021, 9962860. [Google Scholar] [CrossRef]
- Mistry, H.D.; Williams, P.J. The importance of antioxidant micronutrients in pregnancy. Oxid. Med. Cell. Longev. 2011, 2011, 841749. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hu, R.; Shi, M.; Wang, L.; Yan, J.; Gong, J.; Zhang, Q.; He, J.; Wu, S. Placental malfunction, fetal survival and development caused by sow metabolic disorder: The impact of maternal oxidative stress. Antioxidants 2023, 12, 360. [Google Scholar] [CrossRef] [PubMed]
- Musilova, I.; Tothova, L.; Menon, R.; Vlkova, B.; Celec, P.; Hornychova, H.; Kutova, R.; Andrys, C.; Stepan, M.; Kacerovsky, M. Umbilical cord blood markers of oxidative stress in pregnancies complicated by preterm prelabor rupture of membranes. J. Matern. Fetal Neonatal Med. 2016, 29, 1900–1910. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Huang, P.; Lyu, M.; Dong, J. Oxidative stress and preeclampsia-associated prothrombotic state. Antioxidants 2020, 9, 1139. [Google Scholar] [CrossRef]
- Marseglia, L.; D’Angelo, G.; Manti, S.; Arrigo, T.; Barberi, I.; Reiter, R.J.; Gitto, E. Oxidative stress-mediated aging during the fetal and perinatal periods. Oxid. Med. Cell Longev. 2014, 2014, 358375. [Google Scholar] [CrossRef]
- DeFreitas, M.J.; Katsoufis, C.P.; Benny, M.; Young, K.; Kulandavelu, S.; Ahn, H.; Sfakianaki, A.; Abitbol, C.L. Educational Review: The Impact of Perinatal Oxidative Stress on the Developing Kidney. Front. Pediatr. 2022, 10, 853722. [Google Scholar] [CrossRef]
- Sultana, Z.; Maiti, K.; Aitken, J.; Morris, J.; Dedman, L.; Smith, R. Oxidative stress placental ageing-related pathogenesis and adverse pregnancy outcomes. Am. J. Reprod. Immunol. 2017, 77, 12653. [Google Scholar] [CrossRef]
- Gluckman, P.D. Editorial. nutrition, glucocorticoid, birth size and adult disease. J. Clin. Endocrinol. Metab. 2001, 142, 1689–1691. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kanaka-Gantenbein, C. Fetal origins of adult diabetes. Ann. N.Y. Acad. Sci. 2010, 1205, 99–105. [Google Scholar] [CrossRef]
- Symonds, M.E.; Stephenson, T.; Gardner, D.S.; Budge, H. Long-term effects of nutritional programming of the embryo and fetus. mechanisms and critical windows. Reprod. Fertil. Dev. 2007, 19, 53–63. [Google Scholar] [CrossRef]
- Ross, M.G.; Desai, M. Developmental programming of offspring obesity, adipogenesis and appetite. Clin. Obstet. Gynecol. 2013, 56, 529–536. [Google Scholar] [CrossRef]
- Chrustek, A.; Dombrowska-Pali, A.; Olszewska-Slonina, D. Analysis of the composition and antioxidant status of breast milk in women giving birth prematurely and on time. PLoS ONE 2021, 16, 0255252. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, S.; Deng, X.; Luo, Z.; Chen, A.; Yu, R. Effects of antioxidants in human milk on bronchopulmonary dysplasia prevention and treatment: A review. Front Nutr 2022, 9, 924036. [Google Scholar] [CrossRef] [PubMed]
- Rudloff, I.; Cho, S.X.; Bui, C.B.; McLean, C.; Veldman, A.; Berger, P.J.; Nold, M.F.; Nold-Petry, C.A. Refining anti-inflammatory therapy strategies for bronchopulmonary dysplasia. J. Cell Mol. Med. 2017, 21, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Li, Z.; Ai, D.; Ma, J.; Chen, C.; Liu, X. Interleukin-24 as a pulmonary target cytokine in bronchopulmonary dysplasia. Cell Biochem. Biophys. 2021, 79, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Altobelli, E.; Angeletti, P.M.; Verrotti, A.; Petrocelli, R. The impact of human milk on necrotizing enterocolitis: A systematic review and meta-analysis. Nutrients 2020, 12, 1322. [Google Scholar] [CrossRef]
- Sami, A.S.; Frazer, L.C.; Miller, C.M.; Singh, D.K.; Clodfelter, L.G.; Orgel, K.A.; Good, M. The role of human milk nutrients in preventing necrotizing enterocolitis. Front. Pediatr. 2023, 11, 1188050. [Google Scholar] [CrossRef]
- Tang, W.; Gao, T.; Cao, Y.; Zhou, W.; Song, D.; Wang, L. Narrative review of perinatal management of extremely preterm infants: What’s the evidence? Pediatr. Med. 2022, 5, 37. [Google Scholar] [CrossRef]
- Siffel, C.; Hirst, A.K.; Sarda, S.P.; Kuzniewicz, M.W.; Li, D.K. The clinical burden of extremely preterm birth in a large medical records database in the United States: Mortality and survival associated with selected complications. Early Hum. Dev. 2022, 171, 105613. [Google Scholar] [CrossRef]
- Balázs, G.; Balajthy, A.; Seri, I.; Hegyi, T.; Ertl, T.; Szabó, T.; Röszer, T.; Papp, Á.; Balla, J.; Gáll, T.; et al. Prevention of chronic morbidities in extremely premature newborns with LISA-nCPAP respiratory therapy and adjuvant perinatal strategies. Antioxidants 2023, 12, 1149. [Google Scholar] [CrossRef]
- Vohr, B.R.; Poindexter, B.B.; Dusick, A.M.; McKinley, L.T.; Wright, L.L.; Langer, J.C.; Poole, W.K.; NICHD Neonatal Research Network. Beneficial effects of breast milk in the neonatal intensive care unit on the developmental outcome of extremely low birth weight infants at 18 months of age. Pediatrics 2006, 118, 115–123. [Google Scholar] [CrossRef]
- Vohr, B.R.; Poindexter, B.B.; Dusick, A.M.; McKinley, L.T.; Higgins, R.D.; Langer, J.C.; Poole, W.K.; National Institute of Child Health and Human Development National Research Network. Persistent beneficial effects of breast milk ingested in the neonatal intensive care unit on outcomes of extremely low birth weight infants at 30 months of age. Pediatrics 2007, 120, e953–e959. [Google Scholar] [CrossRef]
- Arslanoglu, S.; Boquien, C.Y.; King, C.; Lamireau, D.; Tonetto, P.; Barnett, D.; Bertino, E.; Gaya, A.; Gebauer, C.; Grovslien, A.; et al. Fortification of human milk for preterm infants: Update and recommendations of the European Milk Bank Association (EMBA) working group on human milk fortification. Front. Pediatr. 2019, 7, 76. [Google Scholar] [CrossRef]
- Sudeep, K.C.; Kumar, J.; Ray, S.; Dutta, S.; Aggarwal, R.; Kumar, P. Oral Application of colostrum and mother’s own milk in preterm infants-a randomized, controlled trial. Indian J. Pediatr. 2022, 89, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Lembo, C.; Buonocore, G.; Perrone, S. Oxidative stress in preterm newborns. Antioxidants 2021, 10, 1672. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.; Soulimane Mokhtari, N.; Merzouk, S.A.; Merzouk, H.; Belarbi, B.; Narce, M. Oxidant and antioxidant status in mothers and their newborns according to birthweight. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 141, 95–99. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Raw | HoP | p-Value | |
|---|---|---|---|
| Glutathione (mM) (n = 112) | 0.11 ± 0.02 | 0.12 ± 0.10 | 0.575 |
| TAC-ECL (μM) (n = 59) | 127.31 ± 6.24 | 111.27 ± 5.26 | 0.028 |
| TAC-ORAC (μM) (n = 59) | 3602.34 ± 104.13 | 3163.13 ± 787.94 | 0.001 |
| Calcium (mM) (n = 59)) | 5.08 ± 0.15 | 6.38 ± 0.10 | <0.0001 |
| Total protein (g/L) (n = 59) | 4.91 ± 0.16 | 3.86 ± 0.11 | <0.0001 |
| Transferrin (mg/L) (n = 59) | 80.82 ± 10.85 | 1.34 ± 0.09 | <0.0001 |
| Glutathione mM | TAC µM | ORAC µM | Calcium mM | Total Protein g/L | Transferrin mg/L | ||
|---|---|---|---|---|---|---|---|
| Infant gender | Girl | 0.08 ± 0.01 (n = 57) | 134.06 ± 8.08 (n = 27) | 3752.71 ± 122.34 (n = 27) | 5.21 ± 0.21 (n = 27) | 5.02 ± 0.22 (n = 27) | 108.39 ± 16.62 (n = 27) |
| Boy | 0.15 ± 0.05 (n = 65) | 114.82 ± 9.83 (n = 32) | 3406.66 ± 192.41 (n = 32) | 4.78 ± 0.22 (n = 32) | 4.74 ± 0.27 (n = 32) | 44.22 ± 8.54 * (n = 32) | |
| Delivery | Vaginal delivery | 0.13 ± 0.04 (n = 70) | 142.07 ± 7.96 (n = 25) | 3733.45 ± 149.79 (n = 25) | 5.05 ± 0.25 (n = 25) | 5.07 ± 0.25 (n = 25) | 69.1 ± 16.5 (n = 25) |
| C-section | 0.08 ± 0.01 (n = 52) | 109.94 ± 8.75 * (n = 34) | 3507.41 ± 153.97 (n = 34) | 5.02 ± 0.17 (n = 34) | 4.77 ± 0.24 (n = 34) | 94.16 ± 15.68 (n = 34) | |
| Maternal BMI | <30 | 0.12 ± 0.03 (n = 90) | 130.21 ± 7.07 (n = 41) | 3623.63 ± 132.36 (n = 41) | 4.99 ± 0.21 (n = 41) | 4.84 ± 0.23 (n = 41) | 52.8 ± 7.94 (n = 41) |
| >30 | 0.07 ± 0.01 (n = 32) | 105.97 ± 13.83 (n = 18) | 3605.48 ± 192.65 (n = 18) | 5.12 ± 0.24 (n = 18) | 5.06 ± 0.24 (n = 18) | 139.87 ± 24.30 * (n = 18) | |
| Maternal age | <30 | 0.06 ± 0.01 (n = 51) | 99.61 ± 9.36 (n = 34) | 3690.39 ± 193.75 (n = 34) | 5.15 ± 0.28 (n = 34) | 4.65 ± 0.25 (n = 34) | 81.63 ± 20.04 (n = 34) |
| >30 | 0.11 ± 0.01 * (n = 71) | 134.07 ± 7.56 * (n = 25) | 3592.23 ± 134.22 (n = 25) | 4.99 ± 0.19 (n = 25) | 5.01 ± 0.22 (n = 25) | 82.76 ± 13.93 (n = 25) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vass, R.A.; Mikó, É.; Gál, C.; Kőszegi, T.; Vass, C.I.; Bokor, S.; Molnár, D.; Funke, S.; Kovács, K.; Bódis, J.; et al. The Effect of Holder Pasteurization and Different Variants on Breast Milk Antioxidants. Antioxidants 2023, 12, 1857. https://doi.org/10.3390/antiox12101857
Vass RA, Mikó É, Gál C, Kőszegi T, Vass CI, Bokor S, Molnár D, Funke S, Kovács K, Bódis J, et al. The Effect of Holder Pasteurization and Different Variants on Breast Milk Antioxidants. Antioxidants. 2023; 12(10):1857. https://doi.org/10.3390/antiox12101857
Chicago/Turabian StyleVass, Réka Anna, Éva Mikó, Csenge Gál, Tamás Kőszegi, Csaba I. Vass, Szilvia Bokor, Dénes Molnár, Simone Funke, Kálmán Kovács, József Bódis, and et al. 2023. "The Effect of Holder Pasteurization and Different Variants on Breast Milk Antioxidants" Antioxidants 12, no. 10: 1857. https://doi.org/10.3390/antiox12101857
APA StyleVass, R. A., Mikó, É., Gál, C., Kőszegi, T., Vass, C. I., Bokor, S., Molnár, D., Funke, S., Kovács, K., Bódis, J., & Ertl, T. (2023). The Effect of Holder Pasteurization and Different Variants on Breast Milk Antioxidants. Antioxidants, 12(10), 1857. https://doi.org/10.3390/antiox12101857