Author Contributions
Conceptualization, T.P. and P.M.K.; methodology, T.P., P.M.K. and A.K.; software, T.P., P.M.K. and A.K.; validation, T.P., P.M.K. and Ł.R.; formal analysis, T.P., P.M.K. and A.L.-A.; investigation, T.P., P.M.K. and A.K.; resources, T.P., P.M.K. and Ł.T.; data curation, T.P., P.M.K., Ł.T. and A.L.-A.; writing—original draft preparation, T.P., P.M.K., Ł.T., Ł.R. and T.A.; writing—review and editing, T.P., P.M.K., Ł.T., Ł.R., T.A. and W.C.; visualization, T.P., P.M.K. and A.K; supervision, T.P., P.M.K. and T.A.; project administration, T.P., P.M.K. and W.C.; funding acquisition, T.P. and P.M.K. All authors have read and agreed to the published version of the manuscript.
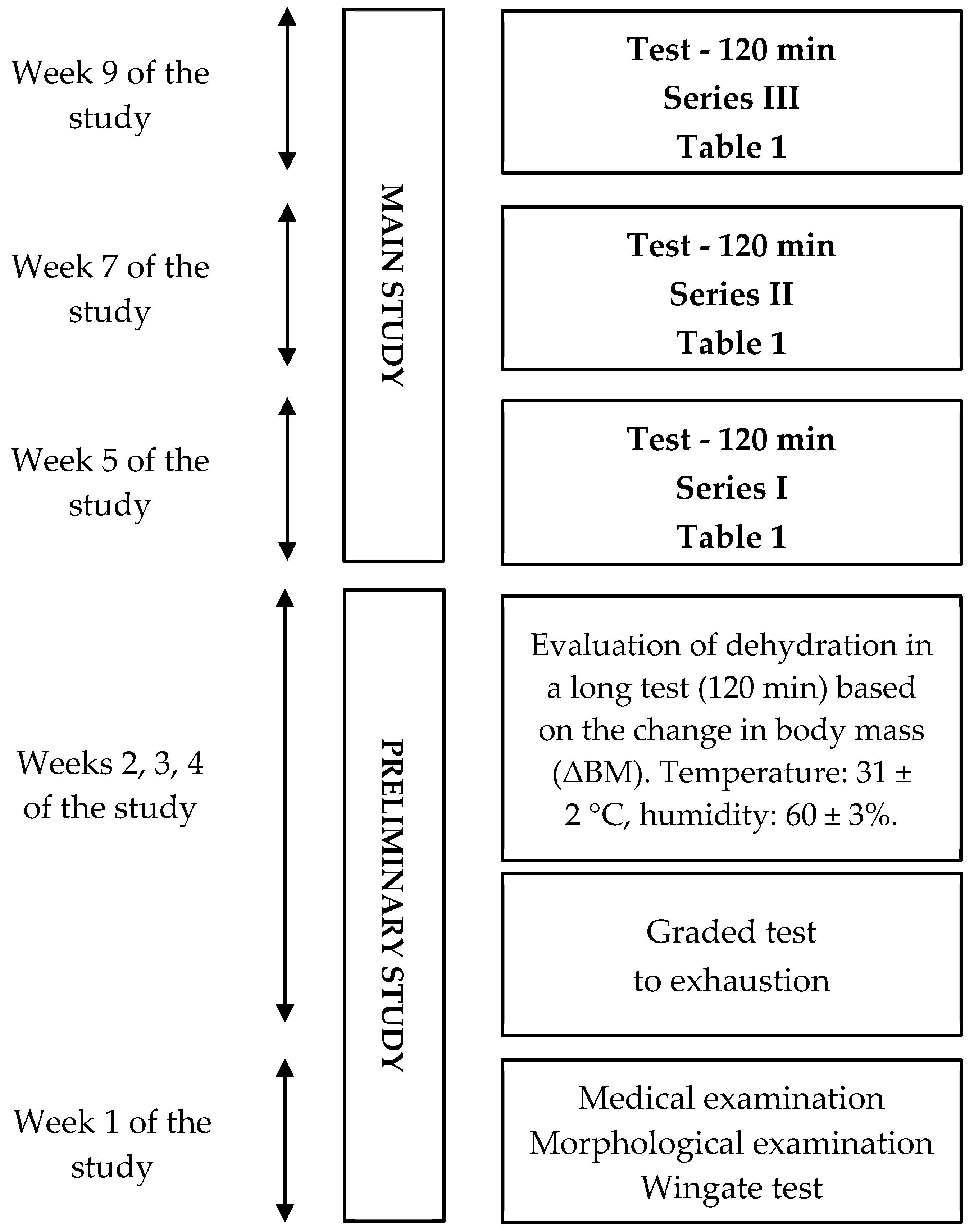
Figure 1.
General experimental design.
Figure 1.
General experimental design.
Figure 2.
Diagram of one series of tests during the main part of the study, including measurement points WBC—white blood cells, tac/toc-pro-oxidant/antioxidant status, LDH—lactate dehydrogenase, Mb—myoglobin, IL−1β—interleukin 1β, CRP-C—reactive protein.
Figure 2.
Diagram of one series of tests during the main part of the study, including measurement points WBC—white blood cells, tac/toc-pro-oxidant/antioxidant status, LDH—lactate dehydrogenase, Mb—myoglobin, IL−1β—interleukin 1β, CRP-C—reactive protein.
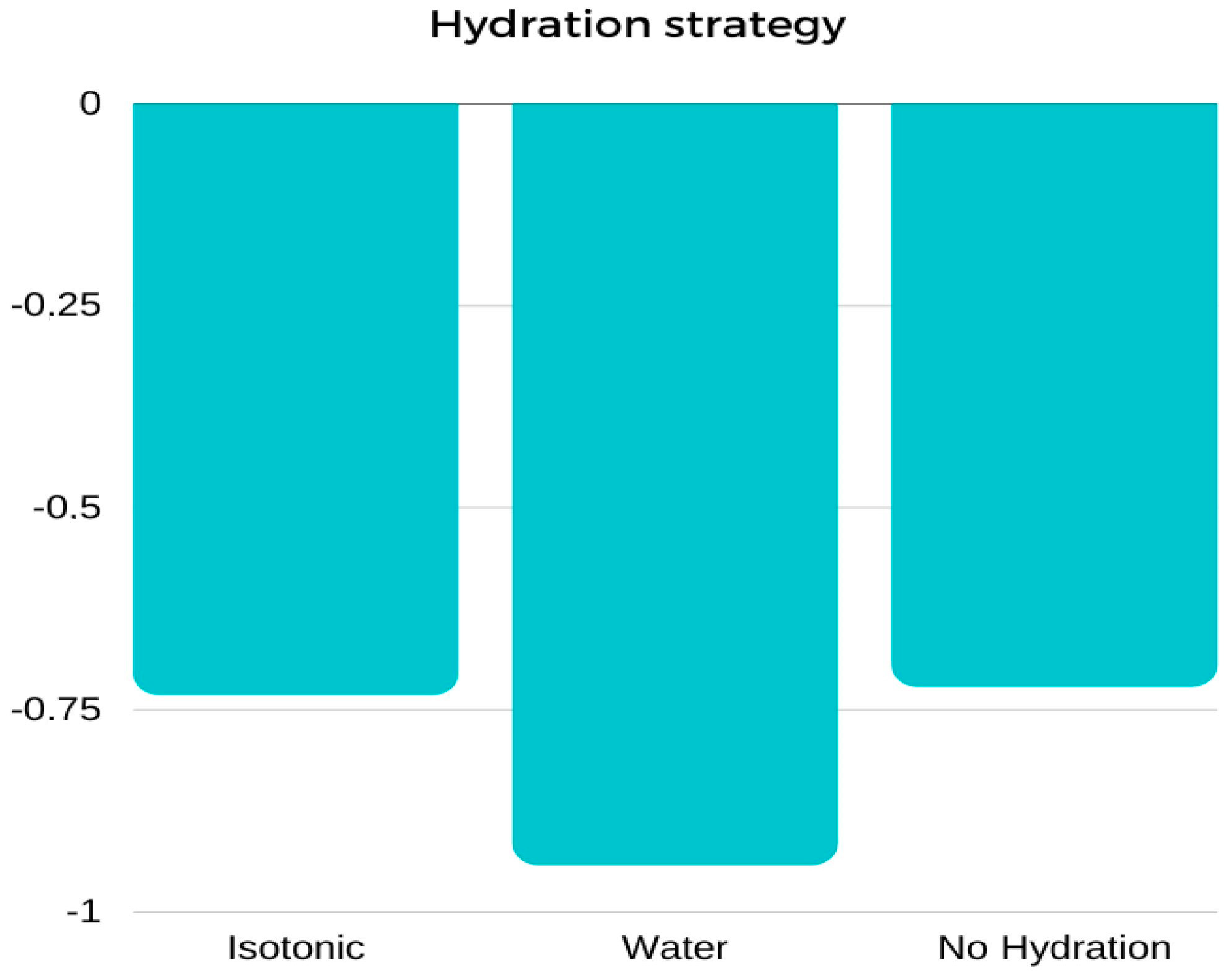
Figure 3.
Mean values of body weight change (∆BM) [kg] between pre-test and post-test results performed using three different hydration strategies: marginal means.
Figure 3.
Mean values of body weight change (∆BM) [kg] between pre-test and post-test results performed using three different hydration strategies: marginal means.
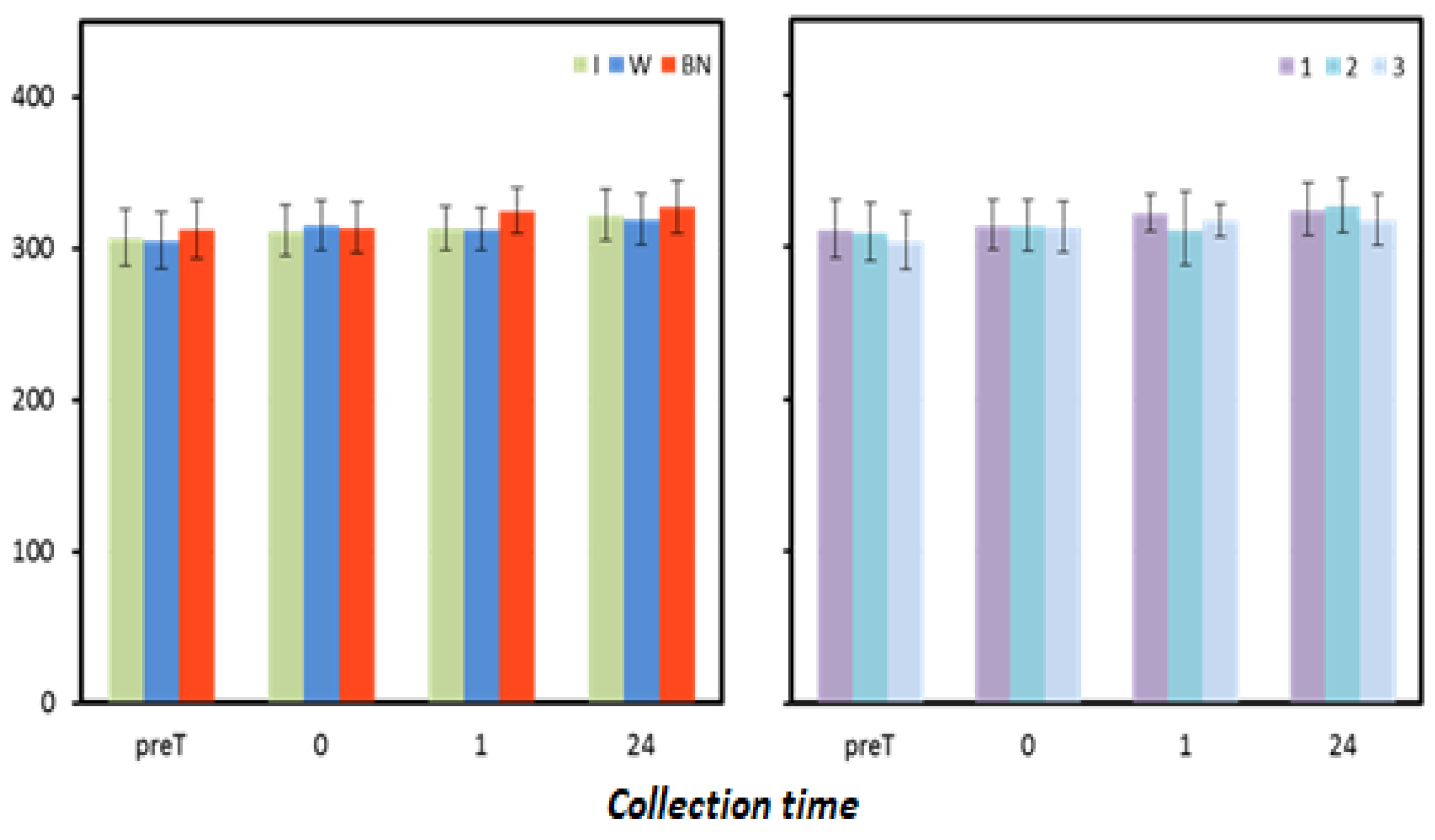
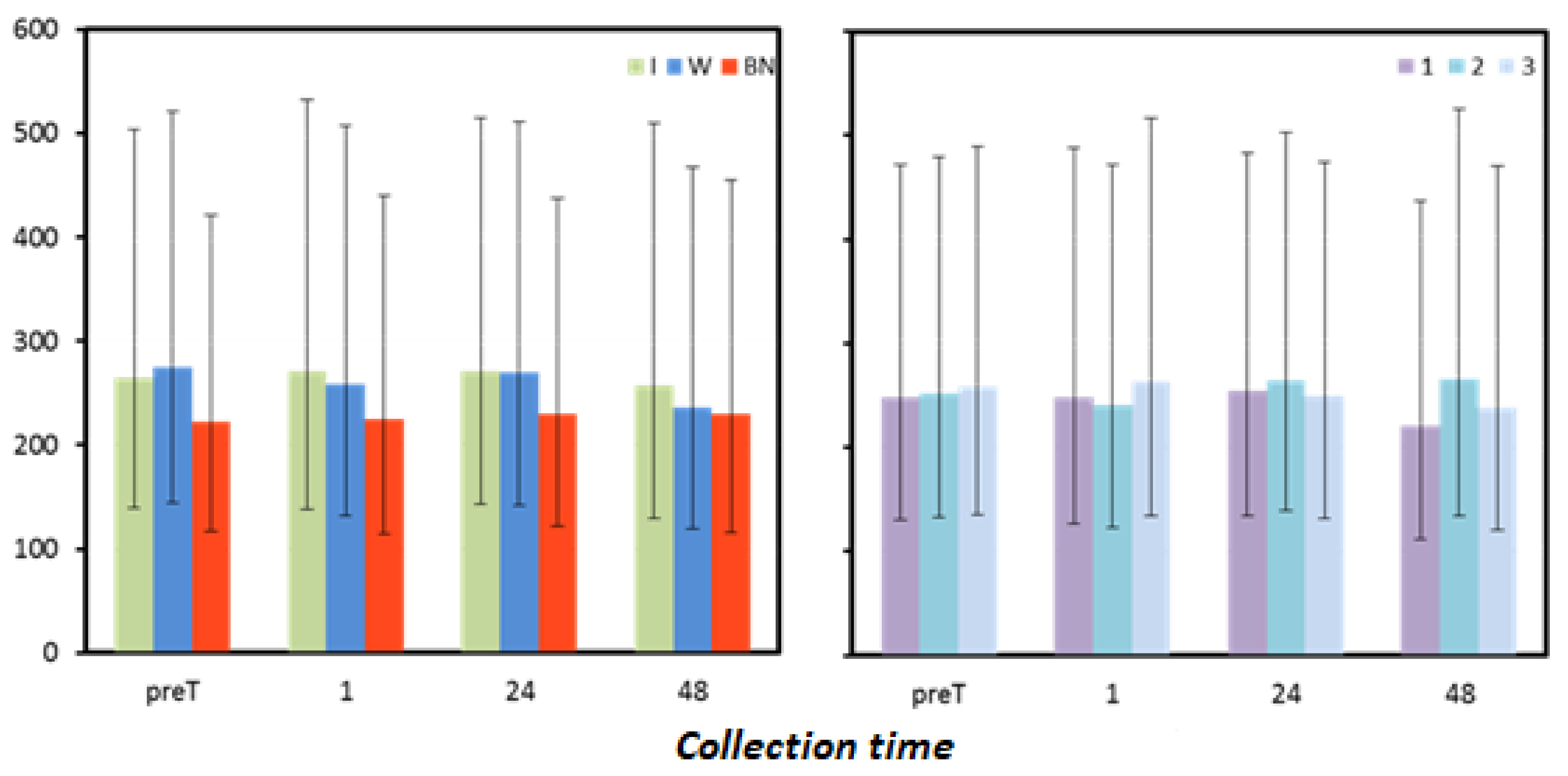
Figure 4.
Oxidative status. Mean values estimated by the method of least squares(µmoL·L−1) (I—isotonic; W—water; BN—none) (immediately preT, postT, 1, 24, and 48 h) taken in 12 men in three consecutive tests (1, 2, 3).
Figure 4.
Oxidative status. Mean values estimated by the method of least squares(µmoL·L−1) (I—isotonic; W—water; BN—none) (immediately preT, postT, 1, 24, and 48 h) taken in 12 men in three consecutive tests (1, 2, 3).
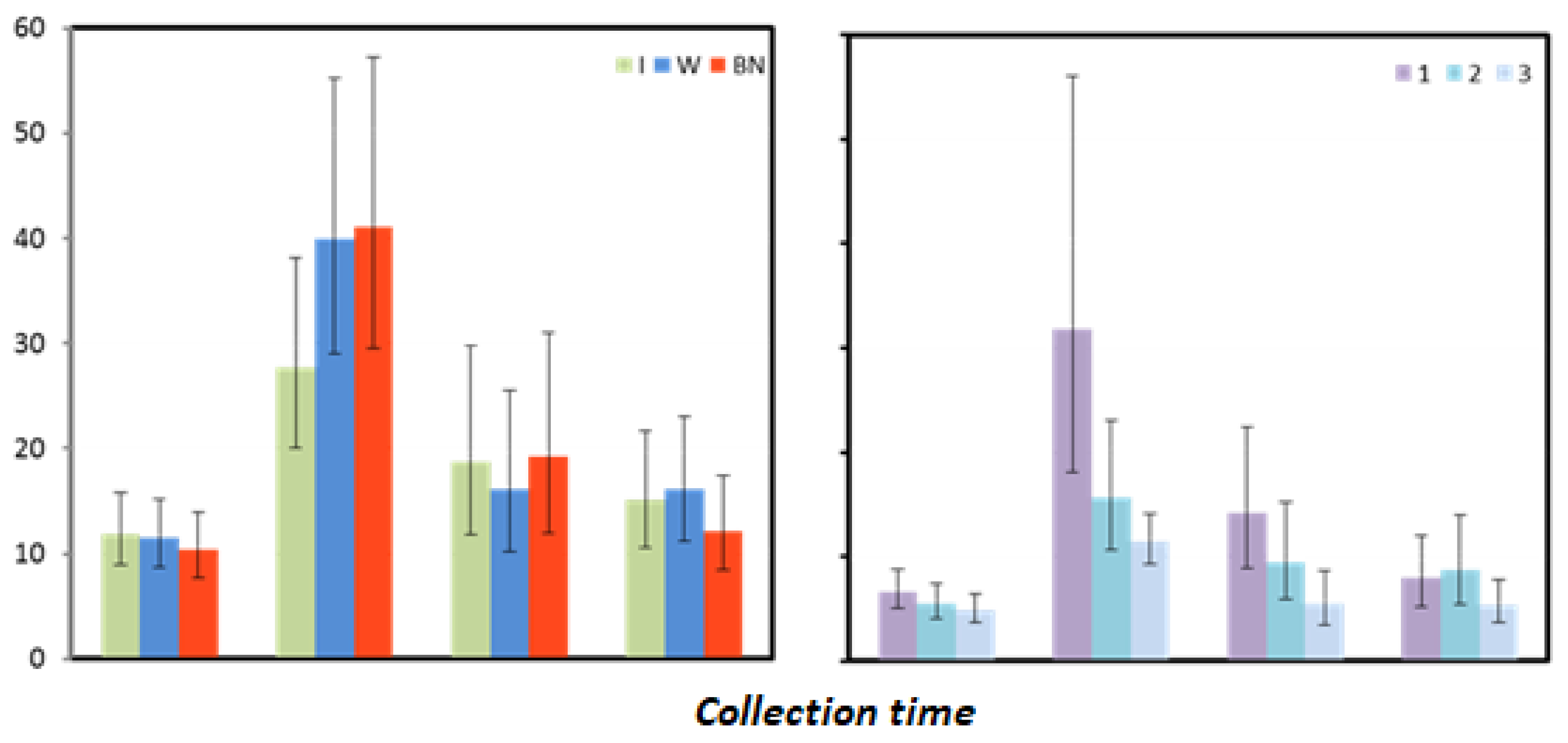
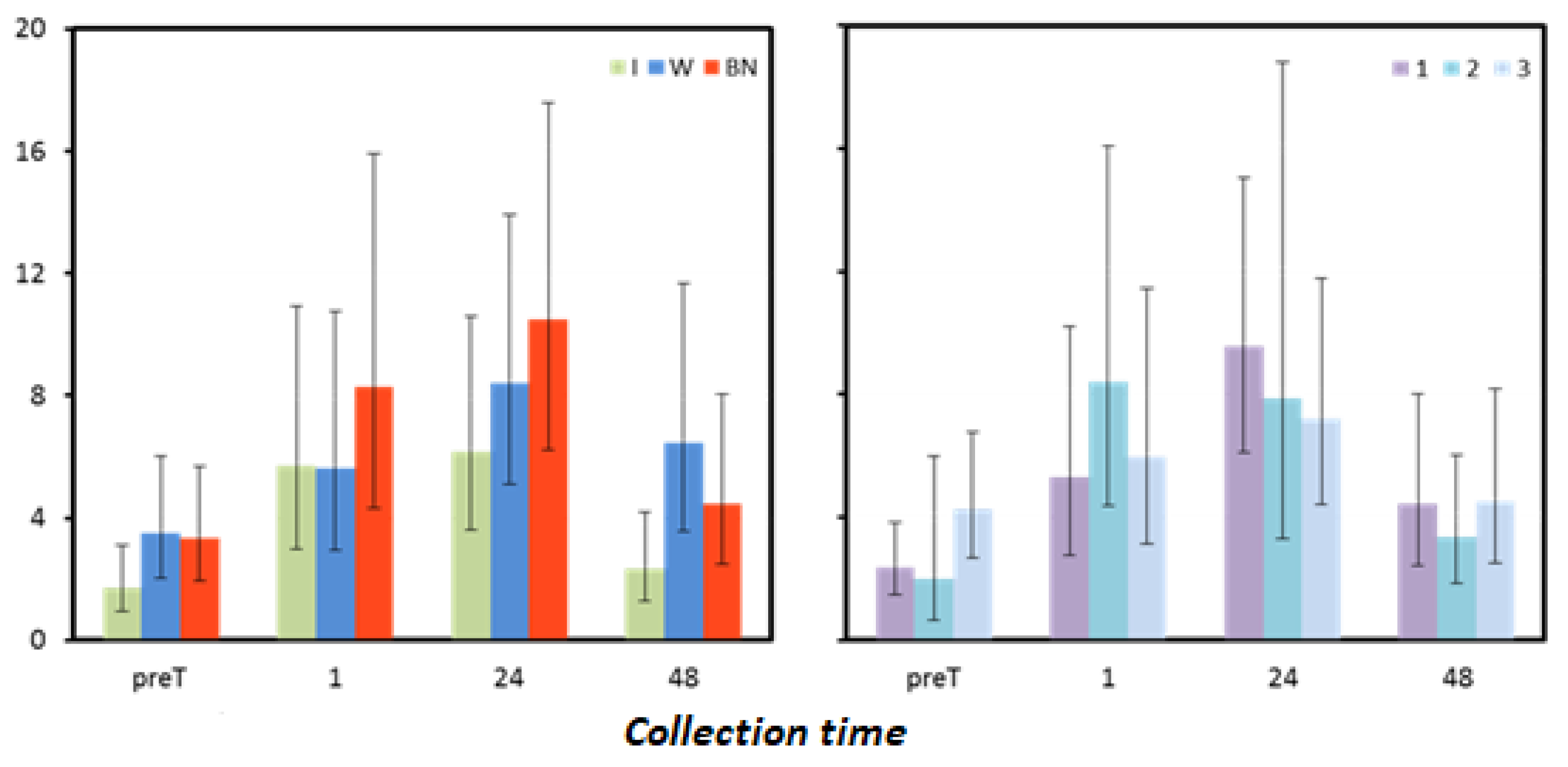
Figure 5.
Myoglobin mean values estimated via the least squares method together with upper and lower confidence intervals for four measurements using three rehydration strategies [ng·mL−1].
Figure 5.
Myoglobin mean values estimated via the least squares method together with upper and lower confidence intervals for four measurements using three rehydration strategies [ng·mL−1].
Figure 6.
Lactate dehydrogenase mean values estimated via the least squares method with upper and lower confidence intervals for four measurements [U.mL−1].
Figure 6.
Lactate dehydrogenase mean values estimated via the least squares method with upper and lower confidence intervals for four measurements [U.mL−1].
Figure 7.
Interleukin mean values estimated via the least squares method with upper and lower confidence intervals for four measurements [pg·mL−1].
Figure 7.
Interleukin mean values estimated via the least squares method with upper and lower confidence intervals for four measurements [pg·mL−1].
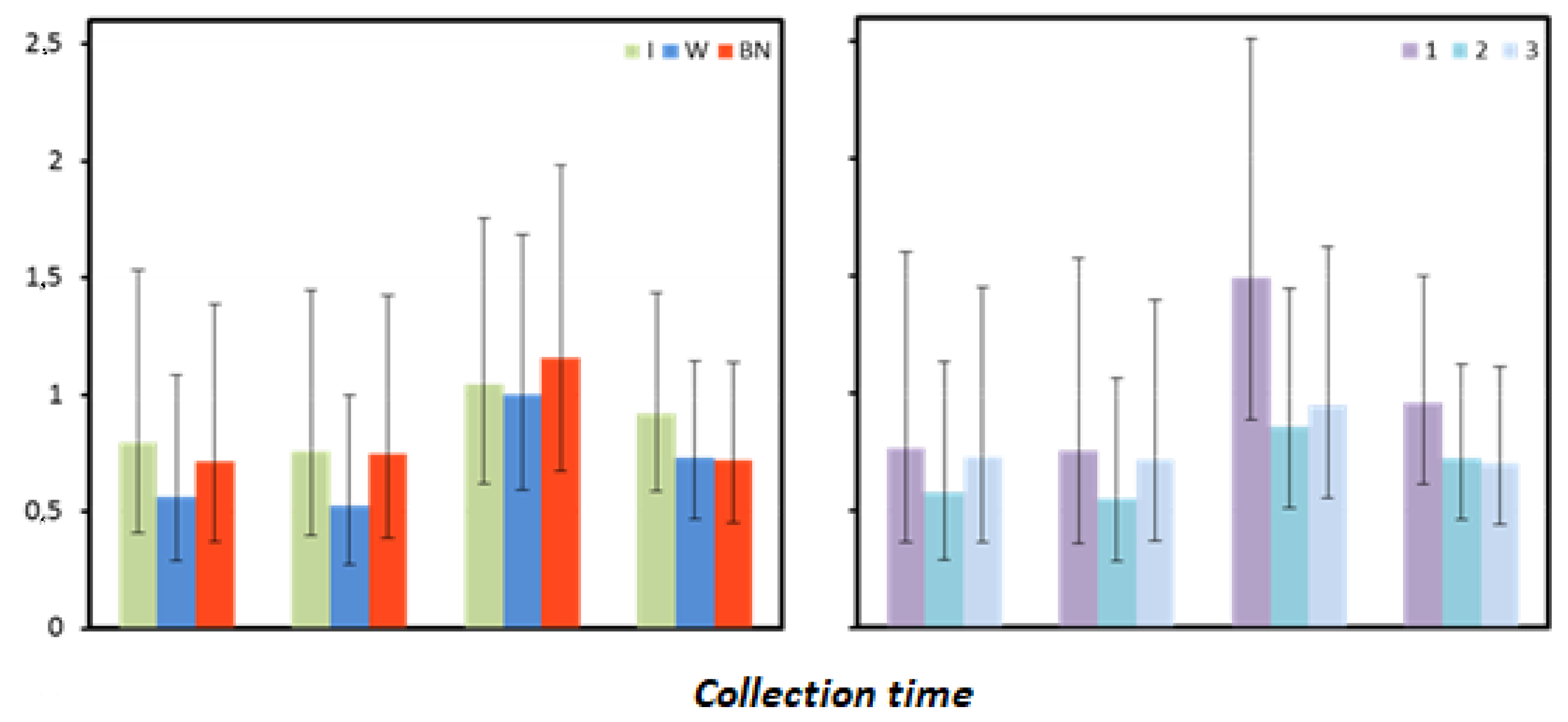
Figure 8.
C-reactive protein mean values estimated by the least squares method with upper and lower confidence intervals for four measurements [mg·mL−1].
Figure 8.
C-reactive protein mean values estimated by the least squares method with upper and lower confidence intervals for four measurements [mg·mL−1].
Table 1.
Cross-over experimental design, in which tests are performed on six groups of two people at 1-week intervals under three hydration conditions (I—isotonic drinks, W—water, NH—no hydration), with a balanced number of interventions in each of the six possible sequences.
Table 1.
Cross-over experimental design, in which tests are performed on six groups of two people at 1-week intervals under three hydration conditions (I—isotonic drinks, W—water, NH—no hydration), with a balanced number of interventions in each of the six possible sequences.
| Group | Number of Participants | Hydration in Individual Tests |
|---|
| Test 1 | Test 2 | Test 3 |
|---|
| I-W-NH | 2 | I | W | NH |
| W-NH-I | 2 | W | NH | I |
| NH-I-W | 2 | NH | I | W |
| I-NH-W | 2 | I | NH | W |
| W-I-NH | 2 | W | I | NH |
| NH-W-I | 2 | NH | W | I |
Table 2.
Mean values of the change in body mass (∆BM) [kg] between pre-test (preT) and post-test (postT) results in three consecutive tests (1, 2, 3) performed using three different hydration strategies (I, W, NH): marginal means (LSM); standard errors (SE); and lower (LCI) and upper (UCI) confidence intervals.
Table 2.
Mean values of the change in body mass (∆BM) [kg] between pre-test (preT) and post-test (postT) results in three consecutive tests (1, 2, 3) performed using three different hydration strategies (I, W, NH): marginal means (LSM); standard errors (SE); and lower (LCI) and upper (UCI) confidence intervals.
| | Statistics | Hydration Strategy | Test Sequence |
|---|
| I | W | NH | 1 | 2 | 3 |
|---|
| ∆BM | LSM | −0.73 | −0.94 | −0.72 | −1.02 | −1.01 | −0.35 |
| SE | 0.19 | 0.19 | 0.19 | 0.19 | 0.19 | 0.19 |
| LCI | −1.11 | −1.32 | −1.10 | −1.41 | −1.39 | −0.74 |
| UCI | −0.35 | −0.56 | −0.33 | −0.64 | −0.63 | 0.03 |
Table 3.
Results of analysis of variance for the change in body mass (∆BM) between the pre-test (preT) and post-test (postT) results of the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 3.
Results of analysis of variance for the change in body mass (∆BM) between the pre-test (preT) and post-test (postT) results of the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| ∆BM | DFm | F | P | DFm | F | P | DFm | F | P |
| 17.7 | 0.39 | 0.683 | 17.7 | 3.59 | 0.049 | 17.2 | 1.46 | 0.257 |
Table 4.
Results of analysis of variance for the change in plasma volume ∆PV calculated from HB and HCT results measured before and 0, 1, and 24 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 4.
Results of analysis of variance for the change in plasma volume ∆PV calculated from HB and HCT results measured before and 0, 1, and 24 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| Time | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| | DFm | F | P | DFm | F | P | DFm | F | P |
| 0 | 9.4 | 11.65 | 0.003 | 8.0 | 2.14 | 0.181 | 11.3 | 3.80 | 0.035 |
| 1 | 15.5 | 9.54 | 0.002 | 15.5 | 0.03 | 0.968 | 27.0 | 1.07 | 0.390 |
| 24 | 15.3 | 2.41 | 0.123 | 15.3 | 3.01 | 0.079 | 23.7 | 0.52 | 0.723 |
Table 5.
Results for plasma volume change ∆PV [%] estimated using Dill and Costill’s formula (1974) as modified by Harisson et al. (1982). Calculations were made based on HB and HCT results measured before the test and 0, 1, and 24 h after the test, performed with different hydration strategies in three consecutive tests: marginal means (LSM), standard errors (SE), and upper (UCI) and lower (LCI) confidence intervals.
Table 5.
Results for plasma volume change ∆PV [%] estimated using Dill and Costill’s formula (1974) as modified by Harisson et al. (1982). Calculations were made based on HB and HCT results measured before the test and 0, 1, and 24 h after the test, performed with different hydration strategies in three consecutive tests: marginal means (LSM), standard errors (SE), and upper (UCI) and lower (LCI) confidence intervals.
| Time | Statistics | Hydration Strategy | Test Sequence |
|---|
| | I | W | NH | 1 | 2 | 3 |
|---|
| 0 | LSM | 0.5 | −2.4 | −4.4 | −0.9 | −2.2 | −3.1 |
| SE | 1.2 | 1.2 | 1.2 | 1.6 | 0.6 | 1.5 |
| LCI | −2.1 | −4.9 | −6.9 | −4.5 | −3.7 | −6.6 |
| UCI | 3.0 | 0.2 | −1.8 | 2.7 | −0.7 | 0.3 |
| 1 | LSM | 5.2 | 2.2 | −1.6 | 2.0 | 2.1 | 1.7 |
| SE | 1.2 | 1.2 | 1.2 | 1.2 | 1.2 | 1.2 |
| LCI | 2.6 | −0.3 | −4.1 | −0.5 | −0.4 | −0.8 |
| UCI | 7.7 | 4.7 | 0.9 | 4.5 | 4.6 | 4.2 |
| 24 | LSM | 4.0 | 1.0 | 6.2 | 7.1 | 2.6 | 1.5 |
| SE | 1.7 | 1.7 | 1.7 | 1.7 | 1.7 | 1.7 |
| LCI | 0.5 | −2.6 | 2.7 | 3.6 | −0.9 | −2.0 |
| UCI | 7.6 | 4.5 | 9.8 | 10.6 | 6.1 | 5.1 |
Table 6.
Results of analysis of variance for pro-oxidant/antioxidant status and oxidative status measured before the test (preT) and 0, 1, and 24 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 6.
Results of analysis of variance for pro-oxidant/antioxidant status and oxidative status measured before the test (preT) and 0, 1, and 24 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| t | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| | DFm | F | P | DFm | F | P | DFm | F | P |
| preT | 16.7 | 0.33 | 0.723 | 16.7 | 0.39 | 0.683 | 23.8 | 1.45 | 0.249 |
| 0 | 17.2 | 0.06 | 0.939 | 17.2 | 0.01 | 0.991 | 26.7 | 2.60 | 0.059 |
| 1 | 12.7 | 1.04 | 0.381 | 8.0 | 0.43 | 0.663 | 14.1 | 3.62 | 0.031 |
| 24 | 17.2 | 0.40 | 0.679 | 17.2 | 0.58 | 0.570 | 25.6 | 1.01 | 0.423 |
Table 7.
Results of pro-oxidative and antioxidant status [µmol-L−1] measured before the test (preT) and 0, 1, and 24 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
Table 7.
Results of pro-oxidative and antioxidant status [µmol-L−1] measured before the test (preT) and 0, 1, and 24 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
| Collection Time | Statistic | Hydration Strategy | Test Sequence |
|---|
| I | W | NH | 1 | 2 | 3 |
|---|
| preT | LSM (log) | 2.49 | 2.48 | 2.49 | 2.49 | 2.49 | 2.48 |
| SE (log) | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| LSM | 306.69 | 305.21 | 312.32 | 311.39 | 309.24 | 303.60 |
| LCI | 288.40 | 287.01 | 293.70 | 292.82 | 290.80 | 285.50 |
| UCI | 326.14 | 324.56 | 332.05 | 331.13 | 328.85 | 322.85 |
| 0 | LSM (log) | 2.49 | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| SE (log) | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| LSM | 311.82 | 315.21 | 313.62 | 314.12 | 313.62 | 312.82 |
| LCI | 295.80 | 299.02 | 297.51 | 297.99 | 297.51 | 296.76 |
| UCI | 328.70 | 332.28 | 330.60 | 331.21 | 330.67 | 329.84 |
| 1 | LSM (log) | 2.50 | 2.50 | 2.51 | 2.51 | 2.49 | 2.50 |
| SE (log) | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| LSM | 313.33 | 312.68 | 324.79 | 322.03 | 311.46 | 317.25 |
| LCI | 299.43 | 298.74 | 310.38 | 310.10 | 288.00 | 306.48 |
| UCI | 327.94 | 327.19 | 339.94 | 334.35 | 336.82 | 328.47 |
| 24 | LSM (log) | 2.51 | 2.50 | 2.51 | 2.51 | 2.51 | 2.50 |
| SE (log) | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| LSM | 321.96 | 319.30 | 327.27 | 324.41 | 326.81 | 317.39 |
| LCI | 305.28 | 302.83 | 310.38 | 307.61 | 309.88 | 300.95 |
| UCI | 339.47 | 336.67 | 345.06 | 342.06 | 344.59 | 334.66 |
Table 8.
Results of analysis of variance for myoglobin measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 8.
Results of analysis of variance for myoglobin measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| t | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| | DFm | F | P | DFm | F | P | DFm | F | P |
| preT | 16.5 | 0.28 | 0.760 | 16.5 | 1.44 | 0.266 | 24.8 | 0.77 | 0.557 |
| 1 | 8.7 | 5.30 | 0.031 | 8.8 | 12.69 | 0.003 | 11.5 | 1.00 | 0.449 |
| 24 | 15.7 | 0.39 | 0.685 | 15.7 | 9.61 | 0.002 | 22.5 | 0.54 | 0.705 |
| 48 | 12.8 | 1.87 | 0.195 | 6.8 | 1.70 | 0.251 | 8.5 | 1.94 | 0.192 |
Table 9.
Results for myoglobin [ng·mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
Table 9.
Results for myoglobin [ng·mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
| Collection Time | Statistic | Hydration Strategy | Test Sequence |
|---|
| I | W | NH | 1 | 2 | 3 |
|---|
| preT | LSM (log) | 1.08 | 1.06 | 1.02 | 1.12 | 1.04 | 0.99 |
| SE (log) | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 | 0.06 |
| LSM | 11.90 | 11.45 | 10.35 | 13.26 | 10.96 | 9.70 |
| LCI | 8.97 | 8.64 | 7.69 | 10.00 | 8.14 | 7.31 |
| UCI | 15.78 | 15.18 | 13.93 | 17.59 | 14.76 | 12.86 |
| 1 | LSM (log) | 1.44 | 1.60 | 1.61 | 1.80 | 1.50 | 1.36 |
| SE (log) | 0.07 | 0.07 | 0.07 | 0.11 | 0.07 | 0.04 |
| LSM | 27.67 | 40.00 | 41.10 | 63.56 | 31.34 | 22.84 |
| LCI | 20.06 | 29.01 | 29.53 | 36.08 | 21.33 | 18.43 |
| UCI | 38.16 | 55.17 | 57.21 | 111.97 | 46.05 | 28.31 |
| 24 | LSM (log) | 1.27 | 1.21 | 1.29 | 1.45 | 1.28 | 1.04 |
| SE (log) | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| LSM | 18.76 | 16.10 | 19.30 | 28.24 | 18.89 | 10.92 |
| LCI | 11.83 | 10.15 | 11.99 | 17.81 | 11.74 | 6.89 |
| UCI | 29.75 | 25.53 | 31.07 | 44.78 | 30.42 | 17.33 |
| 48 | LSM (log) | 1.18 | 1.21 | 1.08 | 1.20 | 1.24 | 1.03 |
| SE (log) | 0.07 | 0.07 | 0.07 | 0.08 | 0.09 | 0.07 |
| LSM | 15.09 | 16.06 | 12.15 | 15.85 | 17.34 | 10.71 |
| LCI | 10.51 | 11.19 | 8.43 | 10.44 | 10.74 | 7.42 |
| UCI | 21.65 | 23.05 | 17.49 | 24.03 | 28.00 | 15.47 |
Table 10.
Results of analysis of variance for lactate dehydrogenase measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 10.
Results of analysis of variance for lactate dehydrogenase measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| t | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| | DFm | F | P | DFm | F | P | DFm | F | P |
| preT | 16.0 | 12.73 | 0.001 | 16.0 | 0.33 | 0.723 | 16.1 | 1.45 | 0.263 |
| 1 | 16.0 | 5.20 | 0.018 | 16.0 | 1.12 | 0.352 | 16.2 | 0.80 | 0.541 |
| 24 | 16.0 | 8.51 | 0.003 | 16.0 | 0.91 | 0.421 | 16.1 | 1.58 | 0.229 |
| 48 | 16.0 | 2.69 | 0.098 | 16.0 | 6.43 | 0.009 | 16.2 | 1.42 | 0.271 |
Table 11.
Results for lactate dehydrogenase [U·mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
Table 11.
Results for lactate dehydrogenase [U·mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
| Collection Time | Statistic | Hydration Strategy | Test Sequence |
|---|
| I | W | NH | 1 | 2 | 3 |
|---|
| preT | LSM (log) | 2.42 | 2.44 | 2.35 | 2.39 | 2.40 | 2.41 |
| SE (log) | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 |
| LSM | 265.34 | 274.22 | 221.72 | 248.26 | 252.23 | 257.57 |
| LCI | 139.70 | 144.38 | 116.73 | 130.74 | 132.80 | 135.61 |
| UCI | 503.85 | 520.83 | 421.02 | 471.52 | 479.07 | 489.22 |
| 1 | LSM (log) | 2.43 | 2.41 | 2.35 | 2.40 | 2.38 | 2.42 |
| SE (log) | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 |
| LSM | 271.46 | 258.82 | 224.54 | 248.71 | 240.71 | 263.45 |
| LCI | 138.39 | 131.98 | 114.50 | 126.82 | 122.74 | 134.34 |
| UCI | 532.35 | 507.57 | 440.45 | 487.75 | 472.17 | 516.65 |
| 24 | LSM (log) | 2.43 | 2.43 | 2.36 | 2.41 | 2.42 | 2.40 |
| SE (log) | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 | 0.13 |
| LSM | 271.21 | 269.15 | 230.41 | 254.57 | 264.79 | 249.52 |
| LCI | 142.82 | 141.71 | 121.34 | 134.06 | 139.41 | 131.40 |
| UCI | 515.11 | 511.09 | 437.52 | 483.39 | 502.81 | 473.91 |
| 48 | LSM (log) | 2.41 | 2.37 | 2.36 | 2.34 | 2.42 | 2.38 |
| SE (log) | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 |
| LSM | 257.34 | 235.83 | 229.51 | 220.85 | 265.40 | 237.63 |
| LCI | 129.90 | 119.04 | 115.85 | 111.48 | 133.97 | 119.95 |
| UCI | 509.80 | 467.09 | 454.57 | 437.52 | 525.65 | 470.65 |
Table 12.
Results of analysis of variance for WBC measured before the test (preT) and 0, 1, and 24 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 12.
Results of analysis of variance for WBC measured before the test (preT) and 0, 1, and 24 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| t | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| | DFm | F | P | DFm | F | P | DFm | F | P |
| preT | 16.9 | 2.00 | 0.166 | 16.9 | 1.43 | 0.267 | 23.7 | 2.36 | 0.083 |
| 0 | 15.7 | 11.81 | 0.001 | 15.7 | 5.80 | 0.013 | 21.3 | 0.83 | 0.520 |
| 1 | 16.0 | 9.51 | 0.002 | 16.0 | 2.39 | 0.123 | 24.8 | 0.47 | 0.756 |
| 24 | 12.6 | 6.13 | 0.014 | 8.3 | 15.64 | 0.002 | 12.2 | 7.69 | 0.003 |
Table 13.
Results for WBC [103/mm3] measured before the test (preT) and 0, 1, and 24 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
Table 13.
Results for WBC [103/mm3] measured before the test (preT) and 0, 1, and 24 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
| Collection Time | Statistic | Hydration Strategy | Test Sequence |
|---|
| I | W | NH | 1 | 2 | 3 |
|---|
| preT | LSM (log) | 0.77 | 0.74 | 0.73 | 0.74 | 0.73 | 0.76 |
| SE (log) | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| LSM | 5.83 | 5.45 | 5.39 | 5.45 | 5.42 | 5.79 |
| LCI | 5.33 | 4.97 | 4.92 | 4.98 | 4.95 | 5.29 |
| UCI | 6.39 | 5.97 | 5.90 | 5.97 | 5.94 | 6.34 |
| 0 | LSM (log) | 0.96 | 1.13 | 1.06 | 1.12 | 1.00 | 1.03 |
| SE (log) | 0.04 | 0.04 | 0.04 | 0.04 | 0.04 | 0.04 |
| LSM | 9.03 | 13.62 | 11.42 | 13.13 | 9.90 | 10.80 |
| LCI | 7.37 | 11.11 | 9.32 | 10.71 | 8.08 | 8.82 |
| UCI | 11.07 | 16.68 | 13.98 | 16.08 | 12.13 | 13.24 |
| 1 | LSM (log) | 0.96 | 1.14 | 1.04 | 1.10 | 1.01 | 1.03 |
| SE (log) | 0.04 | 0.04 | 0.04 | 0.04 | 0.04 | 0.04 |
| LSM | 9.07 | 13.76 | 11.00 | 12.47 | 10.17 | 10.81 |
| LCI | 7.54 | 11.44 | 9.14 | 10.37 | 8.45 | 8.99 |
| UCI | 10.91 | 16.55 | 13.23 | 15.01 | 12.24 | 13.01 |
| 24 | LSM (log) | 0.81 | 0.74 | 0.77 | 0.80 | 0.72 | 0.80 |
| SE (log) | 0.02 | 0.02 | 0.02 | 0.03 | 0.02 | 0.02 |
| LSM | 6.48 | 5.50 | 5.84 | 6.29 | 5.28 | 6.27 |
| LCI | 5.86 | 4.98 | 5.28 | 5.35 | 4.81 | 5.80 |
| UCI | 7.16 | 6.08 | 6.46 | 7.39 | 5.79 | 6.79 |
Table 14.
Results for WBC [103/mm3] measured 24 h after the test performed using different hydration strategies (I, W, BN) in three consecutive tests (1, 2, 3): marginal means (LSM) and standard errors (SE) for values after logarithmic transformation (log).
Table 14.
Results for WBC [103/mm3] measured 24 h after the test performed using different hydration strategies (I, W, BN) in three consecutive tests (1, 2, 3): marginal means (LSM) and standard errors (SE) for values after logarithmic transformation (log).
| WBC | Test Sequence | 1 | 2 | 3 |
|---|
| Strategy | I | W | NH | I | W | NH | I | W | NH |
|---|
| 24 | LSM | 0.905 | 0.789 | 0.702 | 0.706 | 0.640 | 0.821 | 0.824 | 0.793 | 0.776 |
| SE | 0.04 | 0.04 | 0.04 | 0.02 | 0.02 | 0.02 | 0.03 | 0.03 | 0.03 |
Table 15.
Results of analysis of variance for interleukin 1β measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 15.
Results of analysis of variance for interleukin 1β measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| t | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| | DFm | F | P | DFm | F | P | DFm | F | P |
| preT | 11.6 | 2.78 | 0.103 | 6.6 | 6.46 | 0.028 | 9.3 | 1.34 | 0.327 |
| 1 | 15.1 | 0.56 | 0.580 | 15.1 | 0.65 | 0.535 | 23.3 | 1.02 | 0.416 |
| 24 | 11.8 | 1.98 | 0.182 | 5.9 | 2.53 | 0.162 | 8.1 | 3.09 | 0.081 |
| 48 | 14.9 | 5.08 | 0.021 | 14.9 | 0.55 | 0.590 | 22.9 | 0.22 | 0.923 |
Table 16.
Results for interleukin 1β [pg∙mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
Table 16.
Results for interleukin 1β [pg∙mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
| Collection Time | Statistic | Hydration Strategy | Test Sequence |
|---|
| I | W | NH | 1 | 2 | 3 |
|---|
| preT | LSM (log) | 0.24 | 0.54 | 0.52 | 0.37 | 0.30 | 0.63 |
| SE (log) | 0.12 | 0.11 | 0.11 | 0.09 | 0.20 | 0.09 |
| LSM | 1.73 | 3.49 | 3.34 | 2.37 | 1.99 | 4.26 |
| LCI | 0.96 | 2.03 | 1.96 | 1.47 | 0.66 | 2.68 |
| UCI | 3.10 | 6.00 | 5.68 | 3.82 | 5.99 | 6.79 |
| 1 | LSM (log) | 0.76 | 0.75 | 0.92 | 0.73 | 0.92 | 0.78 |
| SE (log) | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 |
| LSM | 5.71 | 5.62 | 8.30 | 5.32 | 8.39 | 5.97 |
| LCI | 2.98 | 2.93 | 4.33 | 2.78 | 4.38 | 3.11 |
| UCI | 10.93 | 10.77 | 15.91 | 10.19 | 16.08 | 11.43 |
| 24 | LSM (log) | 0.79 | 0.93 | 1.02 | 0.98 | 0.90 | 0.86 |
| SE (log) | 0.11 | 0.10 | 0.10 | 0.08 | 0.15 | 0.09 |
| LSM | 6.16 | 8.41 | 10.48 | 9.58 | 7.86 | 7.22 |
| LCI | 3.59 | 5.09 | 6.25 | 6.10 | 3.28 | 4.43 |
| UCI | 10.59 | 13.90 | 17.59 | 15.03 | 18.81 | 11.80 |
| 48 | LSM (log) | 0.37 | 0.81 | 0.65 | 0.65 | 0.52 | 0.66 |
| SE (log) | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 |
| LSM | 2.33 | 6.44 | 4.46 | 4.43 | 3.34 | 4.52 |
| LCI | 1.29 | 3.56 | 2.47 | 2.45 | 1.85 | 2.50 |
| UCI | 4.21 | 11.65 | 8.07 | 8.01 | 6.04 | 8.18 |
Table 17.
Results of analysis of variance for CRP measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
Table 17.
Results of analysis of variance for CRP measured before the test (preT) and 1, 24, and 48 h after the exercise test: significance of (P) and F-statistic along with the number of degrees of freedom (DFi—in the numerator; DFm—in the denominator) for the fixed effects (hydration strategy, test sequence, and their interaction).
| t | Hydration Strategy | Test Sequence | Interaction |
|---|
| | (DFi = 2) | | | (DFi = 2) | | | (DFi = 4) | |
|---|
| | DFm | F | P | DFm | F | P | DFm | F | P |
| preT | 10.4 | 1.13 | 0.361 | 7.1 | 4.43 | 0.057 | 6.6 | 10.85 | 0.005 |
| 1 | 10.6 | 1.53 | 0.262 | 7.3 | 5.32 | 0.038 | 9.3 | 8.11 | 0.004 |
| 24 | 12.5 | 0.15 | 0.863 | 12.5 | 2.57 | 0.116 | 22.2 | 2.93 | 0.044 |
| 48 | 13.3 | 0.65 | 0.536 | 13.3 | 1.07 | 0.371 | 23.8 | 3.42 | 0.024 |
Table 18.
Results for CRP [mg·mL−1] measured before (preT) and one hour (1) after the test performed using different hydration strategies (I, W, BN) in three consecutive tests (1, 2, 3): marginal means (LSM) and standard errors (SE) for values after logarithmic transformation (log).
Table 18.
Results for CRP [mg·mL−1] measured before (preT) and one hour (1) after the test performed using different hydration strategies (I, W, BN) in three consecutive tests (1, 2, 3): marginal means (LSM) and standard errors (SE) for values after logarithmic transformation (log).
| CRP | Test Sequence | 1 | 2 | 3 |
|---|
| Strategy | I | W | NH | I | W | NH | I | W | NH |
|---|
| preT | LSM (log) | −0.097 | −0.204 | −0.050 | −0.298 | −0.491 | 0.065 | 0.094 | −0.058 | −0.449 |
| SE (log) | 0.22 | 0.22 | 0.22 | 0.14 | 0.14 | 0.14 | 0.15 | 0.15 | 0.15 |
| 1 | LSM (log) | −0.132 | −0.214 | −0.023 | −0.288 | −0.534 | 0.039 | 0.060 | −0.095 | −0.398 |
| SE (log) | 0.22 | 0.22 | 0.22 | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 | 0.15 |
Table 19.
Results for CRP [mg∙mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
Table 19.
Results for CRP [mg∙mL−1] measured before the test (preT) and 1, 24, and 48 h after the test performed for different hydration strategies in three consecutive tests: marginal means (LSM); standard errors (SE) for values after logarithmic transformation (log); and marginal means and upper (UCI) and lower (LCI) confidence intervals restored to original form.
| Collection Time | Statistic | Hydration Strategy | Test Sequence |
|---|
| I | W | NH | 1 | 2 | 3 |
|---|
| preT | LSM (log) | −0.10 | −0.25 | −0.14 | −0.12 | −0.24 | −0.14 |
| SE (log) | 0.13 | 0.13 | 0.13 | 0.14 | 0.13 | 0.14 |
| LSM | 0.79 | 0.56 | 0.72 | 0.76 | 0.57 | 0.73 |
| LCI | 0.41 | 0.29 | 0.37 | 0.37 | 0.29 | 0.37 |
| UCI | 1.54 | 1.09 | 1.39 | 1.60 | 1.13 | 1.45 |
| 1 | LSM (log) | −0.12 | −0.28 | −0.13 | −0.12 | −0.26 | −0.14 |
| SE (log) | 0.13 | 0.13 | 0.13 | 0.14 | 0.13 | 0.13 |
| LSM | 0.76 | 0.52 | 0.75 | 0.75 | 0.55 | 0.72 |
| LCI | 0.40 | 0.27 | 0.39 | 0.36 | 0.28 | 0.37 |
| UCI | 1.45 | 1.00 | 1.43 | 1.57 | 1.06 | 1.39 |
| 24 | LSM (log) | 0.02 | 0.00 | 0.06 | 0.17 | −0.07 | −0.02 |
| SE (log) | 0.11 | 0.11 | 0.11 | 0.11 | 0.11 | 0.11 |
| LSM | 1.04 | 1.00 | 1.16 | 1.49 | 0.86 | 0.95 |
| LCI | 0.62 | 0.60 | 0.68 | 0.89 | 0.51 | 0.55 |
| UCI | 1.75 | 1.69 | 1.98 | 2.51 | 1.44 | 1.62 |
| 48 | LSM (log) | −0.04 | −0.14 | −0.14 | −0.02 | −0.14 | −0.16 |
| SE (log) | 0.09 | 0.09 | 0.10 | 0.09 | 0.09 | 0.10 |
| LSM | 0.92 | 0.73 | 0.72 | 0.96 | 0.72 | 0.70 |
| LCI | 0.59 | 0.47 | 0.45 | 0.61 | 0.46 | 0.44 |
| UCI | 1.43 | 1.14 | 1.14 | 1.50 | 1.12 | 1.11 |



 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}