


Exogenous Application of Calcium Ameliorates Salinity Stress Tolerance of Tomato (Solanum lycopersicum L.) and Enhances Fruit Quality

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Assessment of Growth and Yield Parameters
2.3. Estimation of Leaf Relative Water Content and Photosynthetic Pigment
2.4. Determination of Proline and Glycine Betaine (GB) Content
2.5. Determination of Hydrogen Peroxide and Melondealdehyde
2.6. Estimation of Antioxidant Enzymatic Activities
2.7. Determination of Na, K, Ca, and Mg Contents
2.8. Estimation of Total Soluble Solids, Reducing Sugar, Total Sugar, β-Carotene, Ascorbic Acid (Vitamin C) and pH
2.8.1. Total Soluble Solids
2.8.2. Estimation of Sugar
2.8.3. Estimation of β-Carotene
2.8.4. Estimation of Ascorbic Acid (Vit-C)
2.8.5. Fruit-Juice pH
2.9. Statistical Analysis
3. Results
3.1. Growth and Biomass Production of Tomato
3.2. Yield Components of Tomato
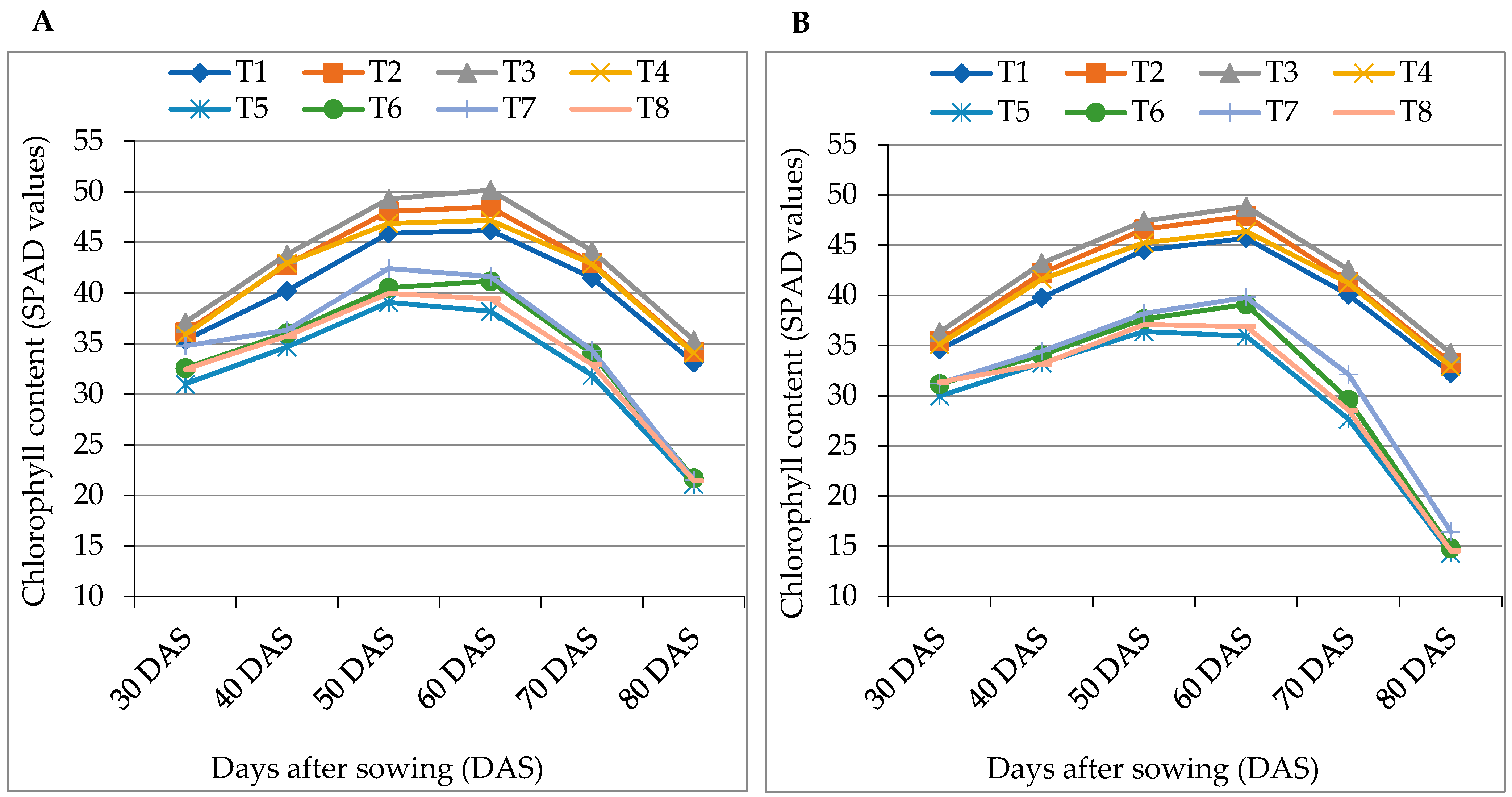
3.3. Relative Water Content and Chlorophyll Content (SPAD values) of Tomato
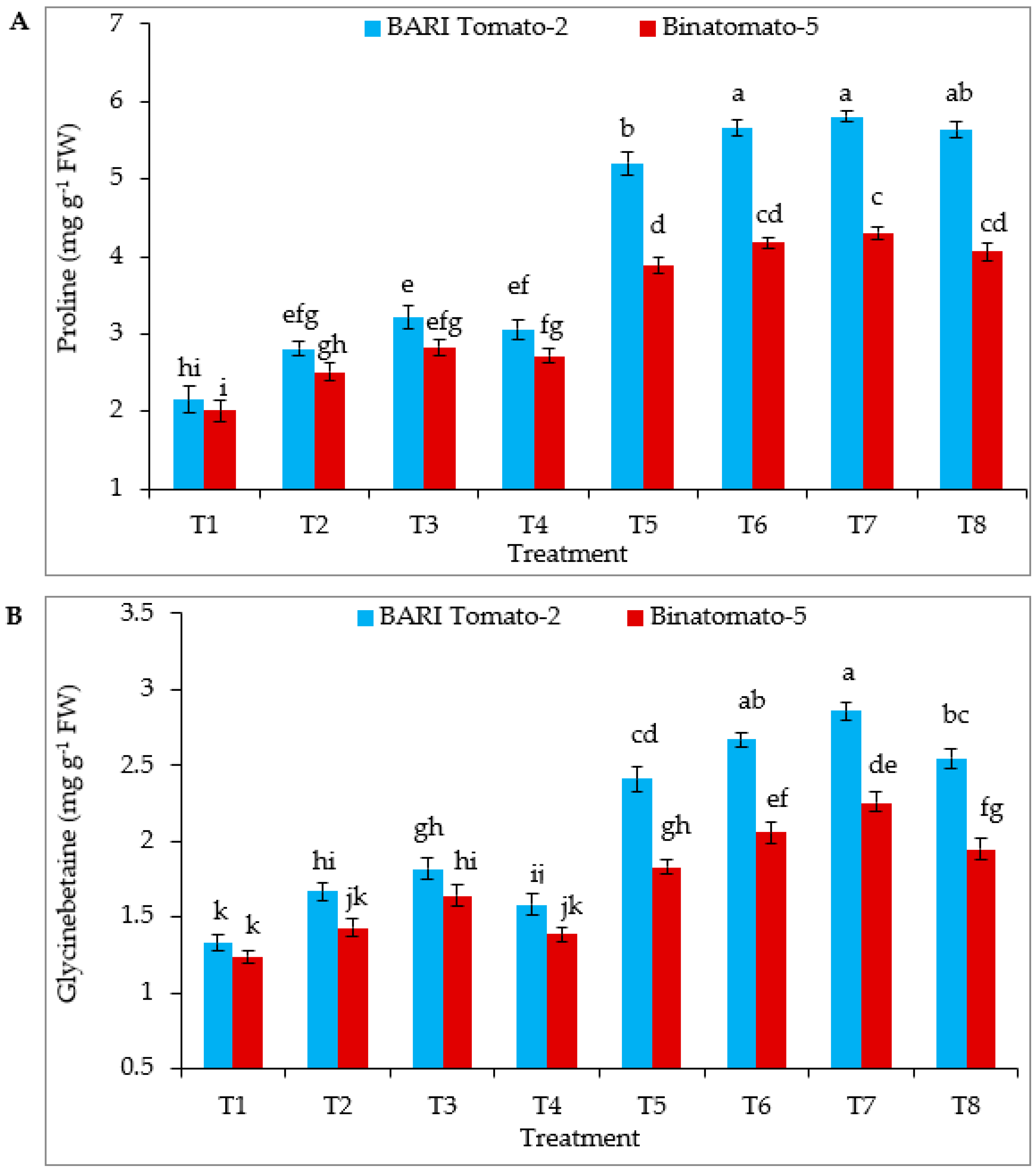
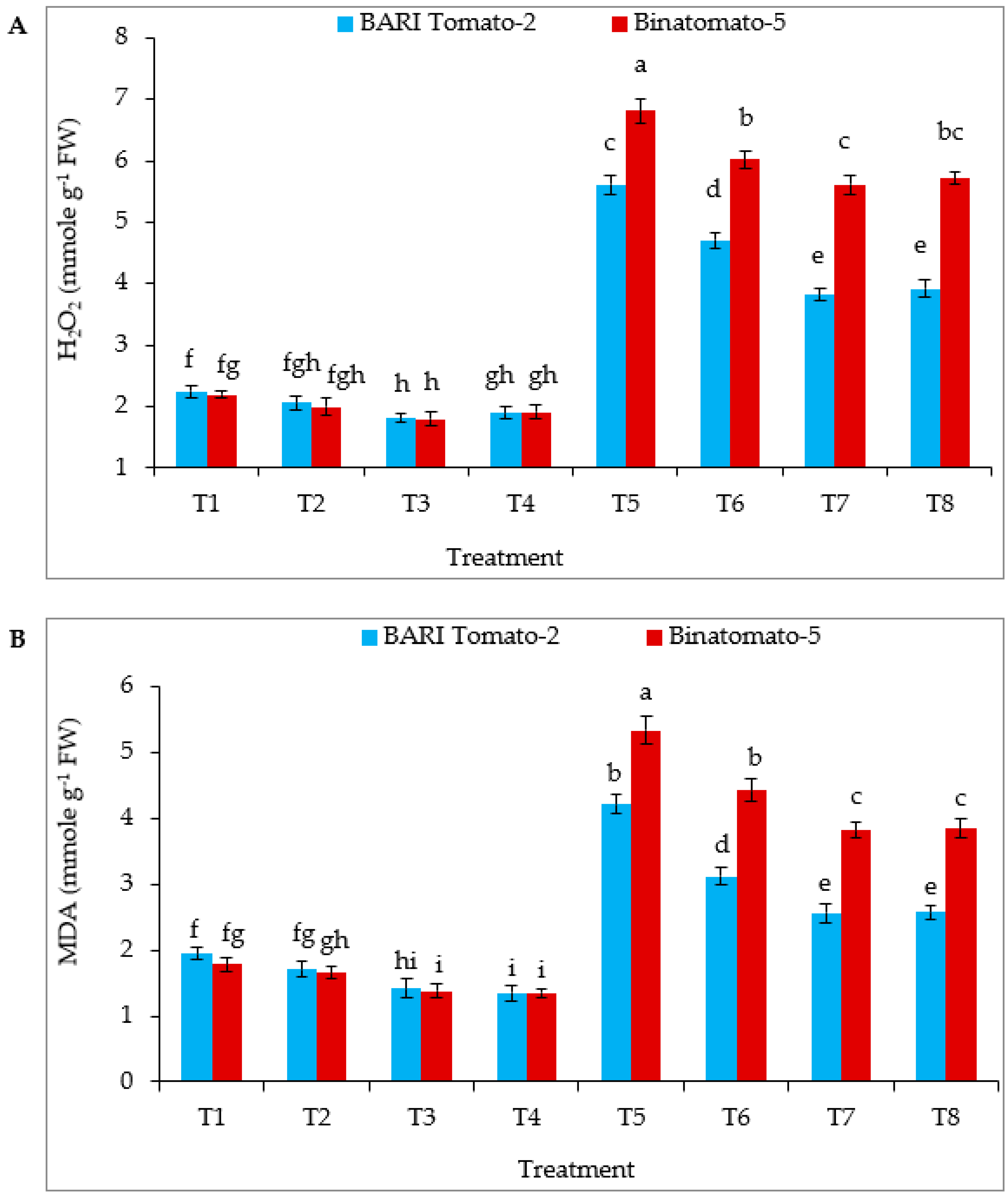
3.4. Proline, Glycinebetaine, Hydrogen Peroxide and Malondialdehyde Content
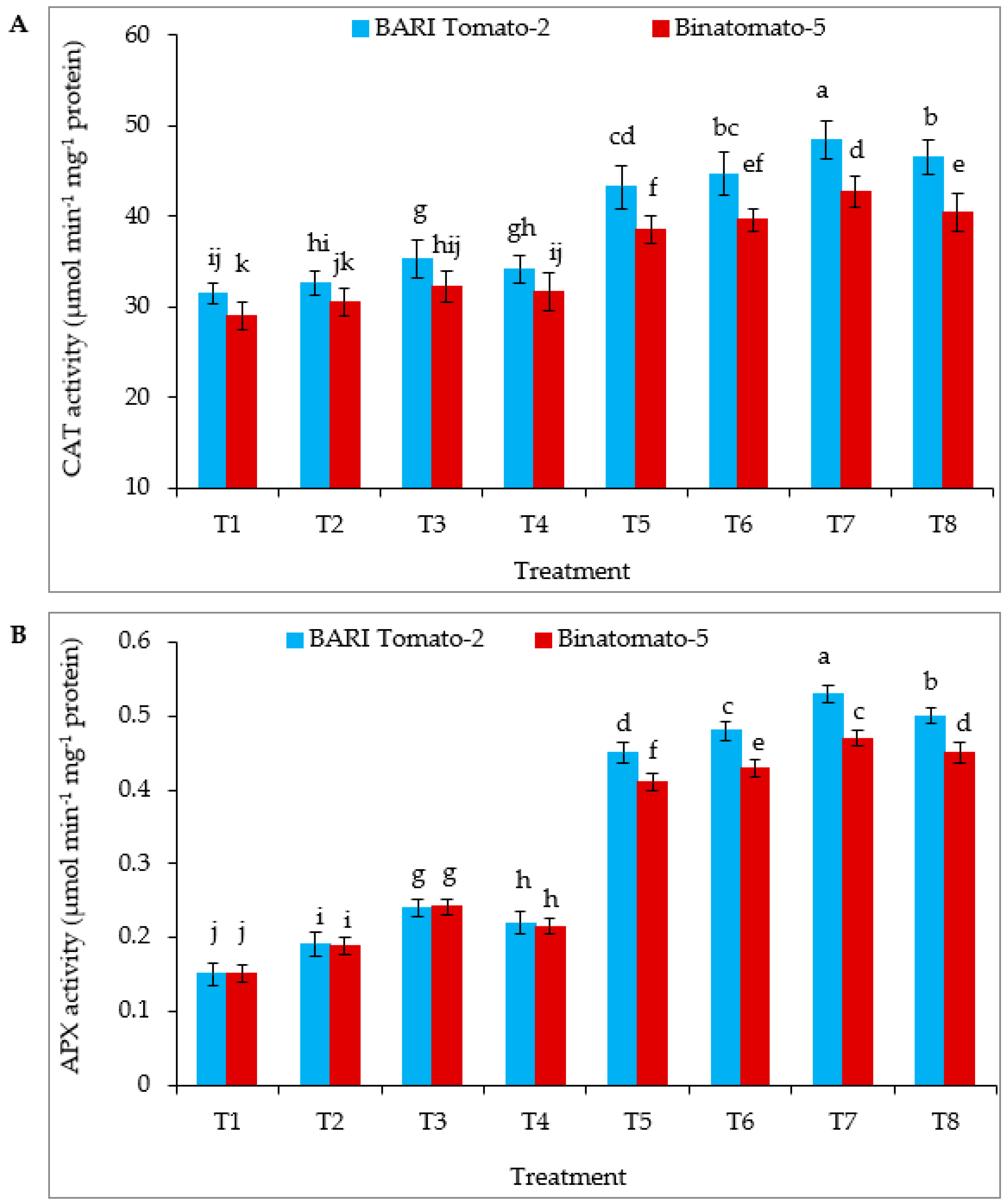
3.5. Antioxidant Enzymes Activity
3.6. Mineral Concentrations
3.7. Fruit-Quality Traits of Tomato
4. Discussion
4.1. Growth and Biomass Production of Tomato
4.2. Yield Components of Tomato
4.3. Relative Water Content and Chlorophyll Content (SPAD Values) of Tomato
4.4. Biochemical Traits of Tomato
4.5. Antioxidant Enzymes Activity
4.6. Mineral Concentrations
4.7. Fruit Quality of Tomato
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molotoks, A.; Smith, P.; Dawson, T.P. Impacts of land use, population, and climate change on global food security. Food Energy Secur. 2021, 10, e261. [Google Scholar] [CrossRef]
- FAO. FAOSTAT, Food and Agriculture Organization, Rome. 2016. Available online: http://www.fao.org/faostat/en/#home (accessed on 1 July 2022).
- Rasool, S.; Ahmad, A.; Siddiqi, T.O.; Ahmad, P. Changes in growth, lipid peroxidation and some key antioxidant enzymes in chickpea genotypes under salt stress. Acta Physiol. Plant. 2013, 35, 1039–1050. [Google Scholar] [CrossRef]
- Dassanayake, M.; Larkin, J.C. Making Plants Break a Sweat, the Structure, Function, and Evolution of Plant Salt Glands. Front. Plant Sci. 2017, 8, 406. [Google Scholar] [CrossRef] [PubMed]
- Otie, V.; Udo, I.; Shao, Y.; Itam, M.O.; Okamoto, H.; An, P.; Eneji, E.A. Salinity Effects on Morpho-Physiological and Yield Traits of Soybean Glycine max L. as Mediated by Foliar Spray with Brassinolide. Plants 2021, 10, 541. [Google Scholar] [CrossRef]
- Hussain, M.; Park, H.W.; Farooq, M.; Jabran, K.; Lee, D.J. Morphological and physiological basis of salt resistance in different rice genotypes. Int. J. Agric. Biol. 2013, 15, 113–118. [Google Scholar]
- Kumari, A.; Parida, A.K. Metabolomics and network analysis reveal the potential metabolites and biological pathways involved in salinity tolerance of the halophyte Salvadora persica. Environ. Exp. Bot. 2018, 148, 85–99. [Google Scholar] [CrossRef]
- Kong, X.; Luo, Z.; Dong, H.; Eneji, A.E.; Li, W. H2O2 and ABA signaling are responsible for the increased Na+ efflux and water uptake in Gossypium hirsutum L. roots in the non-saline side under non-uniform root zone salinity. J. Exp. Bot. 2016, 67, 2247–2261. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Fujita, M. Exogenous Tebuconazole and Trifloxystrobin Regulates Reactive Oxygen Species Metabolism toward Mitigating Salt-Induced Damages in Cucumber Seedling. Plants 2019, 8, 428. [Google Scholar] [CrossRef]
- Safdar, H.; Amin, A.; Shafiq, Y.; Ali, A.; Yasin, R.; Shoukat, A.; Hussan, M.U.; Sarwar, M.I. A review: Impact of salinity on plant growth. Nat. Sci. 2019, 17, 34–40. [Google Scholar]
- Morton, M.J.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negra, O.S.; Tester, M. Salt stress under the scalpel–dissecting the genetics of salt tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Alam, M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous proline and glycine betaine mediated upregulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxidative stress in two rice (Oryza sativa L.) varieties. BioMed Res. Int. 2014, 757219. [Google Scholar] [CrossRef]
- El-Shabrawi, H.; Kumar, B.; Kaul, T.; Reddy, M.K.; Singla-Pareek, S.L.; Sopory, S.K. Redox homeostasis, antioxidant defense, and methylglyoxal detoxification as markers for salt tolerance in Pokkali rice. Protoplasma 2010, 245, 85–96. [Google Scholar] [CrossRef]
- Tada, Y.; Komatsubara, S.; Kurusu, T. Growth and physiological adaptation of whole plants and cultured cells from a halophyte turf grass under salt stress. AoB Plants 2014, 6, plu041. [Google Scholar] [CrossRef]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savoure, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Hurry, V.; Huner, N.P. Interaction of glycine betaine and plant hormones protection of the photosynthetic apparatus during abiotic stress. In Photosynthesis: Structures, Mechanisms, and Applications; Hou, H.J.M., Najafpour, M., Moore, G., Allakhverdiev, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 185–202. [Google Scholar]
- Yan, K.; Shao, H.; Shao, C.; Chen, P.; Zhao, S.; Brestic, M.; Chen, X. Physiological adaptive mechanisms of plants grown in saline soil and implications for sustainable saline agriculture in coastal zone. Acta Physiol. Plant. 2013, 35, 2867–2878. [Google Scholar] [CrossRef]
- Hossain, M.S.; Hasanuzzaman, M.; Sohag, M.M.H.; Bhuyan, M.H.M.B.; Fujita, M. Acetate-induced modulation of ascorbate: Glutathione cycle and restriction of sodium accumulation in shoot confer salt tolerance in Lens culinaris Medik. Physiol. Mol. Biol. Plants 2019, 25, 443–455. [Google Scholar] [CrossRef]
- Tan, H.-L.; Thomas-Ahner, J.M.; Grainger, E.M.; Wan, L.; Francis, D.M.; Schwartz, S.J.; Erdman, J.W.; Clinton, S.K. Tomato-based food products for prostate cancer prevention: What have we learned? Cancer Metastasis Rev. 2010, 29, 553–568. [Google Scholar] [CrossRef]
- Botella, M.Á.; Hernández, V.; Mestre, T.; Hellín, P.; García-Legaz, M.F.; Rivero, R.M.; Martínez, V.; Fenoll, J.; Flores, P. Bioactive Compounds of Tomato Fruit in Response to Salinity, Heat and Their Combination. Agriculture 2021, 11, 534. [Google Scholar] [CrossRef]
- Slimestad, R.; Verheul, M. Review of flavonoids and other phenolics from fruits of different tomato (Lycopersicon esculentum Mill.) cultivars. J. Sci. Food Agric. 2009, 89, 1255–1270. [Google Scholar] [CrossRef]
- Toscano, S.; Trivellini, A.; Cocetta, G.; Bulgari, R.; Francini, A.; Romano, D.; Ferrante, A. Effect of Preharvest Abiotic Stresses on the Accumulation of Bioactive Compounds in Horticultural Produce. Front. Plant. Sci. 2019, 10, 1212. [Google Scholar] [CrossRef]
- Roy, P.R.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Hossen, M.Z.; Hossain, M.A. Physiological mechanisms of exogenous calcium on alleviating salinity-induced stress in rice (Oryza sativa L.). Physiol. Mol. Biol. Plants. 2019, 253, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Ye, W.; Matsushima, D.; Rhaman, M.S.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, M.S.; Mano, J.; Murata, Y. Reactive carbonyl species function as signal mediators downstream of H2O2 production and regulate [Ca2+]cyt elevation in ABA signal pathway in Arabidopsis guard cells. Plant Cell Physiol. 2019, 60, 1146–1159. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Jamra, G.; Singh, U.M.; Sood, S.; Kumar, A. Calcium Biofortification: Three Pronged Molecular Approaches for Dissecting Complex Trait of Calcium Nutrition in Finger Millet (Eleusine coracana) for Devising Strategies of Enrichment of Food Crops. Front. Plant Sci. 2017, 7, 2028. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Meng, Q.; Brestic, M.; Olsovska, K.; Yang, X. Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. J. Plant Physiol. 2011, 168, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Manaa, A.; Faurobert, M.; Valot, B.; Bouchet, J.-P.; Grasselly, D.; Causse, M. Effect of salinity and calcium on tomato fruit proteome. OMICS 2013, 17, 338–352. [Google Scholar] [CrossRef]
- Tanveer, K.; Gilani, S.; Hussain, Z.; Ishaq, R.; Adeel, M.; Ilyas, N. Effect of salt stress on tomato plant and the role of calcium. J. Plant Nutr. 2020, 43, 28–35. [Google Scholar] [CrossRef]
- Henry, E.E.; Sossa, E.; Noumavo, A.P.; Amadji, G.; Baba-Moussa, L.; Gandonou, C.B. Ions and Organic Solutes as Implicated in the Ameliorative Effect of Exogenous Application of Calcium on Salt Stressed Tomato (Lycopersicon esculentum Mill.). Plants. Int. J. Plant Soil Sci. 2021, 33, 200–212. [Google Scholar] [CrossRef]
- Azad, A.K.; Ohab, M.A.; Shaha, M.G.; Nacha, J.; Rahman, M.L.; Rahman, M.H.H.; Al-amin, M. (Eds.) KRISHI PROJUKTI HATBOI (Handbook on Agro-Technology), 8th ed.; Bangladesh Agricultural Research Institute: Gazipur, Bangladesh, 2019. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Station 1950, 347, 32. [Google Scholar]
- Yamasaki, S.; Dillenburg, L.R. Measurements of leaf relative water content in Araucaria angustifolia. Rev. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Madhava Rao, K.V.; Sresty, T.V. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Aebi, H. Methods Enzymol; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press, Inc.: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium supplementation improves Na+/K+ ratio, antioxidant defense and glyoxalase systems in salt-stressed rice seedlings. Front. Plant Sci. 2016, 7, 609. [Google Scholar] [CrossRef]
- Hostettler, C.; Kölling, K.; Santelia, D.; Streb, S.; Kötting, O.; Zeeman, S.C. Analysis of starch metabolism in chloroplasts. Methods Mol. Biol. 2011, 775, 387–410. [Google Scholar] [CrossRef]
- Thalmann, M.; Pazmino, D.; Seung, D.; Horrer, D.; Nigro, A.; Meier, T.; Kolling, K.; Pfeifhofer, H.W.; Zeeman, C.S.; Santelia, D. Regulation of leaf starch degradation by abscisic acid is important for osmotic stress tolerance in plants. Plant Cell 2016, 28, 1860–1878. [Google Scholar] [CrossRef]
- Nagata, M.; Dan, K.; Yamashita, I. Simple methods for simultaneous determination of chlorophyll and carotenoiods in tomato. J. Jpn. Soc. Hort. Sci. 1992, 612, 685–687. [Google Scholar]
- Perrin, P.W.; Gaye, M.M. Effects of simulated retail display and overnight storage treatments on quality maintenance of fresh broccoli. J. Food Sci. 1986, 51, 146–149. [Google Scholar] [CrossRef]
- Iqbal, M.; Niamatullah, M.; Mohammad, D. Effect of different doses of nitrogen on economical yield and physic-chemical characteristics of apple fruits. J. Anil. Plant Sci. 2012, 221, 165–168. [Google Scholar]
- Singh, P.K.; Gautam, S. Role of salicylic acid on physiological and biochemical mechanism of salinity stress tolerance in plants. Acta Physiol. Plant. 2013, 358, 2345–2353. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol. Plant. 2015, 37, 71. [Google Scholar] [CrossRef]
- Islam, M.M.; Faruqe, M.H.; Rana, M.S.; Akter, M.; Karim, M.A. Screening of rice (Oryza sativa L.) genotypes at reproductive stage for their tolerance to salinity. Agriculturists 2018, 16, 55–67. [Google Scholar] [CrossRef]
- Yousuf, P.Y.; Ahmad, A.; Hemant Ganie, A.; Aref, I.M.; Iqbal, M. Potassium and calcium application ameliorates growth and oxidative homeostasis in salt-stressed indian mustard (Brassica juncea) plants. Pak. J. Bot. 2015, 47, 1629–1639. [Google Scholar]
- Parvin, K.; Ahamed, K.U.; Islam, M.M.; Haque, M.N.; Hore, P.K.; Siddik, M.A.; Roy, I. Reproductive behavior of tomato plant under saline condition with exogenous application of calcium. Mid. East J. Sci. Res. 2015, 23, 2920–2926. [Google Scholar] [CrossRef]
- Ahmad, P.; Sarwat, M.; Bhat, N.A.; Wani, M.R.; Kazi, A.G.; Tran, L.S.P. Alleviation of cadmium toxicity in Brassica juncea L. (Czern. & Coss.) by calcium application involves various physiological and biochemical strategies. PLoS ONE 2015, 10, e0114571. [Google Scholar] [CrossRef]
- Khanbabaloo, N.; Seyed Hajizadeh, H.; Behtash, F. Effects of salinity on taste quality and biochemical traits of four tomato varieties Solanum lycopersicum grown under hydroponic conditions. J. Hort. Post. Res. 2018, 1, 15–26. [Google Scholar]
- Sun, K.; Hunt, K.; Hauser, B.A. Ovule abortion in Arabidopsis triggered by stress. Plant Physiol. 2004, 135, 2358–2367. [Google Scholar] [CrossRef]
- Chamekh, Z.; Ayed, S.; Sahli, A.; Ayadi, S.; Hammemi, Z. Effect of salt stress on the flag leaf area and yield components in twenty five durum wheat genotypes Triticum turgidum ssp. durum. J. New Sci. 2014, 6, 15–23. [Google Scholar]
- Sangtarashani, E.S.; Tabatabaei, S.J.; Bolandnazar, S. Yield, Photosynthetic efficiency and Quality parameters of Cherry tomato as affected by Ca2+ and K+ under NaCl salinity. Intl. J. Agri. Crop Sci. 2013, 5, 1280–1288. [Google Scholar]
- Park, S.; Cheng, N.H.; Pittman, J.K.; Yoo, K.S.; Park, J.; Smith, R.H. Increased calcium levels and prolonged shelf-life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol. 2005, 139, 1194–1206. [Google Scholar] [CrossRef]
- Moles, T.M.; de Brito Francisco, R.; Mariotti, L.; Pompeiano, A.; Lupini, A.; Incrocci, L.; Carmassi, G.; Scartazza, A.; Pistelli, L.; Guglielminetti, L.; et al. Salinity in autumn-winter season and fruit quality of tomato landraces. Front. Plant Sci. 2019, 10, 1078. [Google Scholar] [CrossRef]
- Agurla, S.; Gahir, S.; Munemasa, S.; Murata, Y.; Raghavendra, A.S. Mechanism of stomatal closure in plants exposed to drought and cold stress. In Survival Strategies in Extreme Cold and Desiccation; Iwaya-Inoue, M., Ed.; Springer: Singapore, 2018; pp. 215–232. [Google Scholar]
- Demidchik, V.; Davenport, R.J.; Tester, M. Nonselective cation channels in plants. Annu. Rev. Plant Biol. 2002, 53, 67–107. [Google Scholar] [CrossRef]
- Wu, G.Q.; Wang, S.M. Calcium regulates K+/Na+ homeostasis in rice (Oryza sativa L.) under saline conditions. Plant Soil Environ. 2012, 58, 121–127. [Google Scholar] [CrossRef]
- Gu, M.F.; Li, N.; Long, X.H.; Brestic, M.; Shao, H.B.; Li, J.B.; Mbraki, S. Accumulation capacity of ions in cabbage (Brassica oleracea L.) supplied with sea water. Plant Soil Environ. 2016, 62, 314–320. [Google Scholar] [CrossRef]
- Silva, E.N.; Ribeiro, R.V.; Ferreira-Silva, S.L.; Vie’gas, R.A.; Silveira, J.A. Salt stress induced damages on the photosynthesis of physic nut young plants. Sci. Agric. 2011, 68, 62–68. [Google Scholar] [CrossRef]
- Hayat, S.; Hasan, S.A.; Yusuf, M.; Hayat, Q.; Ahmad, A. Effect of 28-homobrassinolide on photosynthesis, fluorescence and antioxidant system in the presence or absence of salinity and temperature in Vigna radiata. Environ. Exp. Bot. 2010, 69, 105–112. [Google Scholar] [CrossRef]
- Lechowski, Z.; Białczyk, J. Calcium mediated cytokinin action on chlorophyll synthesis in isolated embryo of Scots pine. Biol. Plant. 1993, 35, 53. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycinebetaine and ethylene in mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.H.; Murata, N. Glycinebetaine: An effective protectant against abiotic stress in plants. Trends Plant Sci. 2008, 13, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.S.; Jogeswar, G.; Rasineni, G.K.; Maheswari, M.; Reddy, A.R.; Varshney, R.K.; Kishor, P.K. Proline over-accumulation alleviates salt stress and protects photosynthetic and antioxidant enzyme activities in transgenic sorghum [Sorghum bicolor (L.) Moench]. Plant Physiol. Biochem. 2015, 94, 104–113. [Google Scholar] [CrossRef]
- Ohnishi, N.; Murata, N. Glycinebetaine Counteracts the Inhibitory Effects of Salt Stress on the Degradation and Synthesis of D1 Protein during Photoinhibition in Synechococcus sp. PCC 7942. Plant Physiol. 2006, 141, 758–765. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline Metabolism and Its Implications for Plant-Environment Interaction. The Arabidopsis Book 2010, 8, e014.0. [Google Scholar] [CrossRef]
- Manai, J.; Gouia, H.; Corpas, F.J. Redox and nitric oxide homeostasis are affected in tomato Solanum lycopersicum roots under salinity-induced oxidative stress. J. Plant Physiol. 2014, 171, 1028–1035. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A Central Regulator of Plant Growth and Development. Plant Cell. 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- Farooq, M.A.; Gill, R.A.; Islam, F.; Ali, B.; Liu, H.; Xu, J.; He, S.; Zhou, W. Methyl jasmonate regulates antioxidant defense and suppresses arsenic uptake in Brassica napus L. Front. Plant Sci. 2016, 7, 468. [Google Scholar] [CrossRef]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; Ghassemi-Golezani, K. Changes in antioxidant enzymes activity and plant performance by salinity stress and zinc application in soybean (Glycine max L.). Plant Omics J. 2012, 5, 60–67. [Google Scholar]
- Jannat, R.; Uraji, M.; Morofuji, M.; Islam, M.M.; Bloom, R.E.; Nakamura, Y.; McClung, C.R.; Schroeder, J.I.; Mori, I.C.; Murata, Y. Roles of intracellular hydrogen peroxide accumulation in abscisic acid signaling in Arabidopsis guard cells. J. Plant Physiol. 2011, 16816, 1919–1926. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasja¨rvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Hossain, M.A.; Fujita, M. Hydrogen peroxide priming stimulates drought tolerance in mustard Brassica juncea L. seedlings. Plant Gene Trait. 2013, 4, 109–123. [Google Scholar] [CrossRef]
- De Leon, T.B.; Linscombe, S.; Gregorio, G.; Subudhi, P.K. Genetic variation in Southern USA rice genotypes for seedling salinity tolerance. Front. Plant Sci. 2015, 6, 374. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M. Influence of exogenous application of brassinosteroid on growth and mineral nutrients of wheat (Triticum aestivum L.) under saline conditions. Pak. J. Bot. 2007, 39, 513–522. [Google Scholar]
- Khayyat, M.; Tafazoli, E.; Eshghi, S.; Rahemi, M.; Rajaee, S. Salinity, supplementary calcium and potassium effects on fruit yield and quality of strawberry Fragaria ananassa Duch. Am. Eurasian J. Agric. Environ. Sci. 2007, 2, 539–544. [Google Scholar]
- Randome, I.; Basu, S.; Pereira, A. Effect of different stress treatments on mature green tomatoes Solanum lycopersicum to enhance fruit quality. Afr. J Food, Agric. Nutr. Dev. 2017, 174, 12547–12556. [Google Scholar] [CrossRef]
- Ullah, N.; Basit, A.; Ahmad, I.; Ullah, I.; Shah, S.T.; Mohamed, H.I.; Javed, S. Mitigation the adverse effect of salinity stress on the performance of the tomato crop by exogenous application of chitosan. Bull. Natl. Res. Cent. 2020, 44, 181. [Google Scholar] [CrossRef]
- Saito, T.; Fukuda, N.; Matsukura, C.; Nishimura, S. Effects of Salinity on Distribution of Photosynthates and Carbohydrate Metabolism in Tomato Grown using Nutrient Film Technique. J. Jpn. Soc. Hortic. Sci. 2009, 78, 90–96. [Google Scholar] [CrossRef]
- Naeem, M.; Basit, A.; Ahmad, I.; Mohamed, H.I.; Wasila, H. Effect of salicylic acid and salinity stress on the performance of tomato plants. Gesunde Pflanz. 2020, 724, 393–402. [Google Scholar] [CrossRef]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing adverse effects of pb on maize plants by combined treatment with jasmonic, salicylic acids and proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Ehret, D.L.; Usher, K.; Helmer, T.; Block, G.; Steinke, D.; Frey, B.; Kuang, T.; Diarra, M. Tomato Fruit Antioxidants in Relation to Salinity and Greenhouse Climate. J. Agric. Food Chem. 2013, 61, 1138–1145. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Treatment | Plant Height (cm) | Leaves per plant−1 | Shoot Dry Weight (g per plant−1) | Root Dry Weight (g per plant−1) | |
|---|---|---|---|---|---|---|
| BARI Tomato-2 | Control | T1 | 103 ± 1.97 c | 55 ± 2.08 bc | 27.7 ± 0.21 d | 1.5 ± 0.09 cd |
| 5 mM Ca | T2 | 107 ± 2.89 ab | 56 ± 1.53 b | 29.5 ± 0.70 b | 1.6 ± 0.10 abc | |
| 10 mM Ca | T3 | 109 ± 3.06 a | 59 ± 2.08 a | 31.7 ± 0.63 a | 1.7 ± 0.13 a | |
| 15 mM Ca | T4 | 103 ± 2.52 bc | 55 ± 1.73 bc | 29.8 ± 0.60 b | 1.7 ± 0.17 ab | |
| 12 dS m−1 NaCl | T5 | 72 ± 2.45 g | 34 ± 2.52 g | 19.9 ± 0.65 i | 0.9 ± 0.07 g | |
| T5 + 5 mM Ca | T6 | 77 ± 3.09 f | 37 ± 1.53 ef | 22.7 ± 0.25 g | 1.0 ± 0.12 f | |
| T5 + 10 mM Ca | T7 | 81 ± 2.48 e | 38 ± 1.53 e | 25.8 ± 0.15 f | 1.3 ± 0.09 e | |
| T5 + 15 mM Ca | T8 | 75 ± 1.79 f | 36 ± 1.53 fg | 21.4 ± 0.65 h | 1.1 ± 0.08 f | |
| Binatomato-5 | Control | T1 | 98 ± 2.99 d | 51 ± 2.08 d | 26.7 ± 0.07 e | 1.5 ± 0.09 d |
| 5 mM Ca | T2 | 102 ± 2.09 c | 55 ± 1.73 bc | 28.4 ± 0.96 cd | 1.5 ± 0.13 cd | |
| 10 mM Ca | T3 | 103 ± 2.03 bc | 56 ± 1.15 b | 29.6 ± 0.89 b | 1.7 ± 0.14 ab | |
| 15 mM Ca | T4 | 100 ± 2.78 cd | 53 ± 1.63 cd | 28.5 ± 0.33 c | 1.6 ± 0.15 bcd | |
| 12 dS m−1 NaCl | T5 | 59 ± 1.66 i | 21 ± 0.72 i | 9.5 ± 0.13 l | 0.5 ± 0.06 i | |
| T5 + 5 mM Ca | T6 | 62 ± 1.67 hi | 24 ± 1.15 h | 11.1 ± 0.51 k | 0.6 ± 0.08 h | |
| T5 + 10 mM Ca | T7 | 65 ± 2.65 h | 25 ± 0.90 h | 13.6 ± 0.56 j | 0.9 ± 0.12 g | |
| T5 + 15 mM Ca | T8 | 61 ± 1.75 i | 23 ± 0.35 hi | 10.3 ± 0.31 k | 0.8 ± 0.10 gh | |
| CV (%) | 2.51 | 3.26 | 2.03 | 6.44 | ||
| LSD(0.05) | 3.61 | 2.31 | 0.77 | 0.133 | ||
| Variety | Treatment | Number of Fruits per Plant−1 | Fruits Mass (kg) per Plant−1 | |
|---|---|---|---|---|
| BARI Tomato-2 | Control | T1 | 28 ± 0.84 f | 1.71 ± 0.19 c |
| 5 mM Ca | T2 | 31 ± 1.03 c | 2.09 ± 0.29 b | |
| 10 mM Ca | T3 | 34 ± 1.06 a | 2.32 ± 0.26 a | |
| 15 mM Ca | T4 | 31 ± 0.97 d | 2.05 ± 0.16 b | |
| 12 dS m−1 NaCl | T5 | 16 ± 0.69 j | 1.29 ± 0.22 f | |
| T5 + 5 mM Ca | T6 | 21 ± 0.61 h | 1.55 ± 0.14 cd | |
| T5 + 10 mM Ca | T7 | 22 ± 0.85 g | 1.58 ± 0.14 cd | |
| T5 + 15 mM Ca | T8 | 19 ± 0.41 i | 1.37 ± 0.15 ef | |
| Binatomato-5 | Control | T1 | 28 ± 0.67 f | 1.53 ± 0.16 de |
| 5 mM Ca | T2 | 30 ± 1.05 e | 1.71 ± 0.12 c | |
| 10 mM Ca | T3 | 32 ± 1.17 b | 1.98 ± 0.12 b | |
| 15 mM Ca | T4 | 30 ± 0.93 e | 1.64 ± 0.21 cd | |
| 12 dS m−1 NaCl | T5 | 8 ± 0.44 n | 0.61 ± 0.18 h | |
| T5 + 5 mM Ca | T6 | 11 ± 0.56 l | 0.84 ± 0.11 g | |
| T5 + 10 mM Ca | T7 | 12 ± 0.41 k | 0.89 ± 0.10 g | |
| T5 + 15 mM Ca | T8 | 10 ± 0.66 m | 0.77 ± 0.06 gh | |
| CV (%) | 1.51 | 7.03 | ||
| LSD(0.05) | 0.57 | 0.175 | ||
| Variety | Treatment | Relative Water Content (%) | |
|---|---|---|---|
| BARI Tomato-2 | Control | T1 | 85 ± 1.80 bcd |
| 5 mM Ca | T2 | 87 ± 2.21 ab | |
| 10 mM Ca | T3 | 88 ± 1.53 a | |
| 15 mM Ca | T4 | 86 ± 2.12 abc | |
| 12 dS m−1 NaCl | T5 | 75 ± 2.14 g | |
| T5 + 5 mM Ca | T6 | 79 ± 1.75 f | |
| T5 + 10 mM Ca | T7 | 82 ± 2.18 e | |
| T5 + 15 mM Ca | T8 | 76 ± 1.45 g | |
| Binatomato-5 | Control | T1 | 84 ± 1.19 d |
| 5 mM Ca | T2 | 85 ± 1.11 bcd | |
| 10 mM Ca | T3 | 86 ± 1.65 bc | |
| 15 mM Ca | T4 | 85 ± 0.66 cd | |
| 12 dS m−1 NaCl | T5 | 65 ± 1.35 j | |
| T5 + 5 mM Ca | T6 | 69 ± 0.58 i | |
| T5 + 10 mM Ca | T7 | 71 ± 0.94 h | |
| T5 + 15 mM Ca | T8 | 65 ± 1.35 j | |
| CV (%) | 1.34 | ||
| LSD(0.05) | 1.76 | ||
| Variety | Treatment | %Na+ | %K+ | Na + /K+ | %Ca2+ | %Mg2+ | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaves | Roots | Leaves | Roots | Leaves | Roots | Leaves | Roots | Leaves | Roots | |||
| BARI Tomato-2 | Control | T1 | 0.26 ± 0.02 gh | 0.51 ± 0.01 ef | 2.73 ± 0.08 b | 1.37 ± 0.02 f | 0.10 ± 0.01 g | 0.37 ± 0.01 h | 2.37 ± 0.02 f | 3.56 ± 0.01 f | 0.79 ± 0.03 efg | 0.53 ± 0.02 bc |
| 5 mM Ca | T2 | 0.23 ± 0.01 ij | 0.42 ± 0.03 hi | 2.77 ± 0.05 b | 1.52 ± 0.01 e | 0.08 ± 0.00 ghi | 0.28 ± 0.02 i | 2.79 ± 0.05 c | 3.91 ± 0.02 c | 0.84 ± 0.03 cd | 0.53 ± 0.09 bc | |
| 10 mM Ca | T3 | 0.22 ± 0.01 j | 0.38 ± 0.01 i | 2.77 ± 0.05 b | 1.63 ± 0.05 c | 0.08 ± 0.00 ghi | 0.24 ± 0.01 ijk | 3.05 ± 0.06 a | 4.31 ± 0.04 a | 0.93 ± 0.02 a | 0.60 ± 0.00 a | |
| 15 mM Ca | T4 | 0.23 ± 0.01 hij | 0.39 ± 0.01 i | 2.76 ± 0.05 b | 1.59 ± 0.04 d | 0.08 ± 0.00 ghi | 0.24 ± 0.01 ij | 2.89 ± 0.02 b | 4.19 ± 0.02 b | 0.89 ± 0.02 ab | 0.56 ± 0.04 ab | |
| 12 dS m−1 NaCl | T5 | 0.56 ± 0.07 bc | 0.68 ± 0.05 c | 2.13 ± 0.06 f | 0.83 ± 0.01 i | 0.26 ± 0.03 b | 0.82 ± 0.07 d | 2.05 ± 0.08 i | 2.89 ± 0.01 k | 0.65 ± 0.01 i | 0.39 ± 0.00 fgh | |
| T5 + 5 mM Ca | T6 | 0.46 ± 0.01 e | 0.61 ± 0.06 d | 2.23 ± 0.02 def | 0.87 ± 0.03 hi | 0.20 ± 0.00 d | 0.71 ± 0.09 e | 2.27 ± 0.05 g | 3.03 ± 0.02 j | 0.75 ± 0.01 gh | 0.44 ± 0.00 ef | |
| T5 + 10 mM Ca | T7 | 0.37 ± 0.01 f | 0.50 ± 0.01 fg | 2.41 ± 0.03 c | 0.97 ± 0.01 g | 0.15 ± 0.01 f | 0.52 ± 0.01 g | 2.50 ± 0.04 e | 3.25 ± 0.02 h | 0.77 ± 0.00 efg | 0.47 ± 0.01 de | |
| T5 + 15 mM Ca | T8 | 0.43 ± 0.01 e | 0.57 ± 0.02 de | 2.33 ± 0.04 cde | 0.89 ± 0.01 h | 0.18 ± 0.00 e | 0.63 ± 0.03 f | 2.10 ± 0.01 hi | 2.91 ± 0.05 k | 0.72 ± 0.00 h | 0.42 ± 0.01 efg | |
| Binatomato-5 | Control | T1 | 0.29 ± 0.00 g | 0.45 ± 0.07 gh | 3.14 ± 0.02 a | 1.53 ± 0.02 e | 0.09 ± 0.00 gh | 0.30 ± 0.04 i | 2.15 ± 0.00 h | 3.11 ± 0.00 i | 0.76 ± 0.05 fgh | 0.50 ± 0.04 cd |
| 5 mM Ca | T2 | 0.25 ± 0.01 hij | 0.37 ± 0.02 ij | 3.18 ± 0.07 a | 1.78 ± 0.04 b | 0.08 ± 0.00 hi | 0.21 ± 0.01 jkl | 2.57 ± 0.06 d | 3.46 ± 0.02 g | 0.81 ± 0.02 def | 0.51 ± 0.00 bcd | |
| 10 mM Ca | T3 | 0.23 ± 0.01 ij | 0.31 ± 0.01 j | 3.18 ± 0.01 a | 1.86 ± 0.02 a | 0.07 ± 0.00 i | 0.17 ± 0.01 l | 2.82 ± 0.08 bc | 3.83 ± 0.01 d | 0.86 ± 0.03 bc | 0.56 ± 0.01 ab | |
| 15 mM Ca | T4 | 0.26 ± 0.01 ghi | 0.32 ± 0.03 j | 3.18 ± 0.02 a | 1.83 ± 0.04 a | 0.08 ± 0.01 ghi | 0.18 ± 0.02 kl | 2.64 ± 0.05 d | 3.68 ± 0.07 e | 0.81 ± 0.03 cde | 0.53 ± 0.01 bc | |
| 12 dS m−1 NaCl | T5 | 0.69 ± 0.02 a | 0.90 ± 0.02 a | 2.10 ± 0.5 f | 0.74 ± 0.01 l | 0.33 ± 0.03 a | 1.23 ± 0.03 a | 1.30 ± 0.02 l | 1.87 ± 0.01 o | 0.44 ± 0.01 k | 0.31 ± 0.00 j | |
| T5 + 5 mM Ca | T6 | 0.57 ± 0.01 b | 0.84 ± 0.02 b | 2.19 ± 0.01 ef | 0.76 ± 0.01 j | 0.26 ± 0.00 b | 1.11 ± 0.03 b | 1.64 ± 0.02 k | 1.98 ± 0.01 n | 0.58 ± 0.00 j | 0.36 ± 0.00 hij | |
| T5 + 10 mM Ca | T7 | 0.50 ± 0.01 d | 0.70 ± 0.05 c | 2.34 ± 0.04 cd | 0.83 ± 0.01 i | 0.21 ± 0.01 d | 0.84 ± 0.05 d | 1.73 ± 0.02 j | 2.37 ± 0.03 l | 0.61 ± 0.00 ij | 0.38 ± 0.01 ghi | |
| T5 + 15 mM Ca | T8 | 0.53 ± 0.03 c | 0.71 ± 0.05 c | 2.23 ± 0.05 def | 0.76 ± 0.00 j | 0.24 ± 0.01 c | 0.94 ± 0.07 c | 1.66 ± 0.02 jk | 2.16 ± 0.03 m | 0.58 ± 0.00 j | 0.33 ± 0.01 ij | |
| CV (%) | 5.67 | 6.34 | 3.32 | 1.94 | 7.20 | 7.19 | 1.98 | 0.89 | 3.87 | 7.06 | ||
| LSD(0.05) | 0.036 | 0.057 | 0.144 | 0.040 | 0.019 | 0.066 | 0.075 | 0.047 | 0.048 | 0.055 | ||
| Variety | Treatment | TSS (%) | Total Sugar (%) | Reducing Sugar (%) | β-Carotene (mg/100 g FW) | Vitamin C (mg/100 g FW) | Juice pH | |
|---|---|---|---|---|---|---|---|---|
| BARI Tomato-2 | Control | T1 | 5.57 ± 0.55 gh | 3.84 ± 0.13 f | 2.76 ± 0.19 bc | 0.181 ± 0.00 hi | 18.00 ± 0.44 i | 4.50 ± 0.33 |
| 5 mM Ca | T2 | 5.62 ± 0.55 gh | 3.85 ± 0.13 f | 2.78 ± 0.15 bc | 0.191 ± 0.01 fg | 19.78 ± 0.38 g | 4.50 ± 0.55 | |
| 10 mM Ca | T3 | 5.75 ± 0.55 fg | 3.95 ± 0.13 e | 2.86 ± 0.15 b | 0.196 ± 0.01 f | 20.03 ± 0.11 fg | 4.55 ± 0.55 | |
| 15 mM Ca | T4 | 5.50 ± 0.51 h | 3.72 ± 0.08 g | 2.69 ± 0.05 bcd | 0.184 ± 0.01 gh | 18.71 ± 0.09 h | 4.53 ± 0.55 | |
| 12 dS m−1 NaCl | T5 | 7.21 ± 0.55 c | 4.61 ± 0.11 c | 3.18 ± 0.20 a | 0.251 ± 0.01 b | 22.15 ± 0.54 bc | 4.37 ± 0.23 | |
| T5 + 5 mM Ca | T6 | 7.51 ± 0.55 b | 4.74 ± 0.07 b | 3.25 ± 0.20 a | 0.260 ± 0.01 b | 22.51 ± 0.14 ab | 4.48 ± 0.07 | |
| T5 + 10 mM Ca | T7 | 7.78 ± 0.25 a | 5.01 ± 0.13 a | 3.41 ± 0.11 a | 0.274 ± 0.01 a | 22.89 ± 0.19 a | 4.54 ± 0.08 | |
| T5 + 15 mM Ca | T8 | 7.05 ± 0.27 c | 4.46 ± 0.07 d | 2.80 ± 0.20 b | 0.233 ± 0.01 c | 21.86 ± 0.22 c | 4.40 ± 0.55 | |
| Binatomato-5 | Control | T1 | 4.90 ± 0.51 i | 3.21 ± 0.12 k | 2.45 ± 0.19 def | 0.165 ± 0.00 j | 17.90 ± 0.19 i | 4.46 ± 0.08 |
| 5 mM Ca | T2 | 4.94 ± 0.16 i | 3.25 ± 0.08 k | 2.47 ± 0.11 def | 0.169 ± 0.00 j | 19.03 ± 0.11 g | 4.48 ± 0.23 | |
| 10 mM Ca | T3 | 5.01 ± 0.26 i | 3.32 ± 0.6 j | 2.51 ± 0.19 cdef | 0.173 ± 0.00 ij | 19.23 ± 0.23 g | 4.54 ± 0.21 | |
| 15 mM Ca | T4 | 4.90 ± 0.15 i | 3.07 ± 0.12 l | 2.38 ± 0.18 ef | 0.166 ± 0.00 j | 18.08 ± 0.09 h | 4.53 ± 0.22 | |
| 12 dS m−1 NaCl | T5 | 5.95 ± 0.33 ef | 3.58 ± 0.11 i | 2.65 ± 0.09 bcde | 0.206 ± 0.00 e | 21.05 ± 0.13 d | 4.22 ± 0.09 | |
| T5 + 5 mM Ca | T6 | 6.17 ± 0.25 de | 3.67 ± 0.13 h | 2.71 ± 0.11 bcd | 0.211 ± 0.00 de | 21.44 ± 0.19 cd | 4.30 ± 0.17 | |
| T5 + 10 mM Ca | T7 | 6.36 ± 0.33 d | 3.88 ± 0.15 f | 2.83 ± 0.17 b | 0.218 ± 0.01 d | 21.71 ± 0.22 c | 4.47 ± 0.11 | |
| T5 + 15 mM Ca | T8 | 6.01 ± 0.33 e | 3.32 ± 0.008 j | 2.27 ± 0.09 f | 0.187 ± 0.00 fgh | 20.01 ± 0.13 fg | 4.31 ± 0.08 | |
| CV (%) | 2.34 | 2.80 | 6.12 | 2.68 | 1.64 | 5.32 | ||
| LSD(0.05) | 0.234 | 0.051 | 0.281 | 0.009 | 0.545 | 0.394 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.M.; Jahan, K.; Sen, A.; Urmi, T.A.; Haque, M.M.; Ali, H.M.; Siddiqui, M.H.; Murata, Y. Exogenous Application of Calcium Ameliorates Salinity Stress Tolerance of Tomato (Solanum lycopersicum L.) and Enhances Fruit Quality. Antioxidants 2023, 12, 558. https://doi.org/10.3390/antiox12030558
Islam MM, Jahan K, Sen A, Urmi TA, Haque MM, Ali HM, Siddiqui MH, Murata Y. Exogenous Application of Calcium Ameliorates Salinity Stress Tolerance of Tomato (Solanum lycopersicum L.) and Enhances Fruit Quality. Antioxidants. 2023; 12(3):558. https://doi.org/10.3390/antiox12030558
Chicago/Turabian StyleIslam, Md. Moshiul, Khurshida Jahan, Arpita Sen, Tahmina Akter Urmi, M. Moynul Haque, Hayssam M. Ali, Manzer H. Siddiqui, and Yoshiyuki Murata. 2023. "Exogenous Application of Calcium Ameliorates Salinity Stress Tolerance of Tomato (Solanum lycopersicum L.) and Enhances Fruit Quality" Antioxidants 12, no. 3: 558. https://doi.org/10.3390/antiox12030558
APA StyleIslam, M. M., Jahan, K., Sen, A., Urmi, T. A., Haque, M. M., Ali, H. M., Siddiqui, M. H., & Murata, Y. (2023). Exogenous Application of Calcium Ameliorates Salinity Stress Tolerance of Tomato (Solanum lycopersicum L.) and Enhances Fruit Quality. Antioxidants, 12(3), 558. https://doi.org/10.3390/antiox12030558