
Cosmosiin Induces Apoptosis in Colorectal Cancer by Inhibiting PD-L1 Expression and Inducing ROS
by
 , , , and
, , , and
Jung Ho Han
1,
Eun-Ji Lee
1,
Wonyoung Park
2,
Jang-Gi Choi
1,
Ki-Tae Ha
2 and
Hwan-Suck Chung
1,3,* 1
Korean Medicine (KM)-Application Center, Korea Institute of Oriental Medicine (KIOM), Daegu 41062, Republic of Korea
2
Department of Korean Medical Science, School of Korean Medicine, Pusan National University, Yangsan 50612, Republic of Korea
3
Korean Convergence Medical Science Major, University of Science and Technology (UST), KIOM Campus, Daegu 41062, Republic of Korea
*
Author to whom correspondence should be addressed.
Antioxidants 2023, 12(12), 2131; https://doi.org/10.3390/antiox12122131
Submission received: 31 October 2023
/
Revised: 7 December 2023
/
Accepted: 13 December 2023
/
Published: 18 December 2023
(This article belongs to the Section Natural and Synthetic Antioxidants)
Abstract
:Immunotherapies, particularly those concerning immune checkpoint inhibitors, have transformed cancer treatment in recent years. Programmed death-ligand 1 (PD-L1) is a key target for immunotherapy that is overexpressed in the cells of colorectal cancer, a widespread malignant cancer that poses a significant healthcare challenge. This study investigated the effects of cosmosiin treatment on colorectal cancer cell lines. Cosmosiin is a naturally occurring flavone glycoside compound that has potential health benefits, including antioxidant and immunomodulatory effects. This study showed that cosmosiin effectively suppresses the expression of PD-L1 and triggers apoptosis, which is facilitated through pathways that are related to reactive oxygen species. These outcomes suggest that cosmosiin could be a promising candidate for an immune checkpoint inhibitor in the treatment of colorectal cancer.
1. Introduction
Colorectal cancer (CRC) is among the most common forms of malignancy worldwide, imposing a significant global health burden [1]. Although there have been advances in its diagnosis and treatment, CRC remains a formidable challenge, requiring the further exploration of novel therapeutic strategies [2]. Immunotherapy, in particular approaches involving immune checkpoint inhibitors (ICIs), has transformed cancer treatment in recent years, showing remarkable success in treating a range of tumor types [3]. One key target in immunotherapy is programmed death-ligand 1 (PD-L1), a cell-surface protein that plays a pivotal role in evading immune surveillance and promoting tumor immune escape [4]. PD-L1 expression in tumor cells and immune cells in the tumor microenvironment has been found to be associated with immune evasion and disease progression in several malignancies [5]. As understanding of the complex interplay between cancer cells and the immune system grows, the investigation of the link between colon cancer and PD-L1 expression has emerged as a compelling area of research, with potential clinical implications [6].
Cosmosiin (COS) has synonyms, including apigetrin and apigenin-7-glucoside, a naturally occurring flavone glycoside compound [7,8]. COS has been used in traditional medicine systems for its health benefits, and its chemical structure has revealed that it possesses pharmacological properties that have therapeutic implications [8,9,10,11]. Preliminary research has suggested that this compound exhibits a wide array of bioactivities, including antioxidant and immunomodulatory activities. Notably, this compound was among the primary constituents of Salvia plebeia, a medicinal herb employed in prior investigations [12]. The growing demand for safe and effective therapeutic agents has led to an increased focus on alternative treatments, including the study of natural compounds such as COS [13,14]. COS has significant promise as a potential pharmaceutical agent due to its intriguing chemical structure and its demonstrated pharmacological activities [15]. Understanding the mechanisms of its action and evaluating its safety profile are crucial steps for harnessing its therapeutic potential for various diseases. In this context, COS has emerged as a particularly intriguing natural compound with versatile pharmacological properties, which make it a compelling candidate for the modulation of PD-L1 expression in cancer cells. The overexpression of PD-L1 in cancer cells is implicated in immune evasion and tumor progression, making this an attractive target for a therapeutic intervention [16]. Within the realm of cancer immunotherapy, a burgeoning interest has arisen in exploring natural compounds as potential PD-L1 modulators [17]. These compounds, which are derived from a range of plant sources, possess an array of bioactive properties and are being extensively studied for their potential in cancer therapy [18]. This research investigated the impact of controlling PD-L1 expression in cancer, with a focus on the use of natural compounds as promising drug candidates for unlocking novel avenues in cancer treatment and improving patient outcomes.
Several articles have highlighted the significance of PD-L1 expression in cancer progression and have demonstrated its correlation with the efficacy of ICIs [19,20]. Interestingly, a growing body of evidence supports the notion that a single compound, acting as an ICI, can both induce reactive oxygen species (ROS) and trigger apoptosis in cancer cells concurrently [21]. These findings have sparked considerable interest in exploring the therapeutic implications of ROS-mediated apoptosis as a mechanism for modulating PD-L1 expression and enhancing the effectiveness of cancer immunotherapy [22,23]. This thesis builds upon these intriguing observations, investigating the interplay between ROS, apoptosis, and PD-L1 expression, with a specific focus on the use of a unique natural compound as a potential ICI in cancer treatment. However, there have not been any reports on the use of COS to regulate PD-L1 expression in CRC. In this study, we investigated the effects of COS treatment on PD-L1 expression and related pathways, such as ROS and apoptosis, to determine its potential as an ICI and a promising therapeutic agent for CRC.
2. Materials and Methods
2.1. Materials
A cell counting kit-8 (CCK-8) was purchased from Dojindo Molecular Technologies (#CK04, Rockville, MD, USA). COS was purchased from ChemFaces (#CFN98981, purity ≥ 98%, Wuhan, China). Human interferon-gamma (IFN-γ) Recombinant Protein was purchased from R&D Systems (#285-IF-100, purity ≥ 97%, Minneapolis, MN, USA). Crystal violet was purchased from Sigma-Aldrich (#V5265, St. Louis, MO, USA), and 2’,7’-dichlorodihydrofluorescein diacetate (DCFH-DA) was purchased from Thermo Fisher Scientific (#D399, Waltham, MA, USA). Antibodies against PD-L1, p-AKT, AKT, p-ERK, ERK, PARP, and cleaved-PARP were purchased from Cell Signaling Technology (#13684S, #4060S, #4691S, #4370S, #4695S, #9542S, #9541S, Danvers, MA, USA). Antibodies targeting B-cell lymphoma-2 (Bcl-2) and Bcl-2-associated X (BAX) were acquired from Novus Biologicals (#NBP2-34443, #NBP1-88682, Littleton, CO, USA). Antibodies against glycoceramide 3-phosphate dehydrogenase (GAPDH) were purchased from Santa Cruz Biotechnology (#sc-47724, Santa Cruz, CA, USA).
2.2. Cell Culture
DLD-1, Hct116, RKO, and Jurkat T cells were purchased from the American Type Culture Collection (Manassas, VA, USA). DLD-1, Hct116, and RKO cells are human colorectal cancer cell lines, and Table 1 displays the gene types of representative oncogenes [24,25,26]. DLD-1 and Jurkat T cells were cultured in the Roswell Park Memorial Institute 1640 Medium (RPMI 1640), and Hct116 and RKO cells were cultured in Dulbecco’s modified Eagle’s medium containing 10% heat-inactivated fetal bovine serum and 1% penicillin/streptomycin. All of the cells were cultured in a humidified CO2 incubator at 37 °C and 5% CO2. Cell-culture-related solutions were purchased from Hyclone Laboratories, Inc. (GE Healthcare Life Sciences, Chicago, IL, USA).
2.3. Cell Viability Assay
The cell viability assay was conducted using a CCK-8 assay. The DLD-1, RKO, and Hct116 cells were seeded in a 96-well plate at a density of 5 × 103 cells/well and were cultured with the specified dose of COS for the designated duration. After the treatment period, the CCK-8 reagent was added, following the manufacturer’s protocol. The cells were then incubated for 1–2 h at 37 °C and at 5% CO2 in a cell culture incubator. The absorbance was measured at 450 nm with a Spectramax M2 Microplate Reader (Molecular Devices, Sunnyvale, CA, USA).
2.4. Western Blot Analysis
The cells were rinsed using phosphate-buffered saline (PBS), and the total cellular proteins were extracted using a RIPA buffer along with a 1% NP-40 lysis buffer, including protease inhibitor cocktail tablets (Roche, Basel, Switzerland). The protein levels in the samples were measured using the Bio-Rad protein assay. The uniform protein quantities were separated using 8–15% SDS-PAGE in all samples, followed by the electrophoretic transfer of proteins onto nitrocellulose membranes (GE Healthcare, Munich, Germany). Following the blocking of the membranes at room temperature (20–25 °C) for 1 h with 1–5% nonfat dry milk or 1–5% bovine serum albumin, they were subjected to overnight incubation with primary antibodies at 4 °C. Following this, the membranes underwent washing three times with Tris-buffered saline for 10 min each. The Bio-Rad Chemidoc imaging system (Bio-Rad, Hercules, CA, USA) was employed to measure the specific protein bands. The protein expression levels were normalized against GAPDH.
2.5. Flow Cytometry
PD-L1 expression and apoptotic cell analysis were conducted using flow cytometry with APC anti-human CD274 antibody (B7-H1, PD-L1; #393610, BioLegend, San Diego, CA, USA) and a FITC Annexin V Apoptosis Detection Kit I (#556547; BD Biosciences, San Jose, CA, USA), respectively. The cells were stained with the antibody and an Annexin V-FITC Apoptosis Detection Kit following the manufacturer’s instructions. The fluorescence of stained cells was measured in the FITC and PE channels, gating the living and apoptosis cells. The quantification of PD-L1 expression and the ratio of apoptotic cells were assessed using a CytoFLEX flow cytometer (Beckman Coulter Inc., Pasadena, CA, USA).
2.6. Co-Culture Experiments
Jurkat T cells transduced with a PD-1-expressing lentivirus were utilized, and PD-1 expression has been confirmed via FACS analysis in a previous study [10]. Jurkat T cells were activated using human activator beads (Dynabeads™ Human T-Activator CD3/CD28, Thermo Fisher Scientific, Waltham, MA, USA). The cancer cells were seeded at a density of 1 × 105 cells in 12-well plates and were allowed to attach overnight. Then, a pretreatment with COS at the indicated dose was administered for 4 h, followed by the addition of 20 ng/mL of IFN-γ. The activated Jurkat T cells were cocultured with cancer cells over a 2-day period, during which the coculture was incubated at 37 °C and 5% CO2 in a cell culture incubator. Following the 2-day coculture, the supernatants were collected for cytokine measurements. The plates were washed with PBS, and the cells were stained using a crystal violet solution. Following staining, the crystal violet solution was removed, the plates were dried, and the extent of staining was quantified.
2.7. Cytokine Measurement
The medium was obtained from the coculture experiments. Then, the cells were removed by centrifuge, and cell-free media were obtained. The level of interleukin-2 (IL-2, #555,148, BD Biosciences, San Jose, CA, USA) in the cell-free media was measured using an ELISA kit. Each ELISA kit was measured according to the manufacturer’s instructions.
2.8. Determination of Intracellular Reactive Oxygen Species
The intracellular ROS levels were assessed using flow cytometry assays. Cells were seeded in six-well plates at a density of 1–5 × 105 cells per well and were subjected to treatment with the indicated concentrations of COS and N-acetylcysteine (NAC). Following a 2-day incubation period, the cells were washed three times with prewarmed 1 xPBS. A working solution of 0.1 µM DCFH-DA in PBS was prepared, shielded from light, and then incubated at 37 °C for 30 min. Following incubation, cells were washed three times with prewarmed DPBS and subsequently resuspended. The levels of ROS were quantified using a CytoFLEX flow cytometer. Intracellular ROS fluorescence images were obtained with a fluorescence microscope (Nikon ECLIPSE Ti-U, Nikon Co., Tokyo, Japan), and intensity was interpreted as the intracellular ROS level.
2.9. Statistical Analysis
The data for cell viability and PD-L1 expression were obtained by flow cytometry assays and coculture experiments, and intracellular ROS was expressed as ratios relative to the corresponding control values. The results were presented as the means ± standard errors of the mean (SEM) from three experimental replicates. Variations between the mean values within each group were analyzed using a Student’s t-test. The differences among multiple groups were assessed using a one-way analysis of variance, followed by a Tukey’s post hoc examination. All statistical analyses were performed using GraphPad Prism version 8.0 (GraphPad Software, San Diego, CA, USA) to ensure accurate data-handling practices.
3. Results
3.1. Cell Viability on Colon Cancer Cells by COS Treatments
Figure 1A illustrates the structural formula of COS, a compound pivotal to our study. Cell viability assays were performed to determine the toxicity and effectiveness of COS on colon cancer cells. The colorectal cell lines DLD-1, RKO, and Hct116 were treated with COS at the indicated doses for 2 days. After treatment, COS significantly decreased cell viability in all three cell lines. The 50% growth inhibition (GI50) values were 34.29 µM, 63.67 µM, and 50.37 µM for DLD-1, RKO, and Hct116 cells, respectively (Figure 1B). By contrast, no significant difference in cell viability was observed with the IFN-γ treatment for 2 days (Figure 1C).
3.2. COS Inhibits the PD-L1 Expression of Colon Cancer Cells
We have previously shown that Salvia plebeia R. Br. extract and its major compound COS blockade PD-1/PD-L1 interaction [10]. We hypothesized that COS would have the function of inhibiting PD-L1 expression in colon cancer cells. From the results in Figure 1, we determined the treatment conditions of COS (5 µM) and IFN-γ (20 ng/mL) and confirmed changes in PD-L1 expression using Western blot assay. As reported elsewhere, IFN-γ expression increases in a dose-dependent manner, while COS expression was suppressed (Figure 2A,B). When COS and IFN-γ were treated in combination, PD-L1 expression was reduced (Figure 2C). The same results were seen when measured with FACS (Figure 2D). These results suggest that COS suppresses PD-L1 expression.
Considering the critical role of PD-L1 expression in cancer cell immune escape, our study investigated the impact of COS on T-cell-mediated cell death in colon cancer cells, based on its observed effect (Figure 2) on PD-L1 expression regulation. To achieve this, we conducted a series of viability assays involving DLD-1, RKO, and Hct116 colon cancer cells, using Jurkat T cells that expressed PD-1 [10]. These colon cancer cells were subjected to various treatments, including individual exposures to COS (5 µM) and IFN-γ (20 ng/mL), as well as cotreatments involving Jurkat T cells at ratios of 1:10 and 1:20. Our experimental results indicated several important trends. IFN-γ treatment was associated with increased cell viability, while COS treatment led to reduced viability both when administered individually and when combined with other treatments (Figure 3A). Notably, the viability of colon cancer cells exhibited a decreasing trend as the proportion of cocultured Jurkat T cells increased. Cell viability data were presented in the form of a bar graph (Figure 3B). To obtain further insight, we also conducted assessments of IL-2 levels in the culture media of coculture experiments. Encouragingly, we also observed an elevated concentration of this immune-related factor during cotreatment with COS and IFN-γ (Figure 3C). These collective findings provide compelling evidence that COS plays a significant role in enhancing T-cell-mediated cell death by suppressing PD-L1 expression, diminishing the immunosuppressive capacity of the PD-1/PD-L1 axis.
3.3. COS Induces Apoptosis by Downregulating AKT and ERK Signaling in Colon Cancer Cells
The data shown in Figure 2 confirm the reduction in PD-L1 expression following COS treatment. Subsequently, we investigated the underlying pathway that is responsible for this effect. Following the established knowledge, which suggests that the inhibition of the PI3K–AKT–mTOR pathway leads to decreased PD-L1 expression and instigates PD-L1 upregulation via the ERK signaling pathway, we evaluated the expression levels of AKT and ERK [27,28]. Our data indicate that COS (0, 1.25, 2.5, 5 µM) treatment dose-dependently decreases p-AKT and p-ERK in colon cancer cells (Figure 4A). In accordance with the observation that ERK and AKT signaling pathways are downregulated in colon cancer cells to facilitate apoptosis, our study also assessed the apoptosis signaling pathway [29]. The expression patterns of apoptosis-related proteins, including PARP, Bcl-2, and BAX, demonstrate that COS (0, 1.25, 2.5, 5 µM) treatment leads to the induction of apoptosis (Figure 4B). Then, the occurrence of apoptosis was measured by FACS. It was confirmed through Annexin V–FITC/PI staining that the apoptosis extent increased in a concentration-dependent manner with COS (0, 1.25, 2.5, 5 µM) (Figure 4C,D). Taken together, these results show that COS induces apoptosis through the regulation of AKT and ERK signaling.
3.4. COS Treatment Increases ROS Levels in DLD-1, Hct116, and RKO Cells
Several studies have investigated the relationship between reactive oxygen species (ROS) and PD-L1 expression [21]. Among the drugs producing ROS in cancer cells, some have been shown to up- or downregulate PD-L1 [21]. The role of ROS in triggering apoptosis is widely acknowledged in various publications [30]. Because we observed apoptosis resulting from COS treatment in our prior experiment, we formulated the hypothesis that COS treatment induces apoptosis by generating ROS. Consequently, we quantified ROS levels. Measurements using fluorescence and flow cytometry showed a concentration-dependent increase in ROS levels in DLD-1, Hct116, and RKO cells following COS (0, 1.25, 2.5, 5 µM) treatment, which was reversed by NAC treatment (Figure 5A). Fluorescence values were represented in a bar graph (Figure 5B).
4. Discussion
In recent years, ICIs have emerged as a part of groundbreaking therapies in oncology, revolutionizing the treatment landscape for a range of cancers [31]. The regulation of PD-L1 expression has immense significance for ICI therapy [32]. For maximizing the efficacy of ICIs and optimizing patient outcomes, understanding the intricate mechanisms that govern PD-L1 expression is pivotal [33]. PD-L1 expression levels in tumor tissues have been linked with patient responsiveness to ICIs, making it a predictive biomarker for treatment success [16]. Tumors with higher PD-L1 expression tend to exhibit a greater immunosuppressive microenvironment, rendering them more susceptible to an immune checkpoint blockade [34]. At present, extensive research efforts are being directed toward unraveling the complex regulatory pathways that control PD-L1 expression [35]. Multiple factors influence PD-L1 expression, including oncogenic signaling pathways, inflammatory cytokines, genetic alterations, and epigenetic modifications [36]. For instance, oncogenic signaling pathways, including the MAPK and PI3K pathways, can upregulate PD-L1 expression in response to cellular stress and inflammation [37]. In addition, tumor-infiltrating immune cells, such as interferon-gamma-producing T cells, can induce PD-L1 expression in cancer cells through a positive feedback loop [38]. Efforts are also underway to develop strategies of modulating PD-L1 expression and enhancing the effectiveness of ICIs [39]. Combination therapies targeting both PD-L1 expression and other immune checkpoints or signaling pathways are being explored to overcome resistance mechanisms and broaden patient populations that can benefit from ICIs [40]. Additionally, understanding the role of PD-L1 expression dynamics in disease progression and treatment response is crucial for tailoring therapeutic approaches on an individualized basis.
This study investigated the use of COS as a regulator of PD-L1 expression in CRC, a novel and promising avenue within cancer immunotherapy. This investigation aligns with an increasing interest in natural compounds as potential modulators of immune responses, harnessing their diverse bioactive properties for therapeutic benefit [41,42]. The investigation into the effects of COS treatment on CRC cell lines provided two crucial findings, namely, the inhibition of PD-L1 expression and the induction of apoptosis through ROS-related pathways. The observed inhibition of PD-L1 expression for COS treatment aligns with the growing interest in natural compounds as potential PD-L1 modulators [43,44]. COS’s ability to downregulate PD-L1 expression has implications for the disruption of the immune evasion mechanisms employed by cancer cells, potentially enhancing the effectiveness of approaches using immunotherapy. Furthermore, the induction of apoptosis through ROS-related pathways forms a significant mechanistic insight [45]. The connection between apoptosis and immune checkpoint regulation opens an intriguing perspective [46]. ROS-mediated apoptosis not only affects cancer cell survival but also has potential implications for modulating the immunogenicity of the tumor microenvironment [47,48]. This study’s contribution to elucidating the interplay among ROS, apoptosis, and PD-L1 expression contains valuable insights into the potential synergistic mechanisms underlying cancer immunotherapy. Further investigation is warranted to validate the observed effects across a broader range of CRC models and to more comprehensively delineate the molecular pathways that underlie COS’s actions. Additionally, the assessment of COS’s safety profile and its potential interactions with other therapeutic agents will be critical steps for translating these findings into clinical applications.
Previous studies on Salvia plebeia in our laboratory have shown that COS possesses immunotherapeutic properties, including its demonstration of the most potent PD-1/PD-L1 blockade, functioning as an ICI [10]. This study further substantiates the finding that COS directly inhibits PD-L1 expression in colon cancer cell lines, producing an anticancer effect. Because in vivo experiments were carried out in preceding work, additional in vivo experiments were not performed here. However, future research, especially involving the exploration of novel treatment strategies, including combination treatments, necessitates the implementation of new in vivo tests.
Previous studies have established that the AKT and ERK signaling pathways contribute to the upregulation of PD-L1 expression in non-small cell lung cancer, multiple myeloma, and triple-negative breast cancer [27,49,50,51]. On the other hand, reports have indicated that PD-L1 can activate MEK/ERK and PI3K/AKT signaling pathways within CRC cells [52]. Our findings indicated a reduction in the levels of phosphorylated AKT and ERK (p-AKT, p-ERK) in CRC cells following treatment with COS, as depicted in Figure 4A. Because we used colon cancer cells in this paper, it may be that COS treatment downregulates AKT and ERK signaling through the regulation of PD-L1 expression, as is described in previous publications on CRC. IFN-γ regulates the transcription of PD-L1, leading to an increase in its expression [53,54]. However, ROS suppresses PD-L1 expression via the inhibition of fibroblast growth factor receptor 1 expression, the inhibition of the phosphorylation of eukaryotic translation initiation factor 4E-binding protein 1, and the activation of eukaryotic initiation factor 2α [22,55]. Consequently, Figure 2C demonstrates that although there is an increase in PD-L1 expression induced by IFN-γ, PD-L1 expression decreases due to elevated ROS resulting from COS treatment. However, the mechanism by which PD-L1 functions within CRCs remains unclear. ERK and AKT signaling pathways, as well as ROS and IFN-γ, involved in PD-L1 expression, require further investigation.
Several studies have indicated that drug treatments can result in increased ROS levels, leading to a decrease in PD-L1 expression. For instance, Ethaselen, an organoselenium compound, inhibits TrxR1, a redox sensor and antioxidant enzyme, which results in elevated ROS levels within cells and subsequently reduces the expression of PD-L1 [56,57]. Butaselen, a TrxR1 inhibitor and ROS generator, suppresses PD-L1 expression on the tumor cell surface through the STAT3 pathway [58,59]. Metformin, a biguanide drug commonly employed in diabetes treatment, has anticancer effects both in vitro and in vivo [60]. It prompts cancer cell apoptosis through oxidative stress and ROS production [60,61]. Additionally, metformin reduces PD-L1 expression by modulating the Hippo signaling pathway [62]. Chaetocin, a potent inhibitor of TrxR1 and histone methyltransferase, triggers apoptosis in cancer cells by inducing excessive accumulations of ROS [63]. Notably, chaetocin treatment markedly diminishes PD-L1 protein levels in human pancreatic cancer cells [64]. On the other hand, numerous studies have demonstrated that drug usage can elevate ROS levels, resulting in augmented PD-L1 expression [65,66,67,68]. However, our findings align with those of other studies that have indicated a correlation between increased ROS levels and subsequent reductions in PD-L1 expression.
This study suggests that COS may have promising potential in conjunction with other cancer therapies, specifically ICIs. By inhibiting the expression of PD-L1, COS can sensitize cancer cells, making them more susceptible to the effects of ICIs. This synergistic approach can enhance treatment outcomes for cancer patients, although it is crucial to emphasize that further studies and clinical trials are imperative for substantiating this hypothesis [69]. These findings indicate the importance of exploring innovative combination therapies for augmenting the efficacy of cancer treatment and laying the groundwork for more effective strategies against cancer. However, it is important to acknowledge a primary limitation of our study, namely, the relatively small number of CRC cell lines utilized in testing COS. To validate our findings, it is imperative to conduct additional research that encompasses a broader spectrum of types of cancer. Furthermore, comprehensive investigations are required to elucidate the optimal COS treatment methods in the contexts of various cancers.
5. Conclusions
This study demonstrates the potential of COS as an ICI in CRC treatment. Through regulating PD-L1 expression and initiating apoptosis via ROS-related pathways, COS is a naturally occurring compound with versatile effects that can bolster the effectiveness of immunotherapies. Further experimental and clinical validation is imperative for fully harnessing the therapeutic promise of COS to enhance patient outcomes in CRC and potentially other types of malignancies (Figure 6).
Author Contributions
Conceptualization, J.H.H., E.-J.L., J.-G.C. and H.-S.C.; validation and investigation, J.H.H., E.-J.L. and W.P.; resources, E.-J.L. and J.-G.C.; writing—original draft preparation, J.H.H.; writing—review and editing, J.H.H., E.-J.L., W.P., J.-G.C. and K.-T.H.; visualization, J.H.H., E.-J.L., W.P. and K.-T.H.; supervision, H.-S.C.; funding acquisition, H.-S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MIST) (No. NRF-2022R1A2C2092834) and also by the Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT) (No. RS-2023-00261934).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The information will be provided upon sensible inquiry.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Zhai, Z.; Yu, X.; Yang, B.; Zhang, Y.; Zhang, L.; Li, X.; Sun, H. Colorectal cancer heterogeneity and targeted therapy: Clinical implications, challenges and solutions for treatment resistance. Semin. Cell Dev. Biol. 2017, 64, 107–115. [Google Scholar] [CrossRef]
- Havel, J.J.; Chowell, D.; Chan, T.A. The evolving landscape of biomarkers for checkpoint inhibitor immunotherapy. Nat. Rev. Cancer 2019, 19, 133–150. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 10. [Google Scholar] [CrossRef]
- Kim, J.; Chen, D.S. Immune escape to PD-L1/PD-1 blockade: Seven steps to success (or failure). Ann. Oncol. 2016, 27, 1492–1504. [Google Scholar] [CrossRef]
- Masugi, Y.; Nishihara, R.; Yang, J.; Mima, K.; Da Silva, A.; Shi, Y.; Inamura, K.; Cao, Y.; Song, M.; Nowak, J.A. Tumour CD274 (PD-L1) expression and T cells in colorectal cancer. Gut 2017, 66, 1463–1473. [Google Scholar] [CrossRef]
- Kim, S.M.; Vetrivel, P.; Ha, S.E.; Kim, H.H.; Kim, J.-A.; Kim, G.S. Apigetrin induces extrinsic apoptosis, autophagy and G2/M phase cell cycle arrest through PI3K/AKT/mTOR pathway in AGS human gastric cancer cell. J. Nutr. Biochem. 2020, 83, 108427. [Google Scholar] [CrossRef]
- Lakshmi, M.; Swapna, T. A computational study on Cosmosiin, an antiviral compound from Memecylon randerianum SM Almeida & MR Almeida. Med. Plants-Int. J. Phytomed. Relat. Ind. 2021, 13, 515–523. [Google Scholar]
- Rao, Y.K.; Lee, M.-J.; Chen, K.; Lee, Y.-C.; Wu, W.-S.; Tzeng, Y.-M. Insulin-mimetic action of rhoifolin and cosmosiin isolated from Citrus grandis (L.) Osbeck leaves: Enhanced adiponectin secretion and insulin receptor phosphorylation in 3T3-L1 cells. Evid.-Based Complement. Altern. Med. 2011, 2011, 624375. [Google Scholar] [CrossRef]
- Choi, J.-G.; Kim, Y.S.; Kim, J.H.; Kim, T.I.; Li, W.; Oh, T.W.; Jeon, C.H.; Kim, S.J.; Chung, H.-S. Anticancer effect of Salvia plebeia and its active compound by improving T-cell activity via blockade of PD-1/PD-L1 interaction in humanized PD-1 mouse model. Front. Immunol. 2020, 11, 598556. [Google Scholar] [CrossRef]
- Min, Z.; Tang, Y.; Hu, X.-T.; Zhu, B.-L.; Ma, Y.-L.; Zha, J.-S.; Deng, X.-J.; Yan, Z.; Chen, G.-J. Cosmosiin increases ADAM10 expression via mechanisms involving 5’UTR and PI3K signaling. Front. Mol. Neurosci. 2018, 11, 198. [Google Scholar] [CrossRef] [PubMed]
- Mikhaeil, B.R.; Badria, F.A.; Maatooq, G.T.; Amer, M.M. Antioxidant and immunomodulatory constituents of henna leaves. Z. Für Naturforschung C 2004, 59, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-G.; Rosalen, P.; Falsetta, M.; Koo, H. Natural products in caries research: Current (limited) knowledge, challenges and future perspective. Caries Res. 2011, 45, 243–263. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [PubMed]
- Coriano, C.G.; Liu, F.; Sievers, C.K.; Liang, M.; Wang, Y.; Lim, Y.; Yu, M.; Xu, W. A computational-based approach to identify estrogen receptor α/β heterodimer selective ligands. Mol. Pharmacol. 2018, 93, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.P.; Kurzrock, R. PD-L1 expression as a predictive biomarker in cancer immunotherapy. Mol. Cancer Ther. 2015, 14, 847–856. [Google Scholar] [CrossRef]
- Deng, L.-J.; Qi, M.; Li, N.; Lei, Y.-H.; Zhang, D.-M.; Chen, J.-X. Natural products and their derivatives: Promising modulators of tumor immunotherapy. J. Leucoc. Biol. 2020, 108, 493–508. [Google Scholar] [CrossRef]
- Lee, J.; Han, Y.; Wang, W.; Jo, H.; Kim, H.; Kim, S.; Yang, K.-M.; Kim, S.-J.; Dhanasekaran, D.N.; Song, Y.S. Phytochemicals in cancer immune checkpoint inhibitor therapy. Biomolecules 2021, 11, 1107. [Google Scholar] [CrossRef]
- Kloten, V.; Lampignano, R.; Krahn, T.; Schlange, T. Circulating tumor cell PD-L1 expression as biomarker for therapeutic efficacy of immune checkpoint inhibition in NSCLC. Cells 2019, 8, 809. [Google Scholar] [CrossRef]
- Darvin, P.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Immune checkpoint inhibitors: Recent progress and potential biomarkers. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef]
- Bailly, C. Regulation of PD-L1 expression on cancer cells with ROS-modulating drugs. Life Sci. 2020, 246, 117403. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, C.; Xia, X.; He, Y.-Q.; Hu, Y.; Cremer, K.; Robert, A.; Liu, J.; Wang, F.; Ling, J.; Chiao, P.J. Regulation of PD-L1 expression in K-ras-driven cancers through ROS-mediated FGFR1 signaling. Redox Biol. 2021, 38, 101780. [Google Scholar] [CrossRef] [PubMed]
- Adeshakin, A.O.; Liu, W.; Adeshakin, F.O.; Afolabi, L.O.; Zhang, M.; Zhang, G.; Wang, L.; Li, Z.; Lin, L.; Cao, Q. Regulation of ROS in myeloid-derived suppressor cells through targeting fatty acid transport protein 2 enhanced anti-PD-L1 tumor immunotherapy. Cell. Immunol. 2021, 362, 104286. [Google Scholar] [CrossRef] [PubMed]
- Toda, K.; Kawada, K.; Iwamoto, M.; Inamoto, S.; Sasazuki, T.; Shirasawa, S.; Hasegawa, S.; Sakai, Y. Metabolic alterations caused by KRAS mutations in colorectal cancer contribute to cell adaptation to glutamine depletion by upregulation of asparagine synthetase. Neoplasia 2016, 18, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, D.; Eide, P.; Eilertsen, I.; Danielsen, S.; Eknæs, M.; Hektoen, M.; Lind, G.; Lothe, R. Epigenetic and genetic features of 24 colon cancer cell lines. Oncogenesis 2013, 2, e71. [Google Scholar] [CrossRef]
- Whittaker, S.R.; Cowley, G.S.; Wagner, S.; Luo, F.; Root, D.E.; Garraway, L.A. Combined Pan-RAF and MEK inhibition overcomes multiple resistance mechanisms to selective RAF inhibitors. Mol. Cancer Ther. 2015, 14, 2700–2711. [Google Scholar] [CrossRef] [PubMed]
- Lastwika, K.J.; Wilson III, W.; Li, Q.K.; Norris, J.; Xu, H.; Ghazarian, S.R.; Kitagawa, H.; Kawabata, S.; Taube, J.M.; Yao, S. Control of PD-L1 expression by oncogenic activation of the AKT–mTOR pathway in non–small cell lung cancer. Cancer Res. 2016, 76, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Li, P.; Zhang, C.; Zhao, Y.; Hu, H.; Wen, G. The TLR4/ERK/PD-L1 axis may contribute to NSCLC initiation. Int. J. Oncol. 2020, 57, 456–465. [Google Scholar] [CrossRef]
- Liao, C.H.; Sang, S.; Ho, C.T.; Lin, J.K. Garcinol modulates tyrosine phosphorylation of FAK and subsequently induces apoptosis through down-regulation of Src, ERK, and Akt survival signaling in human colon cancer cells. J. Cell. Biochem. 2005, 96, 155–169. [Google Scholar] [CrossRef]
- Simon, H.-U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Kraehenbuehl, L.; Weng, C.-H.; Eghbali, S.; Wolchok, J.D.; Merghoub, T. Enhancing immunotherapy in cancer by targeting emerging immunomodulatory pathways. Nat. Rev. Clin. Oncol. 2022, 19, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.A.; Patel, V.G. The role of PD-L1 expression as a predictive biomarker: An analysis of all US Food and Drug Administration (FDA) approvals of immune checkpoint inhibitors. J. Immunother. Cancer 2019, 7, 278. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, A.J.; Rizvi, H.; Bandlamudi, C.; Sauter, J.L.; Travis, W.D.; Rekhtman, N.; Plodkowski, A.J.; Perez-Johnston, R.; Sawan, P.; Beras, A. Clinical and molecular correlates of PD-L1 expression in patients with lung adenocarcinomas. Ann. Oncol. 2020, 31, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Fang, Y.C.; Li, J. PD-L1 expression levels on tumor cells affect their immunosuppressive activity. Oncol. Lett. 2019, 18, 5399–5407. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.-H.; Chan, L.-C.; Li, C.-W.; Hsu, J.L.; Hung, M.-C. Mechanisms controlling PD-L1 expression in cancer. Mol. Cell 2019, 76, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, X. Study and analysis of antitumor resistance mechanism of PD1/PD-L1 immune checkpoint blocker. Cancer Med. 2020, 9, 8086–8121. [Google Scholar] [CrossRef] [PubMed]
- Ritprajak, P.; Azuma, M. Intrinsic and extrinsic control of expression of the immunoregulatory molecule PD-L1 in epithelial cells and squamous cell carcinoma. Oral Oncol. 2015, 51, 221–228. [Google Scholar] [CrossRef]
- Derer, A.; Spiljar, M.; Bäumler, M.; Hecht, M.; Fietkau, R.; Frey, B.; Gaipl, U.S. Chemoradiation increases PD-L1 expression in certain melanoma and glioblastoma cells. Front. Immunol. 2016, 7, 610. [Google Scholar] [CrossRef]
- Song, C.-H.; Kim, N.; Nam, R.H.; Choi, S.I.; Jang, J.Y.; Kim, J.W.; Na, H.Y.; Lee, H.-N. Combination treatment with 17β-estradiol and anti-PD-L1 suppresses MC38 tumor growth by reducing PD-L1 expression and enhancing M1 macrophage population in MC38 colon tumor model. Cancer Lett. 2022, 543, 215780. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Hsu, J.-M.; Yang, W.-H.; Hung, M.-C. Mechanisms regulating PD-L1 expression in cancers and associated opportunities for novel small-molecule therapeutics. Nat. Rev. Clin. Oncol. 2022, 19, 287–305. [Google Scholar] [CrossRef]
- Dias, A.S.; Helguero, L.; Almeida, C.R.; Duarte, I.F. Natural compounds as metabolic modulators of the tumor microenvironment. Molecules 2021, 26, 3494. [Google Scholar] [CrossRef] [PubMed]
- da Silva Meirelles, L.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhong, A.; Wang, Q.; Zheng, T. Structure-based pharmacophore modeling, virtual screening, molecular docking, ADMET, and molecular dynamics (MD) simulation of potential inhibitors of PD-L1 from the library of marine natural products. Mar. Drugs 2021, 20, 29. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Ding, J.; Cui, X.; Wu, M.; Huang, C.; Zhang, R.; Wang, J.; Li, X.; Cen, S.; Zhou, J. Screening of kinase inhibitors downregulating PD-L1 expression via on/in cell quantitative immunoblots. Eur. J. Pharm. Sci. 2020, 142, 105088. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ragheb, K.; Lawler, G.; Sturgis, J.; Rajwa, B.; Melendez, J.A.; Robinson, J.P. Mitochondrial complex I inhibitor rotenone induces apoptosis through enhancing mitochondrial reactive oxygen species production. J. Biol. Chem. 2003, 278, 8516–8525. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K.; Najafi, M. Immune system in cancer radiotherapy: Resistance mechanisms and therapy perspectives. Crit. Rev. Oncol. /Hematol. 2021, 157, 103180. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Yang, W.; Zhou, Z.; Tian, R.; Lin, L.; Ma, Y.; Song, J.; Chen, X. Targeted scavenging of extracellular ROS relieves suppressive immunogenic cell death. Nat. Commun. 2020, 11, 4951. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Tani, K. Multimodal immunogenic cancer cell death as a consequence of anticancer cytotoxic treatments. Cell Death Differ. 2014, 21, 39–49. [Google Scholar] [CrossRef]
- Liu, J.; Hamrouni, A.; Wolowiec, D.; Coiteux, V.; Kuliczkowski, K.; Hetuin, D.; Saudemont, A.; Quesnel, B. Plasma cells from multiple myeloma patients express B7-H1 (PD-L1) and increase expression after stimulation with IFN-γ and TLR ligands via a MyD88-, TRAF6-, and MEK-dependent pathway. Blood J. Am. Soc. Hematol. 2007, 110, 296–304. [Google Scholar] [CrossRef]
- Loi, S.; Dushyanthen, S.; Beavis, P.A.; Salgado, R.; Denkert, C.; Savas, P.; Combs, S.; Rimm, D.L.; Giltnane, J.M.; Estrada, M.V. RAS/MAPK activation is associated with reduced tumor-infiltrating lymphocytes in triple-negative breast cancer: Therapeutic cooperation between MEK and PD-1/PD-L1 immune checkpoint inhibitors. Clin. Cancer Res. 2016, 22, 1499–1509. [Google Scholar] [CrossRef]
- Zerdes, I.; Matikas, A.; Bergh, J.; Rassidakis, G.Z.; Foukakis, T. Genetic, transcriptional and post-translational regulation of the programmed death protein ligand 1 in cancer: Biology and clinical correlations. Oncogene 2018, 37, 4639–4661. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Zhang, T.; Deng, S.-C.; Wei, J.-C.; Yang, P.; Wang, Q.; Chen, Z.-P.; Li, W.-L.; Chen, H.-C.; Hu, H. PD-L1 promotes colorectal cancer stem cell expansion by activating HMGA1-dependent signaling pathways. Cancer Lett. 2019, 450, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Tang, X.; Wong, S.W.; Guo, A.; Lin, Y.; Kwok, H.F. Regulation of IFN-γ-mediated PD-L1 expression by MYC in colorectal cancer with wild-type KRAS and TP53 and its clinical implications. Front. Pharmacol. 2022, 13, 1022129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Li, Y.; Zhang, J.; Zhang, B. PD-L1 expression increased by IFN-γ via JAK2-STAT1 signaling and predicts a poor survival in colorectal cancer. Oncol. Lett. 2020, 20, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.-W.; Jiang, X.-M.; Xu, Y.-L.; Huang, M.-Y.; Chen, Y.-C.; Yu, W.-B.; Su, M.-X.; Ye, Z.-H.; Chen, X.; Wang, Y. Licochalcone A inhibits interferon-gamma-induced programmed death-ligand 1 in lung cancer cells. Phytomedicine 2021, 80, 153394. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, Z.; Fu, J.; Yin, H.; Xiong, K.; Tan, Q.; Jin, H.; Li, J.; Wang, T.; Tang, W. Ethaselen: A potent mammalian thioredoxin reductase 1 inhibitor and novel organoselenium anticancer agent. Free Radic. Biol. Med. 2012, 52, 898–908. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, Y.; Bai, M.; Liu, Y.; Xu, B.; Sun, R.; Zeng, H. The antimetastatic effect and underlying mechanisms of thioredoxin reductase inhibitor ethaselen. Free Radic. Biol. Med. 2019, 131, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ma, W.; Sun, R.; Yin, H.; Lin, F.; Liu, Y.; Xu, W.; Zeng, H. Butaselen prevents hepatocarcinogenesis and progression through inhibiting thioredoxin reductase activity. Redox Biol. 2018, 14, 237–249. [Google Scholar] [CrossRef]
- Zou, Q.; Chen, Y.-F.; Zheng, X.-Q.; Ye, S.-F.; Xu, B.-Y.; Liu, Y.-X.; Zeng, H.-H. Novel thioredoxin reductase inhibitor butaselen inhibits tumorigenesis by down-regulating programmed death-ligand 1 expression. J. Zhejiang Univ. Science. B 2018, 19, 689. [Google Scholar] [CrossRef]
- Park, D. Metformin induces oxidative stress-mediated apoptosis without the blockade of glycolysis in H4IIE hepatocellular carcinoma cells. Biol. Pharm. Bull. 2019, 42, 2002–2008. [Google Scholar] [CrossRef]
- Park, W.; Wei, S.; Kim, B.-S.; Kim, B.; Bae, S.-J.; Chae, Y.C.; Ryu, D.; Ha, K.-T. Diversity and complexity of cell death: A historical review. Exp. Mol. Med. 2023, 55, 573–1594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-J.; Zhang, Q.-S.; Li, Z.-Q.; Zhou, J.-W.; Du, J. Metformin attenuates PD-L1 expression through activating Hippo signaling pathway in colorectal cancer cells. Am. J. Transl. Res. 2019, 11, 6965. [Google Scholar] [PubMed]
- Wen, C.; Wang, H.; Wu, X.; He, L.; Zhou, Q.; Wang, F.; Chen, S.; Huang, L.; Chen, J.; Wang, H. ROS-mediated inactivation of the PI3K/AKT pathway is involved in the antigastric cancer effects of thioredoxin reductase-1 inhibitor chaetocin. Cell Death Dis. 2019, 10, 809. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Paschall, A.V.; Shi, H.; Savage, N.; Waller, J.L.; Sabbatini, M.E.; Oberlies, N.H.; Pearce, C.; Liu, K. The MLL1-H3K4me3 axis-mediated PD-L1 expression and pancreatic cancer immune evasion. JNCI J. Natl. Cancer Inst. 2017, 109, djw283. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Guo, L.; Zhang, B.; Liu, S.; Zhang, K.; Yan, J.; Zhang, W.; Yu, M.; Chen, Z.; Xu, Y. Disulfiram combined with copper induces immunosuppression via PD-L1 stabilization in hepatocellular carcinoma. Am. J. Cancer Res. 2019, 9, 2442. [Google Scholar] [PubMed]
- Xia, Y.; Jia, C.; Xue, Q.; Jiang, J.; Xie, Y.; Wang, R.; Ran, Z.; Xu, F.; Zhang, Y.; Ye, T. Antipsychotic drug trifluoperazine suppresses colorectal cancer by inducing G0/G1 arrest and apoptosis. Front. Pharmacol. 2019, 10, 1029. [Google Scholar] [CrossRef]
- Wang, X.; Li, J.; Dong, K.; Lin, F.; Long, M.; Ouyang, Y.; Wei, J.; Chen, X.; Weng, Y.; He, T. Tumor suppressor miR-34a targets PD-L1 and functions as a potential immunotherapeutic target in acute myeloid leukemia. Cell. Signal. 2015, 27, 443–452. [Google Scholar] [CrossRef]
- Raninga, P.V.; Lee, A.C.; Sinha, D.; Shih, Y.Y.; Mittal, D.; Makhale, A.; Bain, A.L.; Nanayakarra, D.; Tonissen, K.F.; Kalimutho, M. Therapeutic cooperation between auranofin, a thioredoxin reductase inhibitor and anti-PD-L1 antibody for treatment of triple-negative breast cancer. Int. J. Cancer 2020, 146, 123–136. [Google Scholar] [CrossRef]
- Naimi, A.; Mohammed, R.N.; Raji, A.; Chupradit, S.; Yumashev, A.V.; Suksatan, W.; Shalaby, M.N.; Thangavelu, L.; Kamrava, S.; Shomali, N. Tumor immunotherapies by immune checkpoint inhibitors (ICIs); the pros and cons. Cell Commun. Signal. 2022, 20, 44. [Google Scholar] [CrossRef]
Figure 1.
Cell viability, measured by COS and IFN-γ treatment in colon cancer cell lines. (A) Structural formula of COS. (B,C) Cell viability was assessed for DLD-1, Hct116, and RKO cells following exposure to COS and IFN-γ at the specified doses over a period of 2 days. The viabilities of three cells were analyzed by CCK. The results are shown as the means and SEMs. *** p < 0.001 and **** p < 0.0001 relative to the respective controls. The experiments were conducted independently three times.
Figure 1.
Cell viability, measured by COS and IFN-γ treatment in colon cancer cell lines. (A) Structural formula of COS. (B,C) Cell viability was assessed for DLD-1, Hct116, and RKO cells following exposure to COS and IFN-γ at the specified doses over a period of 2 days. The viabilities of three cells were analyzed by CCK. The results are shown as the means and SEMs. *** p < 0.001 and **** p < 0.0001 relative to the respective controls. The experiments were conducted independently three times.
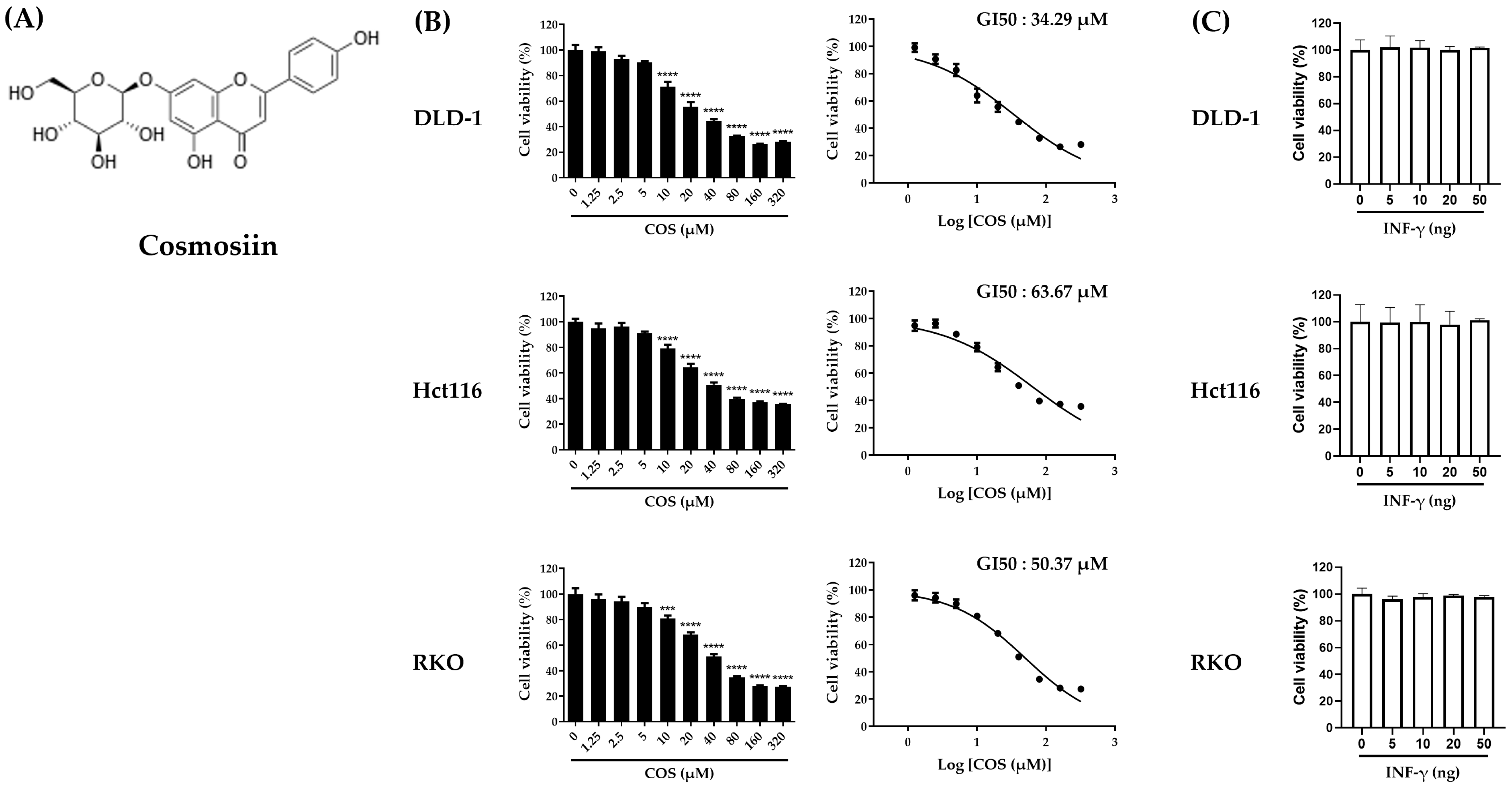
Figure 2.
COS suppresses the PD-L1 expression in a colon cancer cell line. (A–D) The PD-L1 expression of DLD-1, Hct116, and RKO cells was measured following treatment with a COS and IFN-γ indicated dose for 2 days. Experiments (A–C) and (D) were performed with a Western blot assay and FACS, respectively. The results are shown as the means and SEMs. ** p < 0.01, *** p < 0.001, and **** p < 0.0001 relative to the respective controls. The experiments were conducted independently three times.
Figure 2.
COS suppresses the PD-L1 expression in a colon cancer cell line. (A–D) The PD-L1 expression of DLD-1, Hct116, and RKO cells was measured following treatment with a COS and IFN-γ indicated dose for 2 days. Experiments (A–C) and (D) were performed with a Western blot assay and FACS, respectively. The results are shown as the means and SEMs. ** p < 0.01, *** p < 0.001, and **** p < 0.0001 relative to the respective controls. The experiments were conducted independently three times.
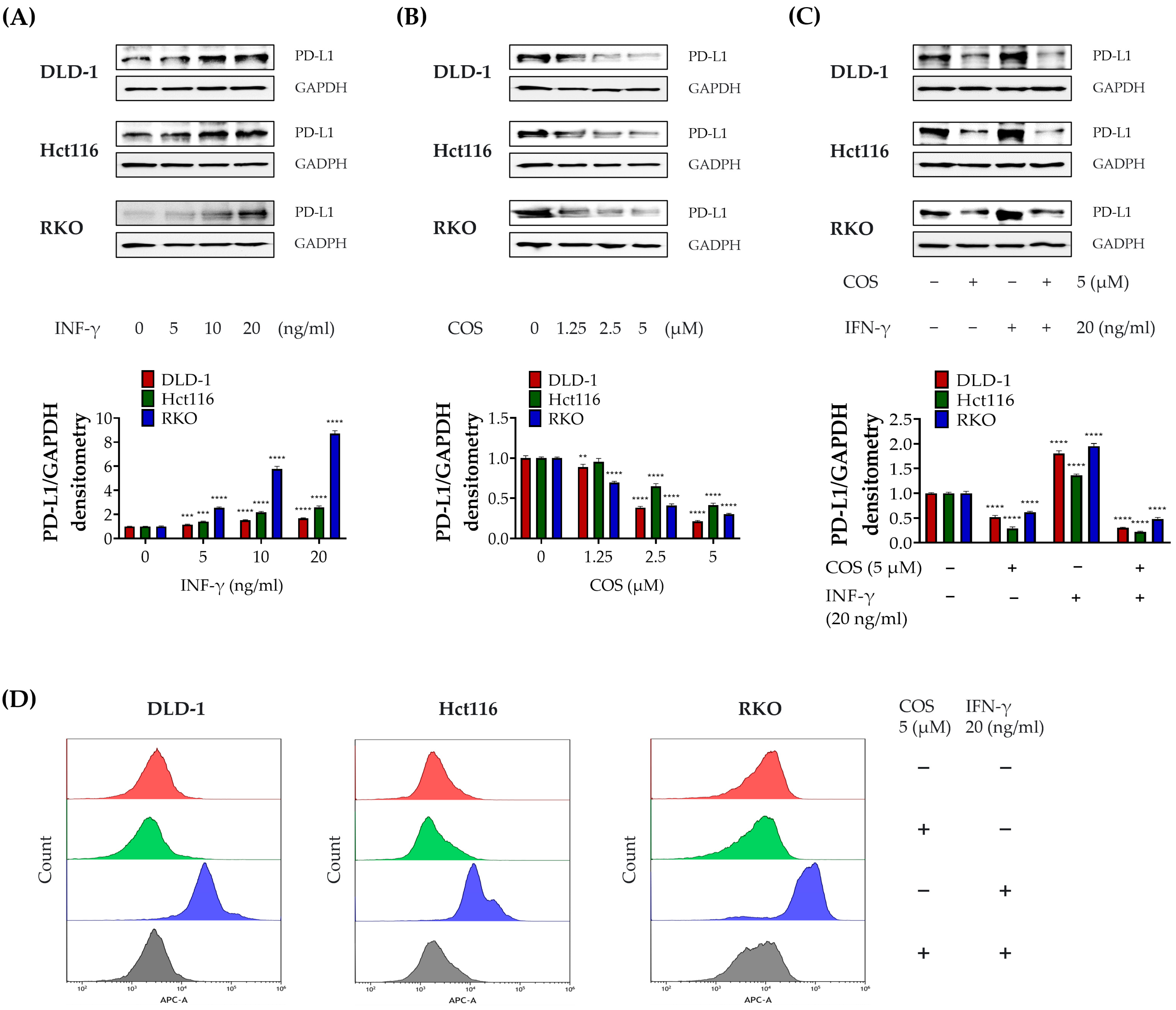
Figure 3.
COS augmentation of T-cell-mediated cell death and immune mediator release in a colon cancer cell line. (A,B) Coculture experiments were conducted in seeded colon cancer cell lines with added COS (5 μM), IFN-γ (20 ng), and Jurkat T cells (1:10, 1:20) for 24 h. After incubation, the cancer cells were stained with crystal violet, and the intensity of live cells was measured and bar graphed. (C) The level of IL-2 in the coculture medium was estimated using an ELISA kit. The viabilities and cytokines were examined individually, using CCK and ELISA. The results are shown as the means and SEMs. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, and ns (not significant) compared to their respective controls. The experiments were conducted three times independently.
Figure 3.
COS augmentation of T-cell-mediated cell death and immune mediator release in a colon cancer cell line. (A,B) Coculture experiments were conducted in seeded colon cancer cell lines with added COS (5 μM), IFN-γ (20 ng), and Jurkat T cells (1:10, 1:20) for 24 h. After incubation, the cancer cells were stained with crystal violet, and the intensity of live cells was measured and bar graphed. (C) The level of IL-2 in the coculture medium was estimated using an ELISA kit. The viabilities and cytokines were examined individually, using CCK and ELISA. The results are shown as the means and SEMs. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, and ns (not significant) compared to their respective controls. The experiments were conducted three times independently.
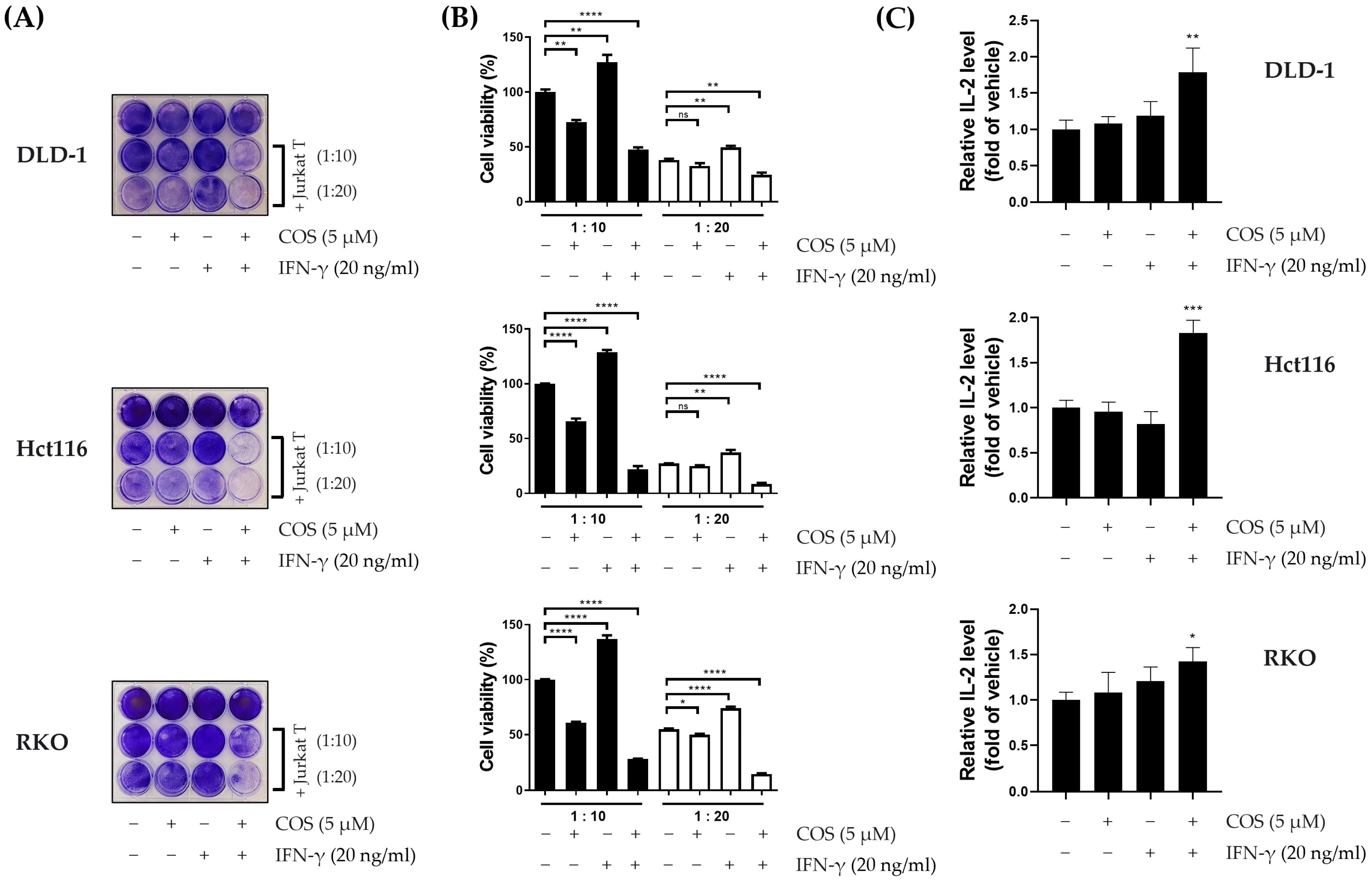
Figure 4.
Measurement of apoptosis-related protein and the ratio of apoptotic cells in a colon cancer cell line by COS. (A,B) Protein expression levels for p-AKT, AKT, p-ERK, ERK, PARP, cleaved-PARP, Bcl-2, BAX, and GAPDH were assessed using Western blot analysis. (C) The quantification of apoptotic cells was conducted through FACS analysis, employing PI-Annexin V staining. (D) The bar chart represents the cells of the ratio of apoptosis. DLD-1, Hct116, and RKO cells were treated with COS (0, 1.25, 2.5, or 5 μM) for a duration of 2 days. The results are shown as the means and SEMs. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001 compared to their respective controls. The experiments were conducted three times independently.
Figure 4.
Measurement of apoptosis-related protein and the ratio of apoptotic cells in a colon cancer cell line by COS. (A,B) Protein expression levels for p-AKT, AKT, p-ERK, ERK, PARP, cleaved-PARP, Bcl-2, BAX, and GAPDH were assessed using Western blot analysis. (C) The quantification of apoptotic cells was conducted through FACS analysis, employing PI-Annexin V staining. (D) The bar chart represents the cells of the ratio of apoptosis. DLD-1, Hct116, and RKO cells were treated with COS (0, 1.25, 2.5, or 5 μM) for a duration of 2 days. The results are shown as the means and SEMs. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001 compared to their respective controls. The experiments were conducted three times independently.
Figure 5.
Increased production of ROS in DLD-1, Hct116, and RKO cells by COS treatment. (A,B) DLD-1, Hct116, and RKO cells were treated with the indicated COS and NAC for 2 days, and ROS was measured using FACS. The bar graphs on the right present the fluorescence values and FITC mean values, respectively. * p < 0.05, ** p < 0.01, and **** p < 0.0001 compared to their respective controls. The experiments were conducted three times independently.
Figure 5.
Increased production of ROS in DLD-1, Hct116, and RKO cells by COS treatment. (A,B) DLD-1, Hct116, and RKO cells were treated with the indicated COS and NAC for 2 days, and ROS was measured using FACS. The bar graphs on the right present the fluorescence values and FITC mean values, respectively. * p < 0.05, ** p < 0.01, and **** p < 0.0001 compared to their respective controls. The experiments were conducted three times independently.

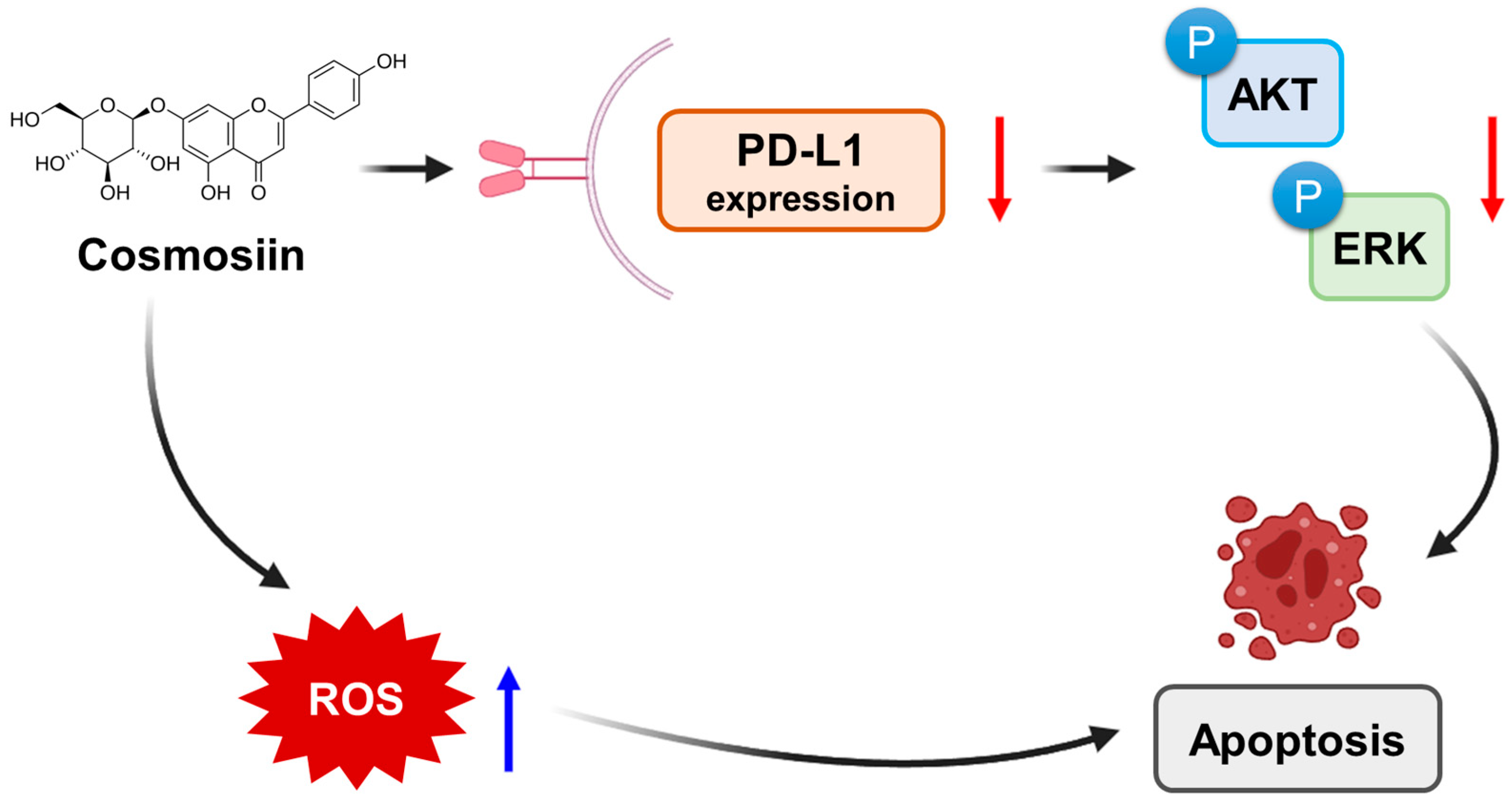
Figure 6.
Schematic representation of the work on COS. COS inhibits the phosphorylation of AKT and ERK, and this inhibition leads to a decrease in the expression of PD-L1. Additionally, COS increases the production of ROS. These effects lead to the apoptosis of cancer cells.
Figure 6.
Schematic representation of the work on COS. COS inhibits the phosphorylation of AKT and ERK, and this inhibition leads to a decrease in the expression of PD-L1. Additionally, COS increases the production of ROS. These effects lead to the apoptosis of cancer cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Gene types of human colorectal cancer cells.
| Cell | KRAS | NRAS | BRAF |
|---|---|---|---|
| DLD-1 | G13D | Wild type | Wild type |
| Hct116 | G13D | Wild type | Wild type |
| RKO | Wild type | Wild type | V600E |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Han, J.H.; Lee, E.-J.; Park, W.; Choi, J.-G.; Ha, K.-T.; Chung, H.-S. Cosmosiin Induces Apoptosis in Colorectal Cancer by Inhibiting PD-L1 Expression and Inducing ROS. Antioxidants 2023, 12, 2131. https://doi.org/10.3390/antiox12122131
AMA Style
Han JH, Lee E-J, Park W, Choi J-G, Ha K-T, Chung H-S. Cosmosiin Induces Apoptosis in Colorectal Cancer by Inhibiting PD-L1 Expression and Inducing ROS. Antioxidants. 2023; 12(12):2131. https://doi.org/10.3390/antiox12122131
Chicago/Turabian StyleHan, Jung Ho, Eun-Ji Lee, Wonyoung Park, Jang-Gi Choi, Ki-Tae Ha, and Hwan-Suck Chung. 2023. "Cosmosiin Induces Apoptosis in Colorectal Cancer by Inhibiting PD-L1 Expression and Inducing ROS" Antioxidants 12, no. 12: 2131. https://doi.org/10.3390/antiox12122131
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.