

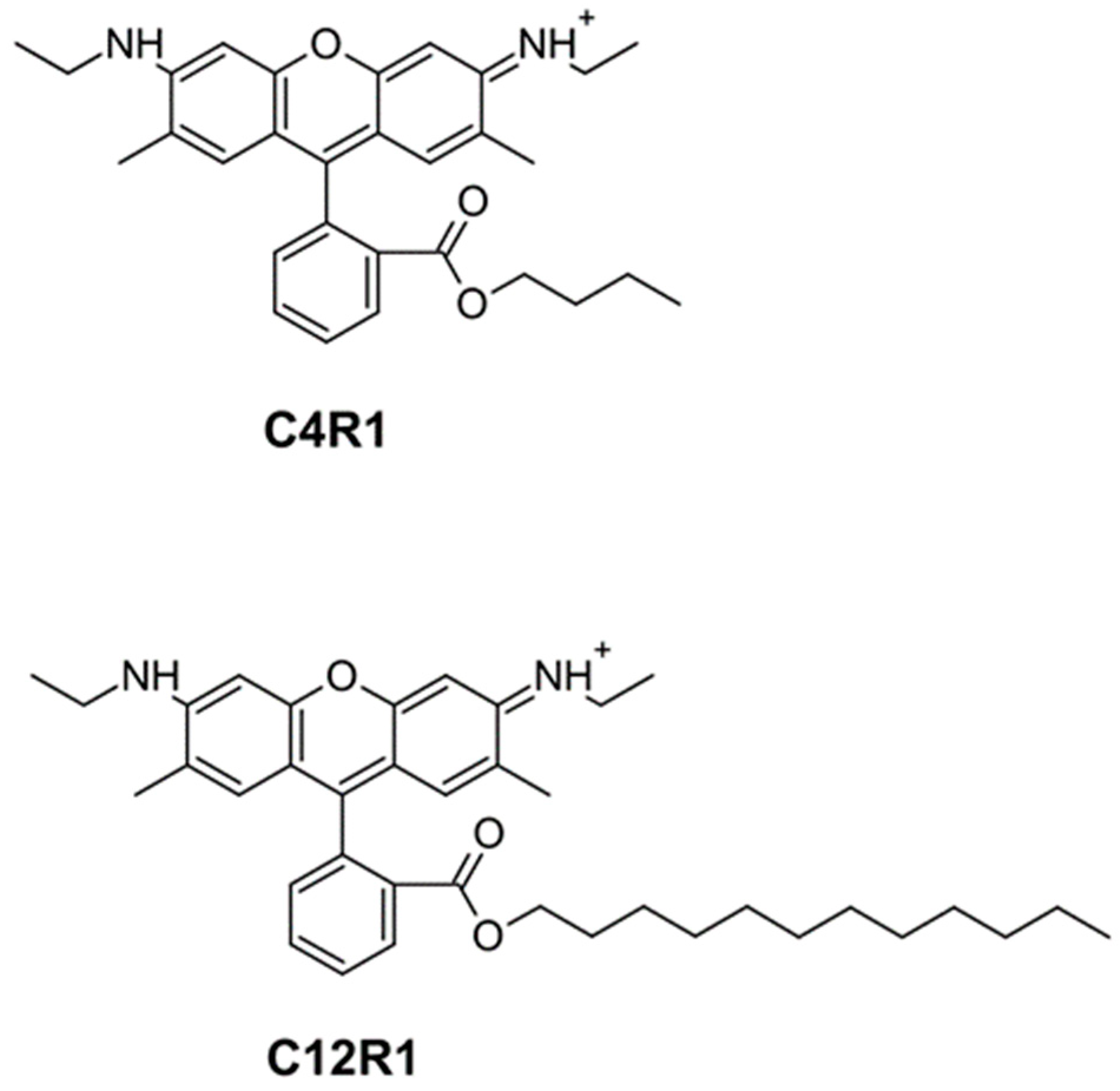
Mitochondrial ATP Synthase and Mild Uncoupling by Butyl Ester of Rhodamine 19, C4R1

 , ,
, ,  , ,
, ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthesis of Rhodamine Derivatives
2.2. Isolation of Rat Liver Mitochondria
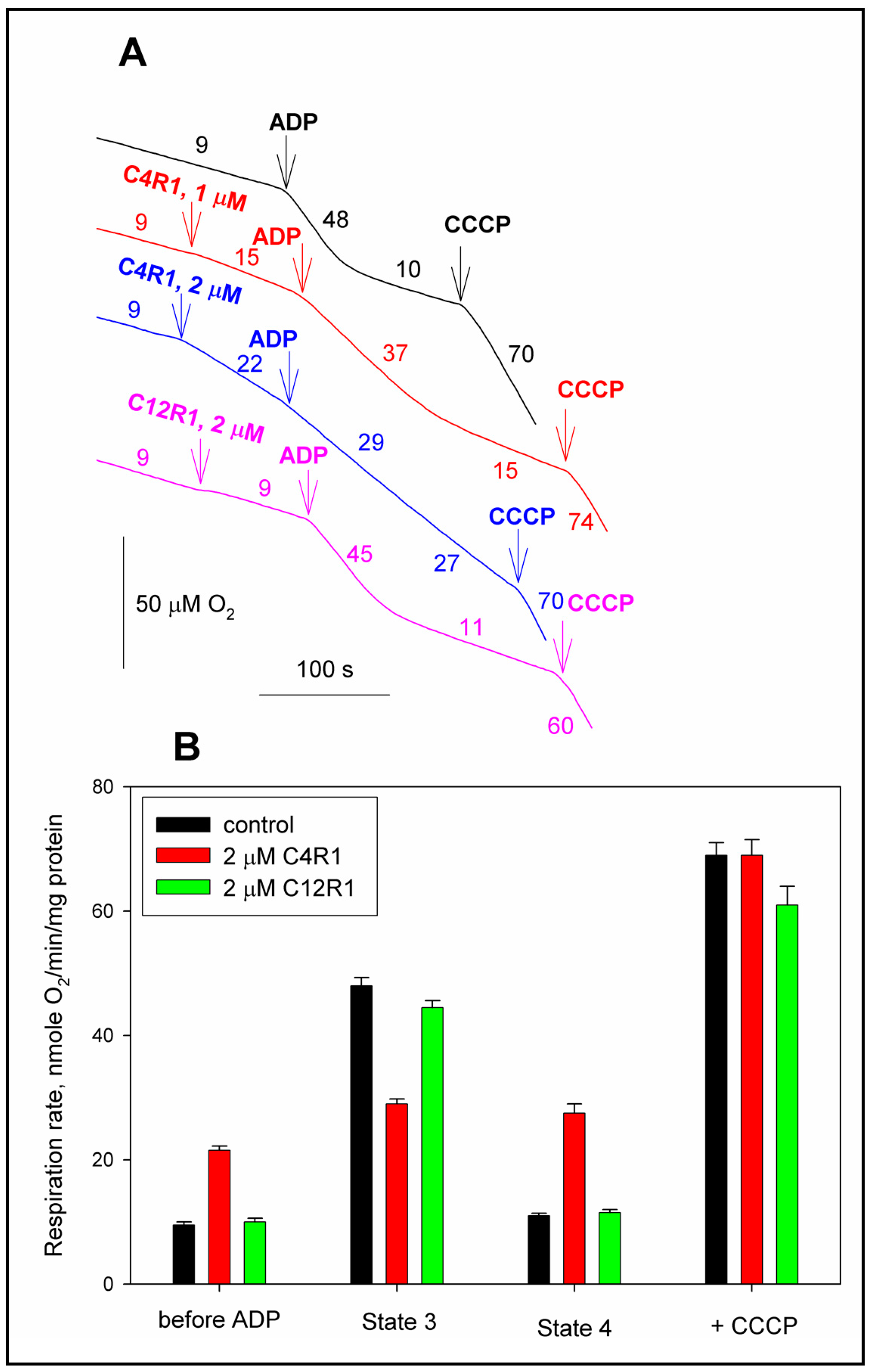
2.3. Mitochondrial Respiration
2.4. Isolation of ATP Synthase Complex (FoF1, Complex V)
2.5. Electric Potential of Membranes of Bacillus subtilis with DiS-C3-(5)
2.6. Binding C4R and C12R1 with Mitochondria and Bacillus subtilis Elucidated by 1D Electrophoresis
2.7. Two-Dimensional Gel Electrophoresis
2.8. Mass Spectrometry
2.9. Molecular Docking
3. Results
3.1. Uncoupling Activity of C4R1 in Mitochondria
3.2. The Effects of C4R1 and C12R1 in Bacillus subtilis Intact Cells
3.3. Binding C4R1 and C12R1 with Mitochondria and Bacterial Cells
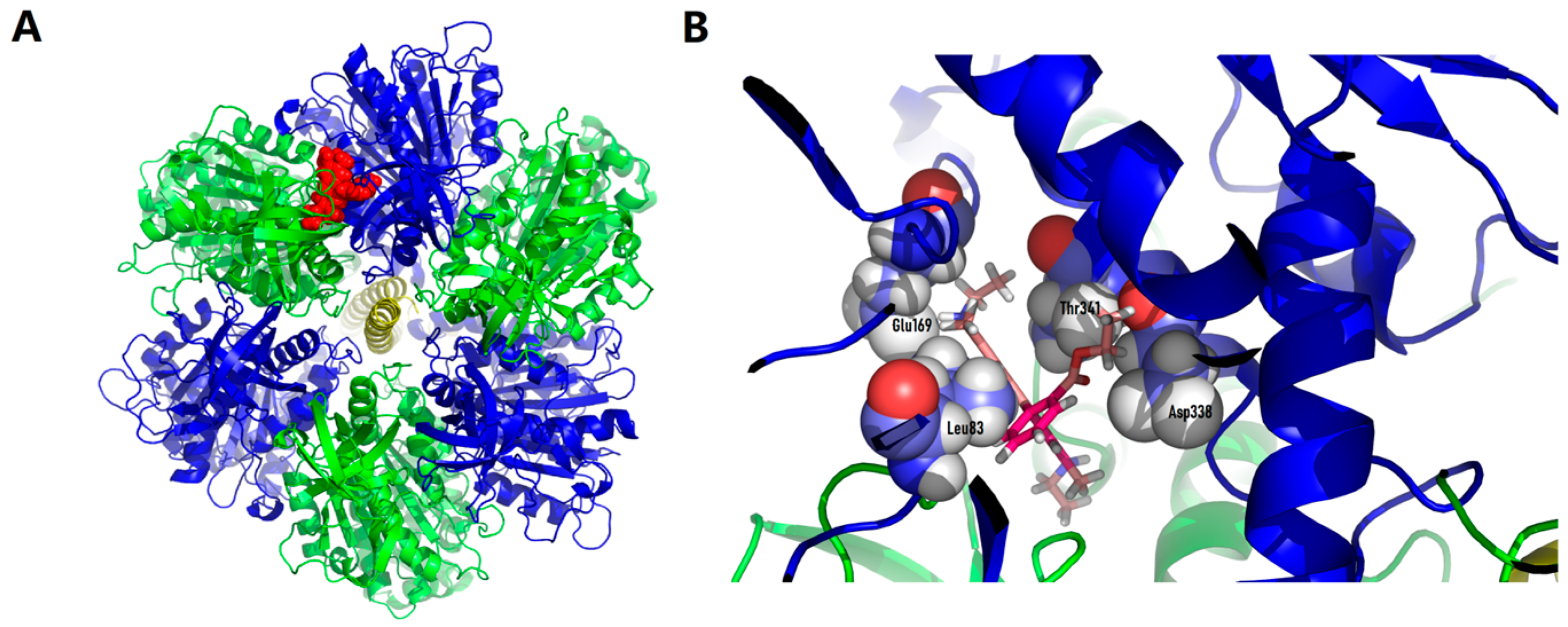
3.4. Identification of the Mitochondrial Component That Binds C4R1
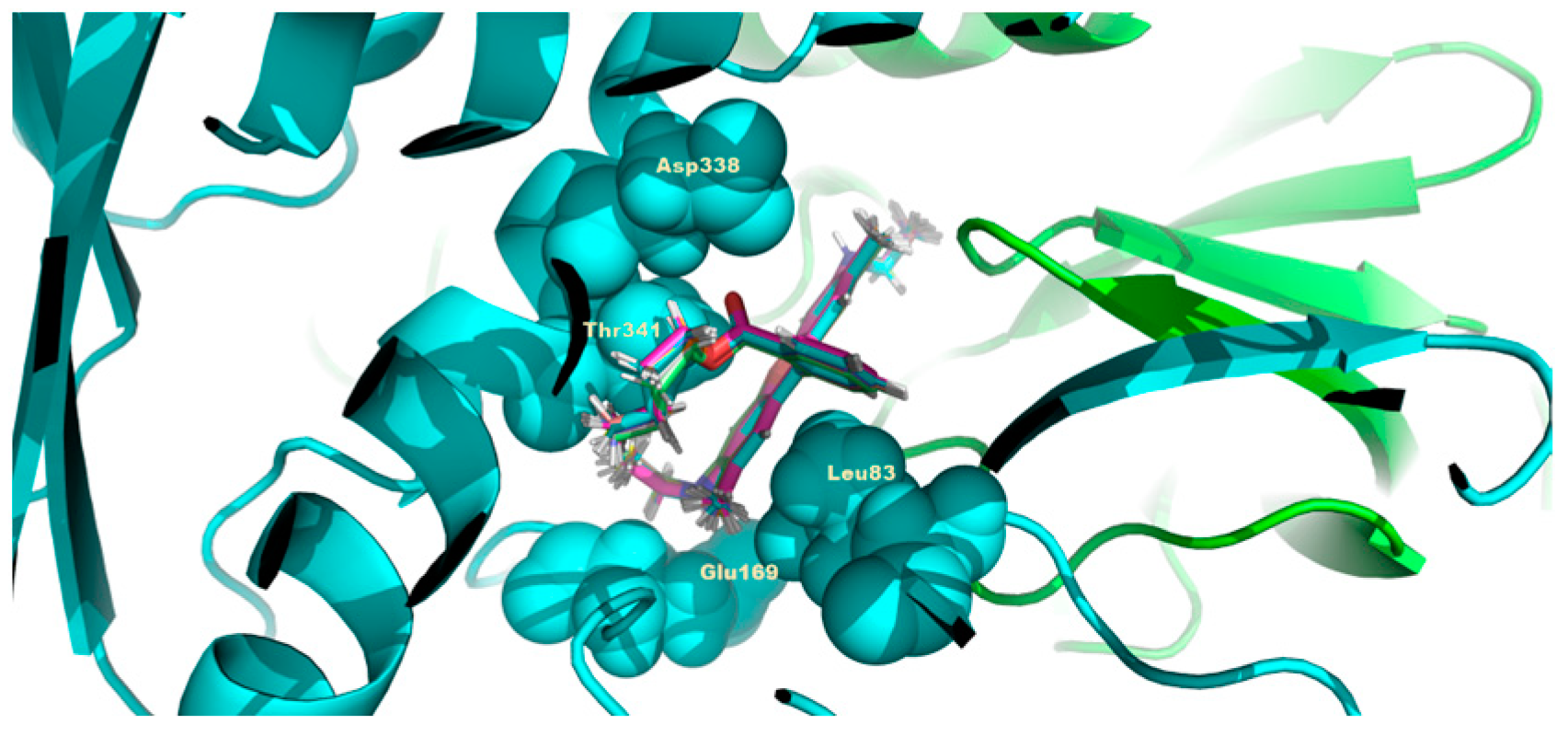
3.5. Modelling of C4R1 Binding to Mitochondrial Component
4. Discussion
5. Conclusions. Uncoupling Proteins: One More?
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev. 1966, 41, 445–501. [Google Scholar] [CrossRef]
- Walker, J.E. The ATP Synthase: The understood, the uncertain and the unknown. Biochem. Soc. Trans. 2013, 41, 1–16. [Google Scholar] [CrossRef]
- Skulachev, V.P. Uncoupling: New approaches to an old problem of bioenergetics. Biochim. Biophys. Acta (BBA)—Bioenerg. 1998, 1363, 100–124. [Google Scholar] [CrossRef]
- Hayashi, M.; Nakayama, Y.; Unemoto, T. Existence of Na+ -translocating NADH-quinone reductase in Haemophilus influenzae. FEBS Lett. 1996, 381, 174–176. [Google Scholar] [CrossRef]
- Häse, C.C.; Barquera, B. Role of sodium bioenergetics in Vibrio Cholerae. Biochim. Biophys. Acta (BBA)—Bioenerg. 2001, 1505, 169–178. [Google Scholar] [CrossRef]
- Bogachev, A.V.; Verkhovsky, M.I. Na(+)-Translocating NADH: Quinone oxidoreductase: Progress achieved and prospects of investigations. Biochemistry (Moscow) 2005, 70, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Bielawski, J.; Thompson, T.E.; Lehninger, A.L. The effect of 2,4-dinitrophenol on the electrical resistance of phospholipid bilayer membranes. Biochem. Biophys. Res. Commun. 1966, 24, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Liberman, E.A.; Topaly, V.P.; Tsofina, L.M.; Jasaitis, A.A.; Skulachev, V.P. Mechanism of coupling of oxidative phosphorylation and the membrane potential of mitochondria. Nature 1969, 222, 1076–1078. [Google Scholar] [CrossRef]
- Klingenberg, M.; Hackenberg, H.; Krämer, R.; Lin, C.S.; Aquila, H. Two transport proteins from mitochondria: I. Mechanistic aspects of asymmetry of the ADP, ATP translocator II. The uncoupling protein of brown adipose tissue mitochondria. Ann. N. Y. Acad. Sci. 1980, 358, 83–95. [Google Scholar] [CrossRef]
- Nedergaard, J.; Matthias, A.; Golozoubova, V.; Jacobsson, A.; Cannon, B. B UCP1: The original uncoupling protein--and perhaps the only one? New perspectives on UCP1, UCP2, and UCP3 in the light of the bioenergetics of the UCP1-ablated mice. J. Bioenerg. Biomembr. 1999, 31, 475–491. [Google Scholar] [CrossRef] [PubMed]
- Andreyev, A.Y.; Bondareva, T.O.; Dedukhova, V.I.; Mokhova, E.N.; Skulachev, V.P.; Tsofina, L.M.; Volkov, N.I.; Vygodina, T.V. The ATP/ADP-antiporter is involved in the uncoupling effect of fatty acids on mitochondria. Eur. J. Biochem. 1989, 182, 585–592. [Google Scholar] [CrossRef]
- Samartsev, V.N.; Smirnov, A.V.; Zeldi, I.P.; Markova, O.V.; Mokhova, E.N.; Skulachev, V.P. Involvement of aspartate/glutamate antiporter in fatty acid-induced uncoupling of liver mitochondria. Biochim. Biophys. Acta (BBA)—Bioenerg. 1997, 1319, 251–257. [Google Scholar] [CrossRef]
- Bertholet, A.M.; Chouchani, E.T.; Kazak, L.; Angelin, A.; Fedorenko, A.; Long, J.Z.; Vidoni, S.; Garrity, R.; Cho, J.; Terada, N.; et al. H+ Transport Is an Integral Function of the Mitochondrial ADP/ATP Carrier. Nature 2019, 571, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Bertholet, A.M.; Natale, A.M.; Bisignano, P.; Suzuki, J.; Fedorenko, A.; Hamilton, J.; Brustovetsky, T.; Kazak, L.; Garrity, R.; Chouchani, E.T.; et al. Mitochondrial Uncouplers Induce Proton Leak by Activating AAC and UCP1. Nature 2022, 606, 180–187. [Google Scholar] [CrossRef]
- Žuna, K.; Jovanović, O.; Khailova, L.S.; Škulj, S.; Brkljača, Z.; Kreiter, J.; Kotova, E.A.; Vazdar, M.; Antonenko, Y.N.; Pohl, E.E. Mitochondrial Uncoupling Proteins (UCP1-UCP3) and Adenine Nucleotide Translocase (ANT1) Enhance the Protonophoric Action of 2,4-Dinitrophenol in Mitochondria and Planar Bilayer Membranes. Biomolecules 2021, 11, 1178. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Frøkiær, J.; Marples, D.; Kwon, T.H.; Agre, P.; Knepper, M.A. Aquaporins in the kidney: From molecules to medicine. Physiol. Rev. 2002, 82, 205–244. [Google Scholar] [CrossRef]
- Starkov, A.A.; Dedukhova, V.I.; Skulachev, V.P. 6-Ketocholestanol Abolishes the Effect of the Most Potent Uncouplers of Oxidative Phosphorylation in Mitochondria. FEBS Lett. 1994, 355, 305–308. [Google Scholar] [CrossRef]
- Grivennikova, V.G.; Khailova, L.S.; Zharova, T.V.; Kotova, E.A.; Antonenko, Y.N. Inhibition of Respiratory Complex I by 6-Ketocholestanol: Relevance to Recoupling Action in Mitochondria. Biochim. Biophys. Acta (BBA)—Bioenerg. 2022, 1863, 148594. [Google Scholar] [CrossRef]
- Brand, M.D.; Esteves, T.C. Physiological functions of the mitochondrial uncoupling proteins UCP2 and UCP3. Cell Metab. 2005, 2, 85–93. [Google Scholar] [CrossRef]
- Cadenas, S.; Echtay, K.S.; Harper, J.A.; Jekabsons, M.B.; Buckingham, J.A.; Grau, E.; Abuin, A.; Chapman, H.; Clapham, J.C.; Brand, M.D. The basal proton conductance of skeletal muscle mitochondria from transgenic mice overexpressing or lacking uncoupling protein-3. J. Biol. Chem. 2002, 277, 2773–2778. [Google Scholar] [CrossRef] [PubMed]
- Couplan, E.; del Mar Gonzalez-Barroso, M.; Alves-Guerra, M.C.; Ricquier, D.; Goubern, M.; Bouillaud, F. No evidence for a basal, retinoic, or superoxide-induced uncoupling activity of the uncoupling protein 2 present in spleen or lung Mitochondria. J. Biol. Chem. 2002, 277, 26268–26275. [Google Scholar] [CrossRef]
- Pohl, E.E.; Rupprecht, A.; Macher, G.; Hilse, K.E. Important Trends in UCP3 Investigation. Front. Physiol. 2019, 10, 470. [Google Scholar] [CrossRef]
- Korshunov, S.S.; Skulachev, V.P.; Starkov, A.A. High protonic potential actuates a mechanism of production of reactive oxygen species in mitochondria. FEBS Lett. 1997, 416, 15–18. [Google Scholar] [CrossRef]
- Plotnikov, E.Y.; Zorov, D.B. Pros and cons of use of mitochondria-targeted antioxidants. Antioxidants 2019, 8, 316. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Cunha, F.M.; Caldeira da Silva, C.C.; Cerqueira, F.M.; Kowaltowski, A.J. Mild mitochondrial uncoupling as a therapeutic strategy. Curr. Drug Targets 2011, 12, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, E.Y.; Silachev, D.N.; Jankauskas, S.S.; Rokitskaya, T.I.; Chupyrkina, A.A.; Pevzner, I.B.; Zorova, L.D.; Isaev, N.K.; Antonenko, Y.N.; Skulachev, V.P.; et al. Mild uncoupling of respiration and phosphorylation as a mechanism providing nephro- and neuroprotective effects of penetrating cations of the SkQ family. Biochemistry (Moscow) 2012, 77, 1029–1037. [Google Scholar] [CrossRef]
- Chernyak, B.V.; Antonenko, Y.N.; Domnina, L.V.; Ivanova, O.Y.; Lyamzaev, K.G.; Pustovidko, A.V.; Rokitskaya, T.I.; Severina, I.I.; Simonyan, R.A.; Trendeleva, T.A.; et al. Novel penetrating cations for targeting mitochondria. Curr. Pharm. Des. 2013, 19, 2795–2806. [Google Scholar] [CrossRef]
- Khailova, L.S.; Silachev, D.N.; Rokitskaya, T.I.; Avetisyan, A.V.; Lyamsaev, K.G.; Severina, I.I.; Il’yasova, T.M.; Gulyaev, M.V.; Dedukhova, V.I.; Trendeleva, T.A.; et al. A short-chain alkyl derivative of rhodamine 19 acts as a mild uncoupler of mitochondria and a neuroprotector. Biochim. Biophys. Acta (BBA)—Bioenerg. 2014, 1837, 1739–1747. [Google Scholar] [CrossRef]
- Antonenko, Y.N.; Denisov, S.S.; Silachev, D.N.; Khailova, L.S.; Jankauskas, S.S.; Rokitskaya, T.I.; Danilina, T.I.; Kotova, E.A.; Korshunova, G.A.; Plotnikov, E.Y.; et al. A long-linker conjugate of fluorescein and triphenylphosphonium as mitochondria-targeted uncoupler and fluorescent neuro- and nephroprotector. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2016, 1860, 2463–2473. [Google Scholar] [CrossRef] [PubMed]
- Antonenko, Y.N.; Avetisyan, A.V.; Cherepanov, D.A.; Knorre, D.A.; Korshunova, G.A.; Markova, O.V.; Ojovan, S.M.; Perevoshchikova, I.V.; Pustovidko, A.V.; Rokitskaya, T.I.; et al. Derivatives of rhodamine 19 as mild mitochondria-targeted cationic uncouplers. J. Biol. Chem. 2011, 286, 17831–17840. [Google Scholar] [CrossRef] [PubMed]
- Kalinovich, A.V.; Shabalina, I.G. Novel mitochondrial cationic uncoupler C4R1 is an effective treatment for combating obesity in mice. Biochemistry (Moscow) 2015, 80, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Khailova, L.S.; Nazarov, P.A.; Sumbatyan, N.V.; Korshunova, G.A.; Rokitskaya, T.I.; Dedukhova, V.I.; Antonenko, Y.N.; Skulachev, V.P. Uncoupling and toxic action of alkyltriphenylphosphonium cations on mitochondria and the bacterium Bacillus Subtilis as a function of alkyl chain length. Biochemistry (Moscow) 2015, 80, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Goleva, T.N.; Rogov, A.G.; Korshunova, G.A.; Trendeleva, T.A.; Mamaev, D.V.; Aliverdieva, D.A.; Zvyagilskaya, R.A. SkQThy, a novel and promising mitochondria-targeted antioxidant. Mitochondrion 2019, 49, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Rogov, A.G.; Trendeleva, T.A.; Aliverdieva, D.A.; Zvyagilskaya, R.A. More about Interactions of Rhodamine 19 Butyl Ester with Rat Liver Mitochondria. Biochemistry (Moscow) 2016, 81, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Rogov, A.G.; Goleva, T.N.; Trendeleva, T.A.; Ovchenkova, A.P.; Aliverdieva, D.A.; Zvyagilskaya, R.A. New Data on Effects of SkQ1 and SkQT1 on Rat Liver Mitochondria and Yeast Cells. Biochemistry (Moscow) 2018, 83, 552–561. [Google Scholar] [CrossRef]
- Chernyak, B.V.; Antonenko, Y.N.; Galimov, E.R.; Domnina, L.V.; Dugina, V.B.; Zvyagilskaya, R.A.; Ivanova, O.Y.; Izyumov, D.S.; Lyamzaev, K.G.; Pustovidko, A.V.; et al. Novel mitochondria-targeted compounds composed of natural constituents: Conjugates of plant alkaloids berberine and palmatine with plastoquinone. Biochemistry (Moscow) 2012, 77, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Rokitskaya, T.I.; Nazarov, P.A.; Golovin, A.V.; Antonenko, Y.N. Blocking of Single α-Hemolysin Pore by Rhodamine Derivatives. Biophys. J. 2017, 112, 2327–2335. [Google Scholar] [CrossRef]
- Antonenko, Y.N.; Avetisyan, A.V.; Bakeeva, L.E.; Chernyak, B.V.; Chertkov, V.A.; Domnina, L.V.; Ivanova, O.Y.; Izyumov, D.S.; Khailova, L.S.; Klishin, S.S.; et al. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 1. Cationic plastoquinone derivatives: Synthesis and in vitro studies. Biochemistry (Moscow) 2008, 73, 1273–1287. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Sumbatyan, N.V.; Tashlitsky, V.N.; Korshunova, G.A.; Antonenko, Y.N.; Skulachev, V.P. Mitochondria-targeted penetrating cations as carriers of hydrophobic anions through lipid membranes. Biochim. Biophys. Acta (BBA)—Biomembr. 2010, 1798, 1698–1706. [Google Scholar] [CrossRef]
- Gear, A.R. Rhodamine 6G. A Potent Inhibitor of Mitochondrial Oxidative Phosphorylation. J. Biol. Chem. 1974, 249, 3628–3637. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Ilyasova, T.M.; Severina, I.I.; Antonenko, Y.N.; Skulachev, V.P. Electrogenic Proton Transport across Lipid Bilayer Membranes Mediated by Cationic Derivatives of Rhodamine 19: Comparison with Anionic Protonophores. Eur. Biophys. J. 2013, 42, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Lardy, H. Isolation of liver or kidney mitochondria. Methods Enzymol. 1967, 10, 94–96. [Google Scholar]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar] [CrossRef] [PubMed]
- Kovalyov, L.I.; Kovalyova, M.A.; Kovalyov, P.L.; Serebryakova, M.V.; Moshkovskii, S.A.; Shishkin, S.S. Polymorphism of Δ3,5-Δ2,4-dienoyl-coenzyme A isomerase (the ECH1 gene product protein) in human striated muscle tissue. Biochemistry (Moscow) 2006, 71, 448–453. [Google Scholar] [CrossRef]
- Sims, P.J.; Waggoner, A.S.; Wang, C.H.; Hoffman, J.F. Mechanism by which cyanine dyes measure membrane potential in red blood cells and phosphatidylcholine vesicles. Biochemistry 1974, 13, 3315–3330. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.B.; Koshland, D.E. Sensory electrophysiology of bacteria: Relationship of the membrane potential to motility and chemotaxis in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 1977, 74, 4752–4756. [Google Scholar] [CrossRef]
- Letellier, L.; Shechter, E. Cyanine dye as monitor of membrane potentials in Escherichia coli cells and membrane vesicles. Eur. J. Biochem. 1979, 102, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Kobrinsky, E.; Zorov, D.B.; Aon, M.A.; Cortassa, S.; Sollott, S.J. Setting the record straight: A new twist on the chemiosmotic mechanism of oxidative phosphorylation. Function 2022, 3, zqac018. [Google Scholar] [CrossRef]
- Brand, M.D. The efficiency and plasticity of mitochondrial energy transduction. Biochem. Soc. Trans. 2005, 33, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Zorov, D.B.; Yaniv, Y.; Nuss, H.B.; Wang, S.; Sollott, S.J. Role of glycogen synthase kinase-3β in cardioprotection. Circ. Res. 2009, 104, 1240–1252. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Zorov, D.B.; Kim, S.H.; Pepe, S.; Fu, Q.; Fishbein, K.W.; Ziman, B.D.; Wang, S.; Ytrehus, K.; Antos, C.L.; et al. Glycogen synthase kinase-3β mediates convergence of protection signaling to inhibit the mitochondrial permeability transition pore. J. Clin. Investig. 2004, 113, 1535–1549. [Google Scholar] [CrossRef]
- Queliconi, B.B.; Marazzi, T.B.M.; Vaz, S.M.; Brookes, P.S.; Nehrke, K.; Augusto, O.; Kowaltowski, A.J. Bicarbonate modulates oxidative and functional damage in ischemia–reperfusion. Free. Radic. Biol. Med. 2013, 55, 46–53. [Google Scholar] [CrossRef]
- Kumar, S.; Flacke, J.P.; Kostin, S.; Appukuttan, A.; Reusch, H.P.; Ladilov, Y. SLC4A7 sodium bicarbonate co-transporter controls mitochondrial apoptosis in ischaemic coronary endothelial cells. Cardiovasc. Res. 2011, 89, 392–400. [Google Scholar] [CrossRef]
- Alka, K.; Casey, J.R. Bicarbonate transport in health and disease. IUBMB Life 2014, 66, 596–615. [Google Scholar] [CrossRef] [PubMed]
- Acin-Perez, R.; Salazar, E.; Kamenetsky, M.; Buck, J.; Levin, L.R.; Manfredi, G. Cyclic AMP produced inside mitochondria regulates oxidative phosphorylation. Cell Metab. 2009, 9, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Zeylemaker, W.P.; Klaasse, A.D.M.; Slater, E.C.; Veeger, C. Studies on succinate dehydrogenase. VI. Inhibition by monocarboxylic acids. Biochim. Biophys. Acta (BBA)—Enzymol. 1970, 198, 415–422. [Google Scholar] [CrossRef]
- Kasho, V.N.; Boyer, P.D. Relationships of inosine triphosphate and bicarbonate effects on F1 ATPase to the binding change mechanism. J. Bioenerg. Biomembr. 1984, 16, 407–419. [Google Scholar] [CrossRef]
- Roveri, O.A.; Calcaterra, N.B. Steady-state kinetics of F1 -ATPase. FEBS Lett. 1985, 192, 123–127. [Google Scholar] [CrossRef]
- Khailova, L.S.; Vygodina, T.V.; Lomakina, G.Y.; Kotova, E.A.; Antonenko, Y.N. Bicarbonate suppresses mitochondrial membrane depolarization induced by conventional uncouplers. Biochem. Biophys. Res. Commun. 2020, 530, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Alavian, K.N.; Beutner, G.; Lazrove, E.; Sacchetti, S.; Park, H.-A.; Licznerski, P.; Li, H.; Nabili, P.; Hockensmith, K.; Graham, M.; et al. An uncoupling channel within the c-subunit ring of the F1FO ATP synthase is the mitochondrial permeability transition pore. Proc. Natl. Acad. Sci. USA 2014, 111, 10580–10585. [Google Scholar] [CrossRef]
- Mnatsakanyan, N.; Park, H.A.; Wu, J.; He, X.; Llaguno, M.C.; Latta, M.; Miranda, P.; Murtishi, B.; Graham, M.; Weber, J.; et al. Mitochondrial ATP synthase C-subunit leak channel triggers cell death upon loss of its F1 subcomplex. Cell Death Differ. 2022, 29, 1874–1887. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Kobrinsky, E.; Zorov, D.B.; Nuss, H.B.; Yaniv, Y.; Fishbein, K.W.; de Cabo, R.; Montoliu, L.; Gabelli, S.B.; Aon, M.A.; et al. ATP Synthase K+- and H+-Fluxes Drive ATP Synthesis and Enable Mitochondrial K+-“Uniporter” Function: I. Characterization of Ion Fluxes. Function 2022, 3, zqab065. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Kobrinsky, E.; Zorov, D.B.; Nuss, H.B.; Yaniv, Y.; Fishbein, K.W.; de Cabo, R.; Montoliu, L.; Gabelli, S.B.; Aon, M.A.; et al. ATP Synthase K+- and H+-Fluxes Drive ATP Synthesis and Enable Mitochondrial K+-“Uniporter” Function: II. Ion and ATP Synthase Flux Regulation. Function 2022, 3, zqac001. [Google Scholar] [CrossRef] [PubMed]
- Chinopoulos, C. Mitochondrial permeability transition pore: Back to the drawing board. Neurochem. Int. 2018, 117, 49–54. [Google Scholar] [CrossRef]
- Hatefi, Y.; Hanstein, W.G.; Galante, Y.; Stiggall, D.L. Mitochondrial ATP-Pi exchange complex and the site of uncoupling of oxidative phosphorylation. Fed. Proc. 1975, 34, 1699–1706. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorova, L.D.; Pevzner, I.B.; Khailova, L.S.; Korshunova, G.A.; Kovaleva, M.A.; Kovalev, L.I.; Serebryakova, M.V.; Silachev, D.N.; Sudakov, R.V.; Zorov, S.D.; et al. Mitochondrial ATP Synthase and Mild Uncoupling by Butyl Ester of Rhodamine 19, C4R1. Antioxidants 2023, 12, 646. https://doi.org/10.3390/antiox12030646
Zorova LD, Pevzner IB, Khailova LS, Korshunova GA, Kovaleva MA, Kovalev LI, Serebryakova MV, Silachev DN, Sudakov RV, Zorov SD, et al. Mitochondrial ATP Synthase and Mild Uncoupling by Butyl Ester of Rhodamine 19, C4R1. Antioxidants. 2023; 12(3):646. https://doi.org/10.3390/antiox12030646
Chicago/Turabian StyleZorova, Ljubava D., Irina B. Pevzner, Ljudmila S. Khailova, Galina A. Korshunova, Marina A. Kovaleva, Leonid I. Kovalev, Marina V. Serebryakova, Denis N. Silachev, Roman V. Sudakov, Savva D. Zorov, and et al. 2023. "Mitochondrial ATP Synthase and Mild Uncoupling by Butyl Ester of Rhodamine 19, C4R1" Antioxidants 12, no. 3: 646. https://doi.org/10.3390/antiox12030646
APA StyleZorova, L. D., Pevzner, I. B., Khailova, L. S., Korshunova, G. A., Kovaleva, M. A., Kovalev, L. I., Serebryakova, M. V., Silachev, D. N., Sudakov, R. V., Zorov, S. D., Rokitskaya, T. I., Popkov, V. A., Plotnikov, E. Y., Antonenko, Y. N., & Zorov, D. B. (2023). Mitochondrial ATP Synthase and Mild Uncoupling by Butyl Ester of Rhodamine 19, C4R1. Antioxidants, 12(3), 646. https://doi.org/10.3390/antiox12030646