
The Functional Profile and Antioxidant Capacity of Tomato Fruits Are Modulated by the Interaction between Microbial Biostimulants, Soil Properties, and Soil Nitrogen Status

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions and Experimental Design
2.2. Carotenoids Determination
2.3. Preparation of Extracts for Total Phenolic, Flavonoids and Biological Activity Assays
2.4. Antioxidant and enzymatic assays
2.4.1. Determination of Total Phenolics Content
2.4.2. Determination of Antioxidant and Enzyme Inhibitory Effects
2.5. UHPLC-ESI/QTOF-MS Untargeted Profiling of Tomato Fruits Polyphenols
2.6. Statistical Analyses
3. Results
3.1. Yield Parameters
3.2. Carotenoids Determination
3.3. Total Bioactive Compounds Determination
3.4. Total Antioxidant Activity
3.5. Radical Scavenging Activity
3.6. Reducing Power
3.7. Metal Chelating Activity on Ferrous Ions
3.8. Enzyme Inhibitory Activity
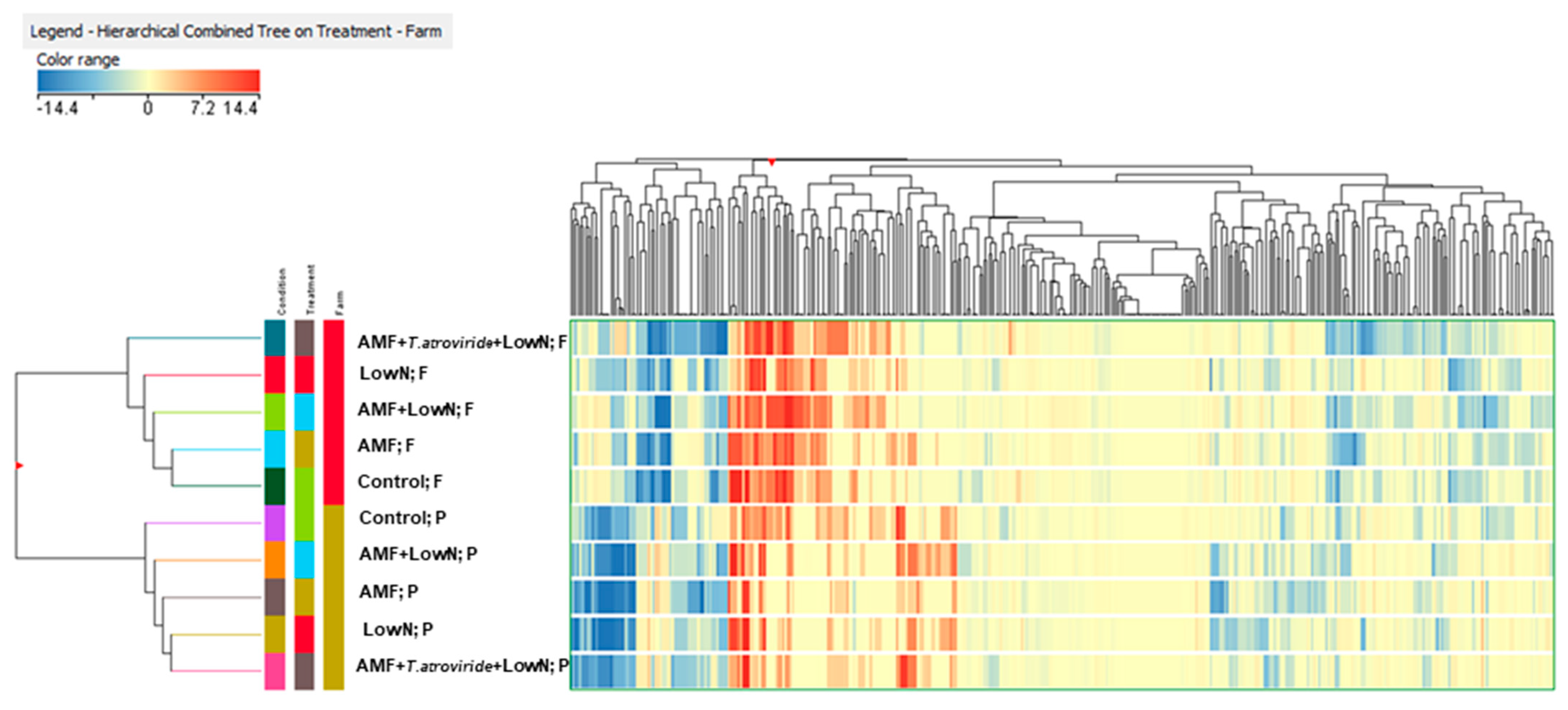
3.9. Metabolomics Untargeted Analysis of the Phenolic Composition of Tomato Fruits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein hydrolysate stimulates growth in tomato coupled with N-dependent gene expression involved in N assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, A.M.; Mannino, G.; Contartese, V.; Bertea, C.M.; Ertani, A. Microbial biostimulants as response to modern agriculture needs: Composition, role and application of these innovative products. Plants 2021, 10, 1533. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Baldotto, L.E.B.; Baldotto, M.A.; Canellas, L.P.; Bressan-Smith, R.; Olivares, F.L. Growth promotion of pineapple’Vitória’by humic acids and Burkholderia spp. during acclimatization. Rev. Bras. Cienc. Solo 2010, 34, 1593–1600. [Google Scholar] [CrossRef]
- Bettoni, M.M.; Mogor, Á.F.; Pauletti, V.; Goicoechea, N. Growth and metabolism of onion seedlings as affected by the application of humic substances, mycorrhizal inoculation and elevated CO2. Sci. Hort. 2014, 180, 227–235. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef]
- Furlan, A.; Bianucci, E.; Sequeira, M.; Álvarez, L.; Peralta, J.M.; Valente, C.; Guarnieri, V.; Castro, S. Combined application of microbial and non-microbial biostimulants to improve growth of peanut plants exposed to abiotic stresses. In Microbial Probiotics for Agricultural Systems; Zúñiga-Dávila, D., González-Andrés, F., Ormeño-Orrillo, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 239–256. [Google Scholar]
- De Vos, R.C.; Hall, R.D.; Moing, A. Metabolomics of a model fruit: Tomato. Biol. Plant Metab. 2011, 43, 109–155. [Google Scholar]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Ganugi, P.; Martinelli, E.; Lucini, L. Microbial biostimulants as a sustainable approach to improve the functional quality in plant-based foods: A review. Curr. Opin. Food Sci. 2021, 41, 217–223. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Understanding the biostimulant action of vegetal-derived protein hydrolysates by high-throughput plant phenotyping and metabolomics: A case study on tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Abou Chehade, L.; Al Chami, Z.; De Pascali, S.A.; Cavoski, I.; Fanizzi, F.P. Biostimulants from food processing by-products: Agronomic, quality and metabolic impacts on organic tomato (Solanum lycopersicum L.). J. Sci. Food Agric. 2018, 98, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Rivero, J.; Gamir, J.; Aroca, R.; Pozo, M.J.; Flors, V. Metabolic transition in mycorrhizal tomato roots. Front. Microbiol. 2015, 6, 598. [Google Scholar] [CrossRef]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Balestrini, R. The association with two different arbuscular mycorrhizal fungi differently affects water stress tolerance in tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef]
- González-González, M.F.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Sánchez-Hernández, C.V.; Casarrubias-Castillo, K.; Becerril-Espinosa, A.; Hernández-Herrera, R.M. Physiological, ecological, and biochemical implications in tomato plants of two plant biostimulants: Arbuscular mycorrhizal fungi and seaweed extract. Front. Plant Sci. 2020, 11, 999. [Google Scholar] [CrossRef]
- Hart, M.; Ehret, D.L.; Krumbein, A.; Leung, C.; Murch, S.; Turi, C.; Franken, P. Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 2015, 25, 359–376. [Google Scholar] [CrossRef]
- Hsu, B.Y.; Pu, Y.S.; Inbaraj, B.S.; Chen, B.H. An improved high performance liquid chromatography–diode array detection–mass spectrometry method for determination of carotenoids and their precursors phytoene and phytofluene in human serum. J. Chromatogr. B 2012, 899, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Salomon, M.V.; Piccoli, P.; Fontana, A. Simultaneous determination of carotenoids with different polarities in tomato products using a C30 core-shell column based approach. Microchem. J. 2020, 159, 105390. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A. Investigation Of Antioxidant Potentials Of Solvent Extracts From Different Anatomical Parts Of Asphodeline Anatolica E. Tuzlaci: An Endemic Plant To Turkey. Afr. J. Trad., Compl. Alt. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Sengul, U.; Zengin, G.; Locatelli, M.; Bahadori, B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharm. 2017, 8, 290. [Google Scholar]
- Rocchetti, G.; Senizza, B.; Putnik, P.; Kovačević, D.B.; Barba, F.J.; Trevisan, M.; Lucini, L. Untargeted screening of the bound/free phenolic composition in tomato cultivars for industrial transformation. J. Sci. Food Agric. 2019, 99, 6173–6181. [Google Scholar] [CrossRef]
- Zuluaga, M.Y.A.; Milani, K.M.L.; Miras-Moreno, M.B.; Lucini, L.; Valentinuzzi, F.; Mimmo, T.; de Oliveira, A.L.M. The adaptive metabolomic profile and functional activity of tomato rhizosphere are revealed upon PGPB inoculation under saline stress. Env. Exp. Bot. 2021, 189, 104552. [Google Scholar] [CrossRef]
- Rocchetti, G.; Blasi, F.; Montesano, D.; Ghisoni, S.; Marcotullio, M.C.; Sabatini, S.; Lucini, L. Impact of conventional/non-conventional extraction methods on the untargeted phenolic profile of Moringa oleifera leaves. Food Res. Int. 2019, 115, 319–327. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. An. Bioch. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Zengin, G.; Mollica, A.; Aktumsek, A.; Picot, C.M.N.; Mahomoodally, M.F. In vitro and in silico insights of Cupressus sempervirens, Artemisia absinthium and Lippia triphylla: Bridging traditional knowledge and scientific validation. EuJIM 2017, 12, 135–141. [Google Scholar] [CrossRef]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions: A Farm to Fork Strategy for a Fair, Healthy and Environmentally-Friendly Food System COM/2020/381 Final. 2020. Available online: https://www.eea.europa.eu/policy-documents/communication-from-the-commission-to-1 (accessed on 9 December 2022).
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular mycorrhizal fungi trigger transcriptional expression of flavonoid and chlorogenic acid biosynthetic pathways genes in tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Rodriguez-Heredia, M.; Djian-Caporalino, C.; Ponchet, M.; Lapeyre, L.; Canaguier, R.; Fazari, A.; Offroy-Chave, M. Protective effects of mycorrhizal association in tomato and pepper against Meloidogyne incognita infection, and mycorrhizal networks for early mycorrhization of low mycotrophic plants. Phytopat. Medit. 2020, 59, 377–384. [Google Scholar]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The role of carotenoids in the prevention of human pathologies. Biomed. Pharmacother. 2004, 58, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Chalker-Scott, L.; Fuchigami, L.H. The role of phenolic compounds in plant stress responses. In Low Temperature Stress Physiology in Crops; Li, P.H., Ed.; CRC press: Boca Rato, FL, USA, 1989; pp. 67–80. [Google Scholar]
- Vetrani, C.; Costabile, G.; Vitale, M.; Giacco, R. (Poly) phenols and cardiovascular diseases: Looking in to move forward. JFF 2020, 71, 104013. [Google Scholar] [CrossRef]
- Brunner, K.; Zeilinger, S.; Ciliento, R.; Woo, S.L.; Lorito, M.; Kubicek, C.P.; Mach, R.L. Improvement of the fungal biocontrol agent Trichoderma atroviride to enhance both antagonism and induction of plant systemic disease resistance. AEM 2005, 71, 3959–3965. [Google Scholar] [CrossRef]
- Coppola, M.; Cascone, P.; Lelio, I.D.; Woo, S.L.; Lorito, M.; Rao, R.; Digilio, M.C. Trichoderma atroviride P1 colonization of tomato plants enhances both direct and indirect defense barriers against insects. Front. Physiol. 2019, 10, 813. [Google Scholar] [CrossRef]
- Macías-Rodríguez, L.; Guzmán-Gómez, A.; García-Juárez, P.; Contreras-Cornejo, H.A. Trichoderma atroviride promotes tomato development and alters the root exudation of carbohydrates, which stimulates fungal growth and the biocontrol of the phytopathogen Phytophthora cinnamomi in a tripartite interaction system. FEMS Microb. Ecol. 2019, 94, fiy137. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.A. The Use of Arbuscular Mycorrhizal Fungi in Combination with Trichoderma spp. in Sustainable Agriculture. In Bioformulations: For Sustainable Agriculture; Arora, N., Mehnaz, S., Balestrini, R., Eds.; Springer: New Delhi, India; pp. 137–146.
- Alonso-Ramírez, A.; Poveda, J.; Martín, I.; Hermosa, R.; Monte, E.; Nicolás, C. Salicylic acid prevents Trichoderma harzianum from entering the vascular system of roots. Mol. Plant Pathol. 2014, 15, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Mendoza, A.; Zaid, R.; Lawry, R.; Hermosa, R.; Monte, E.; Horwitz, B.A.; Mukherjee, P.K. Molecular dialogues between Trichoderma and roots: Role of the fungal secretome. Fungal Biol. Rev. 2018, 32, 62–85. [Google Scholar] [CrossRef]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolás, C. Trichoderma harzianum favours the access of arbuscular mycorrhizal fungi to non-host Brassicaceae roots and increases plant productivity. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Thirkell, T.; Cameron, D.; Hodge, A. Contrasting nitrogen fertilisation rates alter mycorrhizal contribution to barley nutrition in a field trial. Front. Plant Sci. 2019, 10, 1312. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Bicharanloo, B.; Cavagnaro, T.R.; Keitel, C.; Dijkstra, F.A. Nitrogen fertilisation increases specific root respiration in ectomycorrhizal but not in arbuscular mycorrhizal plants: A meta-analysis. Front. Plant Sci. 2021, 12, 711720. [Google Scholar] [CrossRef]
- Savolainen, T.; Kytöviita, M.M. Mycorrhizal symbiosis changes host nitrogen source use. Plant Soil 2022, 471, 643–654. [Google Scholar] [CrossRef]
- Yang, H.; Fang, C.; Li, Y.; Wu, Y.; Fransson, P.; Rillig, M.C.; Weih, M. Temporal complementarity between roots and mycorrhizal fungi drives wheat nitrogen use efficiency. New Phytol. 2022, 236, 1168–1181. [Google Scholar] [CrossRef]
- Lin, C.; Wang, Y.; Liu, M.; Li, Q.; Xiao, W.; Song, X. Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, E.B.; Egerton-Warburton, L.M.; Hart, M.M.; Klironomos, J.N.; Maherali, H.; Tedersoo, L. Arbuscular mycorrhizal fungi as mediators of ecosystem responses to nitrogen deposition: A trait-based predictive framework. J. Ecol. 2018, 106, 480–489. [Google Scholar] [CrossRef]
- Treseder, K.K. A Meta-analysis of Mycorrhizal Responses to Nitrogen, Phosphorus, and Atmospheric CO 2 in Field Studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, G.; Mao, L.; Cheng, G.; Jiang, S.; Ma, X. Direct and Indirect Influences of 8 Yr of Nitrogen and Phosphorus Fertilization on Glomeromycota in an alpine Meadow Ecosystem. New Phytol. 2012, 194, 523–535. [Google Scholar] [CrossRef]
- Emery, S.M.; Bell-Dereske, L.; Stahlheber, K.A.; Gross, K.L. Arbuscular mycorrhizal fungal community responses to drought and nitrogen fertilization in switchgrass stands. Appl. Soil Ecol. 2022, 169, 104218. [Google Scholar] [CrossRef]
- Muneer, M.A.; Tarin, M.W.K.; Chen, X.; Afridi, M.S.; Iqbal, A.; Munir, M.Z.; Ji, B. Differential response of mycorrhizal fungi linked with two dominant plant species of temperate grassland under varying levels of N-addition. Appl. Soil Ecol. 2022, 170, 104272. [Google Scholar] [CrossRef]
- Ganugi, P.; Fiorini, A.; Ardenti, F.; Caffi, T.; Bonini, P.; Taskin, E.; Puglisi, E.; Tabaglio, V.; Lucini, L. Nitrogen use efficiency, rhizosphere bacterial community, and root metabolome reprogramming due to maize seed treatment with microbial biostimulants. Physiol. Plant 2022, 174, e13679. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Wang, X.; Sun, Y.; Cheng, Y.; Liu, S.; Chen, X.; Kuyper, T.W. Arbuscular mycorrhizal fungi negatively affect nitrogen acquisition and grain yield of maize in a N deficient soil. Front. Microb. 2018, 9, 418. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, J.; Yang, Y.; Wang, Y.; Zhang, Y.; Wang, P. Effects of Conventional and Organic Agriculture on Soil Arbuscular Mycorrhizal Fungal Community in Low-Quality Farmland. Front. Microb. 2022, 13, 91462. [Google Scholar] [CrossRef] [PubMed]
- Hoeksema, J.D.; Chaudhary, V.B.; Gehring, C.A.; Johnson, N.C.; Karst, J.; Koide, R. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Zaller, J.G.; Frank, T.; Drapela, T. Soil sand content can alter effects of different taxa of mycorrhizal fungi on plant biomass production of grassland species. Eur. J. Soil Biol. 2011, 47, 175–181. [Google Scholar] [CrossRef]
- Shen, K.; He, Y.; Xu, X.; Umer, M.; Liu, X.; Xia, T.; Yan, J. Effects of AMF on plant nutrition and growth depend on substrate gravel content and patchiness in the karst species Bidens pilosa L. Front. Plant Sci. 2022, 13, 968719. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Yield Parameters | |||||
|---|---|---|---|---|---|
| Marketable (t/ha) | Immature (t/ha) | Rotten (t/ha) | Apical Rot (t/ha) | Total Yield (t/ha) | |
| Treatment (T) | |||||
| LowN | 68.0 | 5.4 | 1.4 | 1.2 a | 76.1 |
| AMF | 65.8 | 3.8 | 1.1 | 0.8 a | 71.5 |
| AMF + LowN | 70.7 | 4.8 | 1.4 | 1.5 a | 78.4 |
| AMF + T. atr. + LowN | 71.9 | 10.1 | 1.4 | 1.5 a | 84.9 |
| Control | 70.2 | 7.9 | 0.8 | 2.1 b | 81.0 |
| Significance | |||||
| T (df = 5) Residuals (df = 18) | n.s. | n.s. | n.s. | n.s. | |
| Yield Parameters | |||||
|---|---|---|---|---|---|
| Marketable (t/ha) | Immature (t/ha) | Rotten (t/ha) | Apical Rot (t/ha) | Total Yield (t/ha) | |
| Treatment (T) | |||||
| LowN | 82.7 b | 13 | 2.1 | 0.4 | 98.2 b |
| AMF | 88.8 a | 12.3 | 3.1 | 0.5 | 104.7 a |
| AMF + LowN | 78.6 b | 7.4 | 1.9 | 0.5 | 88.5 b |
| AMF + T. atr. + LowN | 88.0 a | 10.2 | 2.7 | 1.0 | 101.9 a |
| Control | 89.0 a | 12.1 | 3.1 | 0.8 | 104.9 a |
| Significance | |||||
| T (df = 5) Residuals (df = 18) | n.s. | n.s. | n.s. | ||
| Source of Variance | Phytoene mg/100 g | Phytofluene mg/100 g | Z-b-carotene mg/100 g | B-carotene mg/100 g | Z-carotene mg/100 g | All-E-y-carotene mg/100 g | 13-z-lycopene mg/100 g | 7-z-lycopene mg/100 g | 9-z-lycopene mg/100 g | Cis-Lycopene mg/100 g | All-Trans-Lycopene mg/100 g |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Thesis (T) | |||||||||||
| LowN | 3.278 ± 0.649 a | 1.233 ± 0.015 a | 0.131 ± 0.141 | 0.598 ± 0.262 | 0.020 ± 0.001 | 0.063 ± 0.001 a | 0.126 ± 0.061 | 0.151 ± 0.06 | 0.020 ± 0.001 a | 0.247 ± 0.023 | 11.982 ± 4.985 |
| AMF | 1.821 ± 1.136 c | 0.599 ± 0.138 c | 0.044 ± 0.021 | 0.433 ± 0.103 | 0.006 ± 0.009 | 0.045 ± 0.004 b | 0.103 ± 0.020 | 0.112 ± 0.022 | 0 b | 0.175 ± 0.043 | 9.451 ± 1.763 |
| AMF + LowN | 2.521 ± 0.475 b | 0.839 ± 0.141 b | 0.033 ± 0.010 | 0.543 ± 0.088 | 0.008 ± 0.014 | 0.063 ± 0.010 a | 0.142 ± 0.037 | 0.175 ± 0.036 | 0.008 ± 0.012 b | 0.256 ± 0.044 | 12.415 ± 1.648 |
| AMF + T. atr. + LowN | 2.599 ± 0.424 b | 0.856 ± 0.293 b | 0.038 ± 0.011 | 0.647 ± 0.243 | 0.021 ± 0.021 | 0.063 ± 0.022 a | 0.145 ± 0.052 | 0.154 ± 0.037 | 0.007 ± 0.012 b | 0.233 ± 0.051 | 12.586 ± 1.511 |
| Control | 3.426 ± 0.424 a | 1.042 ± 0.099 ab | 0.082 ± 0.079 | 0.486 ± 0.189 | 0.014 ± 0.021 | 0.071 ± 0.015 a | 0.135 ± 0.053 | 0.191 ± 0.074 | 0.008 ± 0.012 b | 0.215 ± 0.017 | 10.435 ± 4.551 |
| Significance | |||||||||||
| T (df = 4)—p value | 3.23 × 10-5 | 9.63 × 10-6 | 0.264 | 0.337 | 0.374 | 0.0297 | 0.556 | 0.106 | 0.0178 | 0.184 | 0.184 |
| Residuals (df = 25) F value | 10.78 | 12.58 | 1.397 | 1.197 | 1.109 | 3.202 | 0.768 | 2.137 | 3.651 | 1.687 | 1.687 |
| Source of Variance | Phytoene mg/100 g | Phytofluene mg/100 g | Z-b-carotene mg/100 g | B-carotene mg/100 g | Z-carotene mg/100 g | All-E-y-carotene mg/100 g | 13-z-lycopene mg/100 g | 7-z-lycopene mg/100 g | 9-z-lycopene mg/100 g | Cis-Lycopene mg/100 g | All-Trans-Lycopene mg/100 g |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Thesis (T) | |||||||||||
| LowN | 3.718 ± 0.649 | 1.042 ± 0.099 a | 0.315 ± 0.022 | 0.436 ± 0.070 bc | 0 b | 0.079 ± 0.008 | 0.215 ± 0.020 | 0.262 ± 0.037 | 0.024 ± 0.002 | 0.418 ± 0.0766 | 16.781 ± 1.797 |
| AMF | 3.310 ± 1.182 | 1.056 ± 0.280 ab | 0.255 ± 0.055 | 0.332 ± 0.099 c | 0.008 ± 0.013 ab | 0.061 ± 0.022 | 0.170 ± 0.069 | 0.182 ± 0.073 | 0.015 ± 0.012 | 0.274 ± 0.122 | 12.201 ± 3.562 |
| AMF + LowN | 2.611 ± 0.291 | 0.827 ± 0.088 a | 0.279 ± 0.034 | 0.573 ± 0.082 a | 0.020 ± 0.002 a | 0.072 ± 0.014 | 0.179 ± 0.039 | 0.191 ± 0.043 | 0.019 ± 0.003 | 0.315 ± 0.085 | 13.694 ± 2.439 |
| AMF + T. atr. + LowN | 2.763 ± 1.997 | 1.051 ± 0.087 ab | 0.295 ± 0.018 | 0.647 ± 0.243 c | 0.009 ± 0.014 ab | 0.072 ± 0.011 | 0.200 ± 0.206 | 0.206 ± 0.005 | 0.021 ± 0.001 | 0.349 ± 0.027 | 14.549 ± 0.225 |
| Control | 3.563 ± 1.263 | 1.190 ± 0.263 a | 0.310 ± 0.041 | 0.552 ± 0.158 ab | 0.009 ± 0.014 b | 0.079 ± 0.021 | 0.220 ± 0.060 | 0.278 ± 0.113 | 0.023 ± 0.006 | 0.468 ± 0.235 | 16.072 ± 4.187 |
| Significance | |||||||||||
| T (df = 4)—p value | 0.0668 | 0.0267 | 0.514 | 0.00209 | 0.0265 | 0.65 | 0.282 | 0.0599 | 0.121 | 0.102 | 0.0632 |
| Residuals (df = 25) F value | 2.517 | 3.294 | 2.735 | 5.709 | 3.302 | 0.624 | 1.344 | 2.607 | 2.027 | 2.171 | 2.563 |
| Source of Variance | Total Phenolic Content mgGAE/g | Total Flavonoid Content mgRE/g | DPPH mgTE/g | ABTS mgTE/g | CUPRAC mgTE/g | FRAP mgTE/g | Metal Chelating mgEDTAE/g | Phosphomolybdenum mmolTE/g | AChe inh. mgGALAE/g | BChe inh. mgGALAE/g | Tyrosinase mgKAE/g | Amylase mmolACAE/g |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Thesis (T) | ||||||||||||
| LowN | 11.9 ± 2.0 | 0.8 ± 0.1 b | 1.6 ± 0.4 | 23.0 ± 2.9 | 33.3 ± 3.8 ab | 20.4 ± 1.1 ab | 15.4 ± 1.4 | 0.8 ± 0.1 ab | 2.3 ± 0.1 | 2.4 ± 0.3 | 56.9 ± 4.9 c | 0.21 ± 0.0 |
| AMF | 11.9 ± 2.2 | 0.9 ± 0.1 ab | 1.4 v 1.3 | 21.6 ± 4.4 | 33.4 ± 5.0 ab | 20.6 ± 2.5 ab | 13.6 ± 1.8 | 0.9 ± 0.1 a | 2.4 ± 0.1 | 2.2 ± 0.6 | 54.9 ± 4.2 bc | 0.20 ± 0.0 |
| AMF + LowN | 10.6 ± 1.1 | 0.9 ± 0.2 a | 1.6 ± 0.7 | 20.3 ± 2.5 | 29.5 ± 1.9 b | 19.6 ± 0.8 b | 14.0 ± 2.8 | 0.7 ± 0.1 b | 2.2 ± 0.1 | 2.4 ± 0.4 | 56.9 ± 6.2 abc | 0.21 ± 0.0 |
| AMF + T. atr. + LowN | 12.9 ± 2.9 | 0.8 ± 0.1ab | 3.4 ± 3.7 | 24.5 ± 8.3 | 37.4 ± 7.6a | 23.6 ± 6.3a | 15.2 ± 1.6 | 0.9 ± 0.2a | 2.4 ± 0.1 | 2.3 ± 0.4 | 62.2 ± 3.1ab | 0.21 ± 0.0 |
| Control | 11.2 ± 1.3 | 0.8 ± 0.1b | 1.2 ± 0.4 | 21.2 ± 0.8 | 31.2 ± 1.8b | 19.2 ± 1.0b | 14.4 ± 2.5 | 0.8 ± 0.1ab | 2.2 ± 0.1 | 2.7 ± 0.9 | 63.5 ± 4.6a | 0.21 ± 0.0 |
| Significance | ||||||||||||
| T (df = 4)—p value | 0.166 | 0.0452 | 0.0964 | 0.331 | 0.0114 | 0.0431 | 0.282 | 0.0483 | 0.00116 | 0.551 | 0.0315 | 0.104 |
| Residuals (df = 40) F value | 1.712 | 2.681 | 2.118 | 1.188 | 3.73 | 2.717 | 1.313 | 2.632 | 5.599 | 0.77 | 2.951 | 2.064 |
| Source of Variance | Total Phenolic Content mgGAE/g | Total Flavonoid Content mgRE/g | DPPH mgTE/g | ABTS mgTE/g | CUPRAC mgTE/g | FRAP mgTE/g | Metal Chelating mgEDTAE/g | Phosphomolybdenum mmolTE/g | AChe inh. mgGALAE/g | BChe inh. mgGALAE/g | Tyrosinase mgKAE/g | Amylase mmolACAE/g |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Thesis (T) | ||||||||||||
| LowN | 14.4 ± 1.7 a | 0.9 ± 0.1 bc | 3.2 ± 1.6 a | 29.8 ± 1.7 a | 35.6 ± 5.7 | 21.8 ± 2.2 a | 14.9 ± 1.4 a | 0.9 ± 0.1 | 2.4 ± 0.2 | 2.8 ± 0.7 | 53.9 ± 4.7 | 0.19 ± 0.0 |
| AMF | 12.7 ± 0.4 b | 0.8 ± 0.1 c | 1.4 ± 05 b | 23.2 ± 1.4 c | 32.5 ± 1.6 | 19.5 ± 0.6 bc | 12.5 ± 1.2 b | 0.9 ± 0.1 | 2.4 ± 0.2 | 3.0 ± 0.7 | 54.7 ± 3.5 | 0.18 ± 0.0 |
| AMF + LowN | 12.4 ± 0.8 b | 1.0 ± 0.2 ab | 1.5 ± 0.2 b | 23.6 ± 0.9 c | 31.6 ± 2.5 | 19.2 ± 0.9 c | 11.5 ± 1.7 b | 0.8 ± 0.1 | 2.3 ± 0.1 | 2.9 ± 0.4 | 53.8 ± 3.2 | 0.19 ± 0.0 |
| AMF + T.atr. + LowN | 12.5 ± 1.7 b | 0.8 ± 0.2 bc | 1.6 ± 1.4 b | 24.8 ± 3.1 bc | 32.9 ± 3.6 | 19.7 ± 1.4 bc | 12.7 ± 2.7 b | 0.9 ± 0.1 | 2.4 ± 0.1 | 3.1 ± 0.3 | 53.9 ± 4.1 | 0.19 ± 0.0 |
| Control | 13.4 ± 0.6 ab | 1.2 ± 0.3 a | 1.8 ± 0.1 b | 27.1 ± 3.8 b | 34.7 ± 2.1 | 20.7 ± 1.2 ab | 12.4 ± 1.7 b | 0.9 ± 0.1 | 2.4 ± 0.1 | 2.7 ± 0.6 | 52.9 ± 2.3 | 0.19 ± 0.0 |
| Significance | ||||||||||||
| T (df = 4)—p value | 0.00368 | 0.00118 | 0.00286 | 3.69 x 10-6 | 0.117 | 0.00156 | 0.00586 | 0.96 | 0.581 | 0.686 | 0.29 | 0.833 |
| Residuals (df = 40) F value | 4.619 | 5.561 | 4.824 | 11.12 | 1.976 | 5.323 | 4.247 | 0.153 | 0.723 | 0.571 | 1.29 | 0.364 |
| Source of Variance | Flavonoids mg eq. g−1 DM | Lignans mg eq. g−1 DM | Other Polyphenols mg eq. g−1 DM | Phenolic Acids mg eq. g−1 DM | Stilbenes mg eq. g−1 DM |
|---|---|---|---|---|---|
| Thesis (T) | |||||
| LowN | 4.7 ± 1.8 b | 13.2 ± 1.2 a | 52.6 ± 14.9 a | 27.7 ± 4.5 | 0.4 ± 0.2 |
| AMF | 5.2 ± 0.8 ab | 11.7 ± 3.3 ab | 37.7 ± 15.9 ab | 28.2 ± 5.6 | 0.4 ± 0.1 |
| AMF + LowN | 6.7 ± 1.4 a | 6.4 ± 1.8 c | 22.1 ± 4.6 b | 21.9 ± 8.7 | 0.5 ± 0.1 |
| AMF + T. atr. + LowN | 4.5 ± 1.1 b | 8.4 ± 3.7 bc | 24.8 ± 13.3 b | 17.1 ± 9.7 | 0.5 ± 0.1 |
| Control | 6.1 ± 0.9 ab | 11.4 ± 2.7 ab | 30.9 ± 14.1 b | 21.9 ± 8.7 | 0.5 ± 0.1 |
| Significance | |||||
| T (df = 4)—p value | 0.024 | 0.00189 | 0.00382 | 0.106 | 0.566 |
| Residuals (df = 25) F value | 3.388 | 5.815 | 5.098 | 2.136 | 0.752 |
| Source of Variance | Flavonoids mg eq. g−1 DM | Lignans mg eq. g−1 DM | Other Polyphenols mg eq. g−1 DM | Phenolic Acids mg eq. g−1 DM | Stilbenesmg eq. g−1 DM |
|---|---|---|---|---|---|
| Thesis (T) | |||||
| LowN | 3.5 ± 0.7 | 13.0 ± 1.9 | 55.0 ± 11.5 | 30.8 ± 7.2 | 0.5 ± 0.2 |
| AMF | 2.4 ± 0.3 | 12.8 ± 3.1 | 44.2 ± 16.2 | 32.4 ± 6.2 | 0.5 ± 0.1 |
| AMF + LowN | 3.1 ± 1.0 | 10.9 ± 4.9 | 31.9 ± 13.6 | 24.0 ± 11.7 | 0.5 ± 0.2 |
| AMF + T. atr. + LowN | 3.7 ± 1.0 | 11.2 ± 4.0 | 37.9 ± 19.3 | 27.3 ± 12.8 | 0.6 ± 0.1 |
| Control | 3.8 ± 1.4 | 12.7 ± 2.8 | 37.4 ± 15.7 | 27.7 ± 9.6 | 0.6 ± 0.2 |
| Significance | |||||
| T (df = 4)—p value | 0.105 | 0.36 | 0.291 | 0.179 | 0.709 |
| Residuals (df = 25) F value | 2.148 | 1.142 | 1.316 | 1.714 | 0.538 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganugi, P.; Fiorini, A.; Tabaglio, V.; Capra, F.; Zengin, G.; Bonini, P.; Caffi, T.; Puglisi, E.; Trevisan, M.; Lucini, L. The Functional Profile and Antioxidant Capacity of Tomato Fruits Are Modulated by the Interaction between Microbial Biostimulants, Soil Properties, and Soil Nitrogen Status. Antioxidants 2023, 12, 520. https://doi.org/10.3390/antiox12020520
Ganugi P, Fiorini A, Tabaglio V, Capra F, Zengin G, Bonini P, Caffi T, Puglisi E, Trevisan M, Lucini L. The Functional Profile and Antioxidant Capacity of Tomato Fruits Are Modulated by the Interaction between Microbial Biostimulants, Soil Properties, and Soil Nitrogen Status. Antioxidants. 2023; 12(2):520. https://doi.org/10.3390/antiox12020520
Chicago/Turabian StyleGanugi, Paola, Andrea Fiorini, Vincenzo Tabaglio, Federico Capra, Gokhan Zengin, Paolo Bonini, Tito Caffi, Edoardo Puglisi, Marco Trevisan, and Luigi Lucini. 2023. "The Functional Profile and Antioxidant Capacity of Tomato Fruits Are Modulated by the Interaction between Microbial Biostimulants, Soil Properties, and Soil Nitrogen Status" Antioxidants 12, no. 2: 520. https://doi.org/10.3390/antiox12020520
APA StyleGanugi, P., Fiorini, A., Tabaglio, V., Capra, F., Zengin, G., Bonini, P., Caffi, T., Puglisi, E., Trevisan, M., & Lucini, L. (2023). The Functional Profile and Antioxidant Capacity of Tomato Fruits Are Modulated by the Interaction between Microbial Biostimulants, Soil Properties, and Soil Nitrogen Status. Antioxidants, 12(2), 520. https://doi.org/10.3390/antiox12020520