
The Role of Seminal Oxidative Stress in Recurrent Pregnancy Loss

Abstract
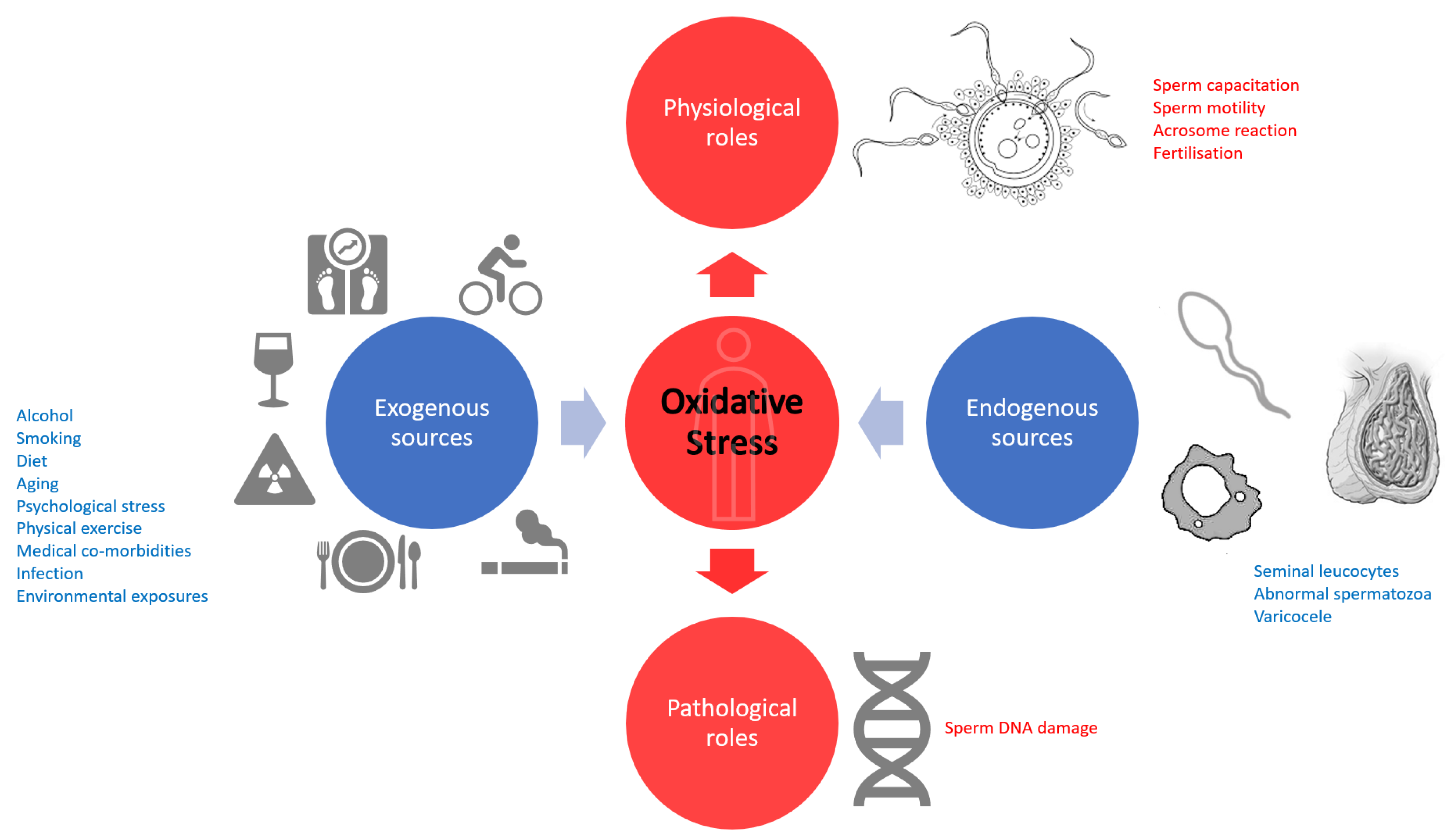
:1. Introduction
2. Spermatic Function
3. Seminal Redox Balance
4. Assessment of Male Productive Function
5. Elevated Seminal ROS and RPL
6. Sperm DNA Damage and RPL
7. Sperm Protein Expression and RPL
8. Use of Antioxidants
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rai, R.; Regan, L. Recurrent Miscarriage. Lancet 2006, 368, 601–611. [Google Scholar] [CrossRef] [PubMed]
- American Society for Reproductive Medicine. Practice Committee Report: Patient’s Fact Sheet: Recurrent Pregnancy Loss. Available online: https://www.asrm.org/topics/topics-index/miscarriage-or-recurrent-pregnancy-loss/ (accessed on 1 December 2022).
- The ESHRE Guideline Group on RPL. ESHRE guideline: Recurrent pregnancy loss. Hum. Reprod. Open 2018, 2, hoy004. Available online: https://www.eshre.eu/Guidelines-and-Legal/Guidelines/Recurrent-pregnancy-loss.aspx (accessed on 1 December 2022).
- Royal College Obstetricians and Gynaecologists (RCOG). The Investigation and Treatment of Couples with Recurrent First-Trimester and Second-Trimester Mis-Carriage; RCOG: London, UK, 2011; Volume 17, pp. 1–18. Available online: https://www.rcog.org.uk/guidance/browse-all-guidance/green-top-guidelines/the-investigation-and-treatment-of-couples-with-recurrent-miscarriage-green-top-guideline-no-17/ (accessed on 1 December 2022).
- Gupta, S.; Agarwal, A.; Banerjee, J.; Alvarez, J.G. The role of oxidative stress in spontaneous abortion and recurrent pregnancy loss: A systematic review. Obstet. Gynecol. Surv. 2007, 62, 334–335. [Google Scholar] [CrossRef] [Green Version]
- Gil-Villa, A.M.; Cardona-Maya, W.D.; Cadavid Jaramillo, A.P. Early embryo death: Does the male factor play a role? Arch. Esp. Urol. 2007, 60, 1057–1068. [Google Scholar]
- Puscheck, E.E.; Jeyendran, R.S. The impact of male factor on recurrent pregnancy loss. Curr. Opin. Obstet. Gynecol. 2007, 19, 222–228. [Google Scholar] [CrossRef]
- Aitken, R. Possible redox regulation of sperm motility activation. J. Androl. 2000, 21, 491–496. [Google Scholar] [PubMed]
- Lewis, S.E.M.; John Aitken, R.; Conner, S.J.; Iuliis, G.D.; Evenson, D.P.; Henkel, R.; Giwercman, A.; Gharagozloo, P. The impact of sperm DNA damage in assisted conception and beyond: Recent advances in diagnosis and treatment. Reprod. Biomed. Online 2013, 27, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Ikawa, M.; Inoue, N.; Benham, A.M.; Okabe, M. Fertilization: A sperm’s journey to and interaction with the oocyte. J. Clin. Investig. 2010, 120, 984–994. [Google Scholar] [CrossRef] [Green Version]
- Jena, S.R.; Nayak, J.; Kumar, S.; Kar, S.; Dixit, A.; Samanta, L. Paternal contributors in recurrent pregnancy loss: Cues from comparative proteome profiling of seminal extracellular vesicles. Mol. Reprod. Dev. 2021, 88, 96–112. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.; Jodar, M.; Oliva, R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum. Reprod. Update 2018, 24, 535–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brevini, T.A.; Georgia, P. Gametogenesis, Early Embryo Development and Stem Cell Derivation; Springer: New York, NY, USA, 2013. [Google Scholar]
- Agarwal, A.; Allamaneni, S.S. The effect of sperm DNA damage on assisted reproduction outcomes. A review. Minerva Ginecol. 2004, 56, 235–245. [Google Scholar] [PubMed]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Gosalvez, J.; Tvrda, E.; Agarwal, A. Free radical and superoxide reactivity detection in semen quality assessment: Past, present, and future. J. Assist. Reprod. Genet. 2017, 34, 697–707. [Google Scholar] [CrossRef]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef] [Green Version]
- Zini, A.; Defreitas, G.; Freeman, M.; Hechter, S.; Jarvi, K. Varicocele is associated with abnormal retention of cytoplasmic droplets by human spermatozoa. Fertil. Steril. 2000, 74, 461–464. [Google Scholar] [CrossRef]
- Park, J.; Rho, H.K.; Kim, K.H.; Choe, S.S.; Lee, Y.S.; Kim, J.B. Overexpression of glucose-6-phosphate dehydrogenase is associated with lipid dysregulation and insulin resistance in obesity. Mol. Cell. Biol. 2005, 25, 5146–5157. [Google Scholar] [CrossRef] [Green Version]
- Frederiks, W.M.; Vreeling-Sindelárová, H. Localization of glucose-6-phosphate dehydrogenase activity on ribosomes of granular endoplasmic reticulum, in peroxisomes and peripheral cytoplasm of rat liver parenchymal cells. Histochem. J. 2001, 33, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Sanocka, D.; Kurpisz, M. Reactive oxygen species and sperm cells. Reprod. Biol. Endocrinol. 2004, 2, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, C.; Obert, G.; Deffosez, A.; Formstecher, P.; Marchetti, P. Study of mitochondrial membrane potential, reactive oxygen species, DNA fragmentation and cell viability by flow cytometry in human sperm. Hum. Reprod. 2002, 17, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Said, T.M.; Agarwal, A.; Sharma, R.K.; Thomas, A.J., Jr.; Sikka, S.C. Impact of sperm morphology on DNA damage caused by oxidative stress induced by beta-nicotinamide adenine dinucleotide phosphate. Fertil. Steril. 2005, 83, 95–103. [Google Scholar] [CrossRef]
- Morielli, T.; O’Flaherty, C. Oxidative stress impairs function and increases redox protein modifications in human spermatozoa. Reproduction 2015, 49, 113–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamirande, E.; Gagnon, C. Reactive oxygen species and human spermatozoa. J. Androl. 1992, 13, 368–378. [Google Scholar]
- Fedder, J. Nonsperm cells in human semen: With special reference to seminal leukocytes and their possible influence on fertility. Syst. Biol. Reprod. Med. 1996, 36, 41–65. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Makker, K.; Sharma, R. Clinical relevance of oxidative stress in male factor infertility: An update. Am. J. Reprod. Immunol. 2008, 59, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Muzio, M.; Bosisio, D.; Polentarutti, N.; D’amico, G.; Stoppacciaro, A.; Mancinelli, R.; Veer, C.; Penton-Rol, G.; Ruco, L.P.; Allavena, P.; et al. Differential expression and regulation of toll-like receptors (TLR) in human leukocytes: Selective expression of TLR3 in dendritic cells. J. Immunol. 2000, 164, 5998–6004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talebi, A.R.; Sarcheshmeh, A.A.; Khalili, M.A.; Tabibnejad, N. Effects of ethanol consumption on chromatin condensation and DNA integrity of epididymal spermatozoa in rat. Alcohol 2011, 45, 403–409. [Google Scholar] [CrossRef]
- Dietrich, M.; Block, G.; Norkus, E.P.; Hudes, M.; Traber, M.G.; Cross, C.E.; Packer, L. Smoking and exposure to environmental tobacco smoke decrease some plasma antioxidants and increase γ-tocopherol in vivo after adjustment for dietary antioxidant intakes. Am. J. Clin. Nutr. 2003, 77, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Kashou, A.H.; du Plessis, S.S.; Agarwal, A. The Role of Obesity in ROS Generation and Male Infertility. Studies on Men’s Health and Fertility; Springer: Berlin/Heidelberg, Germany, 2012; pp. 571–590. [Google Scholar]
- Moskovtsev, S.I.; Willis, J.; Mullen, J.B.M. Age-related decline in sperm deoxyribonucleic acid integrity in patients evaluated for male infertility. Fertil. Steril. 2006, 85, 496–949. [Google Scholar] [CrossRef]
- Eskiocak, S.; Gozen, A.S.; Kilic, A.S.; Molla, S. Association between mental stress & some antioxidant enzymes of seminal plasma. Indian J. Med. Res. 2005, 122, 491–496. [Google Scholar]
- Peake, J.M.; Suzuki, K.; Coombes, J.S. The influence of antioxidant supplementation on markers of inflammation and the relationship to oxidative stress after exercise. J. Nutr. Biochem. 2007, 18, 357–371. [Google Scholar] [CrossRef]
- Vignera, S.; Condorelli, R.; Vicari, E.; D’Agata, R.; Calogero, A.E. Diabetes mellitus and sperm parameters. J. Androl. 2012, 33, 145–153. [Google Scholar] [CrossRef]
- Agarwal, A.; Prabakaran, S.; Allamaneni, S.S. Relationship between oxidative stress, varicocele and infertility: A meta-analysis. Reprod. BioMed. Online 2006, 12, 630–633. [Google Scholar] [CrossRef]
- Sies, H. Strategies of antioxidant defence. FEBS J. 1993, 215, 213–219. [Google Scholar]
- Yeung, C.; Cooper, T.; De Geyter, M.; Rolf, C.; Kamischke, A.; Nieschlag, E. Studies on the origin of redox enzymes in seminal plasma and their relationship with results of in-vitro fertilization. Mol. Hum. Reprod. 1998, 4, 835–839. [Google Scholar] [CrossRef]
- Agarwal, A.; Durairajanayagam, D.; Halabi, J.; Peng, J.; VazquezLevin, M. Proteomics, oxidative stress and male infertility. Reprod. Biomed. Online 2014, 29, 32–58. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Sharma, R.K.; Nallella, K.P.; Thomas, A.J., Jr.; Alvarez, J.G.; Sikka, S.C. Reactive oxygen species as an independent marker of male factor infertility. Fertil. Steril. 2006, 86, 878–885. [Google Scholar] [CrossRef]
- Tremellen, K. Treatment of sperm oxidative stress: A collaborative approach between clinician and embryologist. In Oxidants, Antioxidants, and Impact of the Oxidative Status in Male Reproduction; Henkel, R., Samanta, L., Agarwal, A., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press: London, UK, 2018; pp. 225–235. [Google Scholar]
- Agarwal, A.; Allamaneni, S.S. Free radicals and male reproduction. J. Indian Med. Assoc. 2011, 109, 184–187. [Google Scholar]
- Agarwal, A.; Mulgund, A.; Sharma, R.; Sabanegh, E. Mechanisms of oligozoospermia: An oxidative stress perspective. Syst. Biol. Reprod. Med. 2014, 60, 206–221. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Parekh, N.; Panner Selvam, M.K.; Henkel, R.; Shah, R.; Homa, S.T.; Ramasamy, R.; Ko, E.; Tremellen, K.; Esteves, S.; et al. Male Oxidative Stress Infertility (MOSI): Proposed Terminology and Clinical Practice Guidelines for Management of Idiopathic Male Infertility. World J. Mens Health 2019, 37, 296–312. [Google Scholar] [CrossRef]
- Kamkar, N.; Ramezanali, F.; Sabbaghian, M. The relationship between sperm DNA fragmentation, free radicals and antioxidant capacity with idiopathic repeated pregnancy loss. Reprod. Biol. 2018, 18, 330–335. [Google Scholar] [CrossRef]
- Evenson, D.P.; Jost, L.K.; Marshall, D.; Zinaman, M.J.; Clegg, E.; Purvis, K.; Angelis, P.; Claussen, O.P. Utility of the sperm chromatin structure assay as a diagnostic and prog-nostic tool in the human fertility clinic. Hum. Reprod. 1999, 14, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Larson, K.L.; DeJonge, C.J.; Barnes, A.M.; Jost, L.K.; Evenson, D.P. Sperm chro- matin structure assay parameters as predictors of failed pregnancy following assisted reproductive techniques. Hum. Reprod. 2000, 15, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Carrell, D.T.; Liu, L.; Peterson, C.M.; Jones, K.P.; Hatasaka, H.H.; Erickson, L.; Campbell, B. Sperm DNA fragmentation is increased in couples with unexplained recurrent pregnancy loss. Arch. Androl. 2003, 49, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Sergerie, M.; Laforest, G.; Bujan, L.; Bissonnette, F.; Bleau, G. Sperm DNA fragmentation: Threshold value in male fertility. Hum. Reprod. 2005, 20, 3446–3451. [Google Scholar] [CrossRef]
- Caglar, G.S.; Koster, F.; Schopper, B.; Asimakopoulos, B.; Nehls, B.; Nikolettos, N.; Diedrich, K.; Al-Hasani, S. Semen DNA fragmentation index, evaluated with both TUNEL and comet assay, and the ICSI outcome. In Vivo 2007, 21, 1075–1080. [Google Scholar]
- Vessey, W.; Perez-Miranda, A.; Macfarquhar, R.; Agarwal, A.; Homa, S. Reactive oxygen species in human semen: Validation and qualification of a chemiluminescence assay. Fertil. Steril. 2014, 102, 1576–1583.e4. [Google Scholar] [CrossRef]
- Castleton, P.E.; Deluao, J.C.; Sharkey, D.J.; McPherson, N.O. Measuring Reactive Oxygen Species in Semen for Male Preconception Care: A Scientist Perspective. Antioxidants 2022, 11, 264. [Google Scholar] [CrossRef]
- Gil-Villa, A.M.; Cardona-Maya, W.; Agarwal, A.; Sharma, R.; Cadavid, A. Assessment of sperm factors possibly involved in early recurrent pregnancy loss. Fertil. Steril. 2010, 94, 1465–1472. [Google Scholar] [CrossRef]
- Ghiselli, A.; Serafini, M.; Natella, F.; Scaccini, C. Total antioxidant capacity as a tool to assess redox status: Critical view and experimental data. Free Radic. Biol. Med. 2000, 29, 1106–1114. [Google Scholar] [CrossRef]
- Mohanty, G.; Swain, N.; Goswami, C.; Kar, S.; Samanta, L. Histone retention, protein carbonylation, and lipid peroxidation in spermatozoa: Possible role in recurrent pregnancy loss. Syst. Biol. Reprod. Med. 2016, 62, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, S.; Thilagavathi, J.; Kumar, K.; Deka, D.; Talwar, P.; Dada, R. Cytogenetic, Y chromosome microdeletion, sperm chromatin and oxidative stress analysis in male partners of couples experiencing recurrent spontaneous abortions. Arch. Gynecol. Obstet. 2011, 284, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Imam, S.; Shamsi, M.; Kumar, K.; Deka, D.; Dada, R. Idiopathic recurrent pregnancy loss: Role of paternal factors; a pilot study. J. Reprod. Infertil. 2011, 12, 267–276. [Google Scholar]
- Jayasena, C.N.; Radia, U.K.; Figueiredo, M.; Revill, L.F.; Dimakopoulou, A.; Osagie, M.; Vessey, W.; Regan, L.; Rai, R.; Dhillo, W. Reduced Testicular Steroidogenesis and Increased Semen Oxidative Stress in Male Partners as Novel Markers of Recurrent Miscarriage. Clin. Chem. 2019, 66, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Ghanaie, M.; Asgari, S.A.; Dadrass, N.; Allahkhah, A.; Iran-Pour, E.; Safarinejad, M.R. Effects of varicocele repair on spontaneous first trimester miscarriage: A randomized clinical trial. Urol. J. 2012, 9, 505–513. [Google Scholar]
- Negri, L.; Levi-Setti, P.E. Pregnancy rate after varicocele repair: How many miscarriages? J. Androl. 2011, 32, 1. [Google Scholar] [CrossRef]
- Bellver, J.; Meseguer, M.; Muriel, L.; Garcia-Herrero, S.; Barreto, M.A.; Garda, A.L.; Remohi, J.; Pellicer, A.; Garrido, N. Y chromosome microdeletions, sperm DNA fragmentation and sperm oxidative stress as causes of recurrent spontaneous abortion of unknown etiology. Hum. Reprod. 2010, 25, 1713–1721. [Google Scholar] [CrossRef] [Green Version]
- Ribas-Maynou, J.; Garcia-Piero, A.; Fernandez-Encinas, A.; Amengual, M.; Prada, E.; Cortes, P.; Navarro, J.; Benet, J. Double stranded sperm DNA breaks, measured by comet assay, are associated with unexplained recurrent miscarriage in couples without a female factor. PLoS ONE 2012, 7, e44679. [Google Scholar] [CrossRef] [Green Version]
- Ruizue, W.; Hongli, Z.; Zhihong, Z.; Rulin, D.; Dongfeng, G.; Ruizhi, L. The impact of semen quality, occupational exposure to environmental factors and lifestyle on recurrent pregnancy loss. J. Assist. Reprod. Genet. 2013, 30, 1513–1518. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, L.; Zhang, X.; Xu, G.; Zhang, W.; Wang, K.; Wang, Q.; Qiu, Y.; Li, J.; Gai, L. Sperm chromatin integrity may predict future fertility for unexplained spontaneous abortion patients. Int. J. Androl. 2012, 35, 752–757. [Google Scholar] [CrossRef]
- Zidi-Jrah, I.; Hajlaoui, A.; Mougou-Zerelli, S.; Kammoun, M.; Meniaoui, I.; Sallem, A.; Brahem, S.; Fekih, M.; Bibi, M.; Saad, A.; et al. Relationship between sperm aneuploidy, sperm DNA integrity, chro- matin packaging, traditional semen parameters, and recurrent pregnancy loss. Fertil. Steril. 2016, 105, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Khadem, N.; Poorhoseyni, A.; Jalali, M.; Akbary, A.; Heydari, S.T. Sperm DNA fragmentation in couplesl with unexplained recurrent spontaneous abortions. Andrologia 2014, 46, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, C.; Clarke, H.; Cutting, R.; Saxton, J.; Waite, S.; Ledger, W.; Li, T.; Pacey, A. Sperm DNA Fragmentation, recurrent implantation failure and recurrent miscarriage. Asian J. Androl. 2015, 17, 681–685. [Google Scholar]
- McQueen, D.B.; Zhang, J.; Robins, J.C. Sperm DNA fragmentation and recurrent pregnancy loss: A systematic review and meta-analysis. Fertil. Steril. 2019, 112, 54–60. [Google Scholar] [CrossRef]
- Absalan, F.; Ghannadi, A.; Kazerooni, M.; Parifar, R.; Jamalzadeh, F.; Amiri, S. Value of sperm chromatin dispersion test in couples with unexplained recurrent abortion. J. Assist. Reprod. Genet. 2012, 29, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Esquerre-Lamare, C.; Walschaerts, M.; Debordeaux, L.; Moreau, J.; Bretelle, F.; Isus, F.; Karsenty, G.; Monteil, L.; Perrin, J.; Papaxanthos-Roche, A.; et al. Sperm aneuploidy and DNA fragmentation in unexplained recur- rent pregnancy loss: A multicenter case-control study. Basic Clin. Androl. 2018, 28, 4. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Deka, D.; Singh, A.; Mitra, D.; Vanitha, B.; Dada, R. Predictive value of DNA integrity analysis in idiopathic recurrent pregnancy loss following spontaneous conception. J. Assist. Reprod. Genet. 2012, 29, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Bareh, G.; Jacoby, E.; Binkley, P.; Chang, T.; Schenken, R.; Robinson, R. Sperm Deoxyribonucleic Acid (DNA) Damage Assessment in Normozoospermic Male Partners of Couples with Unexplained Recurrent Pregnancy Loss: A Prospective Cohort Study. Fertil. Steril. 2016, 105, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Brahem, S.; Mehdi, M.; Landolsi, H.; Mougou, S.; Elghezal, H.; Saad, A. Semen parameters and sperm DNA fragmentation as causes of recurrent pregnancy loss. Urology 2011, 75, 792–796. [Google Scholar] [CrossRef]
- Bhattacharya, S. Association of various sperm parameters with unexplained repeated early pregnancy loss—Which is most important? Int. Urol. Nephrol. 2008, 40, 391–395. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, K.; Jain, S.; Hassan, T.; Dada, R. Novel insights into the genetic and epigenetic paternal contribution to the human embryo. Clinics 2013, 68 (Suppl. S1), 5–14. [Google Scholar] [CrossRef]
- Brunner, A.M.; Nanni, P.; Mansuy, I.M. Epigenetic marking of sperm by post-translational modification of histones and protamines. Epigenetics Chromatin 2014, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.; Amaral, A.; Oliva, R. Sperm nuclear proteome and its epigenetic potential. Andrology 2014, 2, 326–338. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Erkek, S.; Hisano, M.; Liang, C.Y.; Gill, M.; Murr, R.; Dieker, J.; Schübeler, D.; van der Vlag, J.; Stadler, M.B.; Peters, A.H.F.M. Molecular determinants of nucleosome retention at CpG-rich sequences in mouse spermatozoa. Nat. Struct. Mol. Biol. 2013, 20, 868–875. [Google Scholar] [CrossRef] [Green Version]
- Dadoune, J.P. Expression of mammalian spermatozoal nucleoproteins. Microsc. Res. Tech. 2003, 61, 56–75. [Google Scholar] [CrossRef]
- Kazerooni, T.; Asadi, N.; Jadid, L.; Kazerooni, M.; Ghanadi, A.; Ghaffarpasand, F.; Kazerooni, Y.; Zolghadr, J. Evaluation of sperm’s chromatin quality with acridine orange test, chromomycin A3 and aniline blue staining in couples with unexplained recurrent abortion. J. Assist. Reprod. Genet. 2009, 26, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, G.; Jena, S.R.; Nayak, J.; Kar, S.; Samanta, L. Proteomic Signatures in Spermatozoa Reveal the Role of Paternal Factors in Recurrent Pregnancy Loss. World J. Mens Health 2020, 38, 103–114. [Google Scholar] [CrossRef]
- Mohanty, G.; Jena, S.R.; Nayak, J.; Kar, S.; Samanta, L. Quantitative proteomics decodes clusterin as a critical regulator of paternal factors responsible for impaired compensatory metabolic reprogramming in recurrent pregnancy loss. Andrologia 2020, 52, e13498. [Google Scholar] [CrossRef]
- Majzoub, A.; Agarwal, A. Systematic review of antioxidant types and doses in male infertility: Benefits on semen parameters, advanced sperm function, assisted reproduction and live-birth rate. Arab. J. Urol. 2018, 16, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Smits, R.M.; Mackenzie-Proctor, R.; Yazdani, A.; Stankiewicz, M.T.; Jordan, V.; Showell, M.G. Antioxidants for male subfertility. Cochrane Database Syst. Rev. 2019, 3, CD007411. [Google Scholar] [CrossRef]
- Steiner, A.Z.; Hansen, K.R.; Barnhart, K.T.; Cedars, M.I.; Legro, R.S.; Diamond, M.P.; Krawetz, S.A.; Usadi, R.; Baker, V.L.; Coward, R.M.; et al. Reproductive Medicine Network. The effect of antioxidants on male factor infertility: The Males, Antioxidants, and Infertility (MOXI) randomized clinical trial. Fertil. Steril. 2020, 113, 552–560. [Google Scholar] [CrossRef]
- Henkel, R.; Sandhu, I.S.; Agarwal, A. The excessive use of antioxidant therapy: A possible cause of male infertility? Andrologia 2018, 51, e13162. [Google Scholar] [CrossRef] [Green Version]
- Gil-Villa, A.M.; Cardona-Maya, W.; Agarwal, A.; Sharma, R.; Cadavid, A. Role of male factor in early recurrent embryo loss: Do antioxidants have any effect? Fertil. Steril. 2009, 92, 565–571. [Google Scholar] [CrossRef]
- Nazari, A.; Sabeti, P.; Pourmasumi, S. Comparison between sperm parameters and chromatin in recurrent pregnancy loss couples after antioxidant therapy. J. Fam. Med. Prim. Care 2020, 9, 597–601. [Google Scholar]


{kind=link}
| Author | Year | Recurrent Pregnancy Loss Group | Control Group | Sperm Preparation | Sperm DNA Damage * | Oxidative Stress | ||
|---|---|---|---|---|---|---|---|---|
| Definition | N= | Definition | N= | |||||
| Jayasena | 2019 | ≥3 consecutive losses at <20/40 | 50 | No co-morbidities | 33 | Fresh | SCD | Luminol-based chemiluminescence assay |
| Bellver | 2015 | ≥3 losses at 5–14/40 | 30 | Fertile + no history of RPL + normal karyotype + no co-morbidities | 30 | Fresh | SCD | Flow cytometric assay (OxyDNA) for sperm DNA oxidative damage |
| Imam | 2011 | ≥3 consecutive losses at <20/40 | 20 | ≥1 live birth | 20 | Frozen | SCSA | Luminol-based chemiluminescence assay ELISA for total antioxidative capacity (TAC) |
| Venkatesh | 2011 | ≥2 losses at <24/40 | 32 | ≥1 live birth | 20 | Frozen | SCSA | Luminol-based chemiluminescence assay |
| Author | Year | Recurrent Pregnancy Loss Group | Control Group | Sperm Preparation | Sperm DNA Damage * | ||
|---|---|---|---|---|---|---|---|
| Definition | N= | Definition | N= | ||||
| Jayasena | 2019 | ≥3 consecutive losses at <20/40 | 50 | No co-morbidities | 33 | Fresh | SCD |
| Esquerre-Lamare | 2018 | ≥3 losses at <12/40 | 33 | ≥1 live birth | 27 | Frozen with cryoprecipitate | SCSA and TUNEL |
| Zidi-Jrah | 2016 | ≥2 losses at <24/40 | 22 | ≥1 live birth | 20 | Washed then frozen | TUNEL |
| Bareh | 2016 | ≥2 losses at <20/40 | 26 | ≥1 live birth | 31 | Fresh | TUNEL |
| Bellver | 2015 | ≥3 losses at 5–14/40 | 30 | Fertile + no history of RPL + normal karyotype + no co-morbidities | 30 | Fresh | SCD |
| Coughlan | 2014 | ≥3 consecutive losses at <20/40 | 16 | ≥1 live birth | 7 | Density centrifugation gradient vs. fresh | SCD and TUNEL |
| Ruixue | 2013 | ≥3 losses at <12/40 | 68 | Current pregnancy | 63 | Fresh | Aniline blue |
| Khadem | 2012 | ≥3 losses at <20/40 | 30 | Currently pregnancy | 30 | Fresh | SCD |
| Ribas-Maynou | 2012 | ≥2 losses at <12/40 | 20 | ≥1 live birth | 25 | Frozen with cryoprecipitate | Comet and SCD |
| Kumar | 2012 | ≥3 losses at <20 | 45 | ≥1 live birth | 20 | Frozen | SCSA |
| Zhang | 2012 | ≥2 consecutive losses at <12/40 | 111 | ≥1 live birth | 30 | Fresh | SCD |
| Absalan | 2012 | ≥3 losses at <20/40 | 30 | Fertile + no history of RPL in partner | 30 | Fresh | SCD |
| Imam | 2011 | ≥3 consecutive losses at <20/40 | 20 | ≥1 live birth | 20 | Frozen | SCSA |
| Venkatesh | 2011 | ≥2 losses at <24/40 | 32 | ≥1 live birth | 20 | Frozen | SCSA |
| Brahem | 2011 | ≥2 consecutive losses at <24/40 | 31 | ≥1 live birth | 20 | Frozen | TUNEL |
| Gil-Villa | 2010 | ≥2 losses at <12/40 | 23 | Established recent fertility | 11 | Fresh | SCSA |
| Bhattacharya | 2008 | ≥2 consecutive losses at <8/40 | 74 | ≥1 live birth | 65 | Fresh | Acridine orange |
| Carrell | 2003 | ≥3 losses at <20/40 | 21 | ≥1 live birth | 26 | Frozen | SCD and TUNEL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davies, R.; Jayasena, C.N.; Rai, R.; Minhas, S. The Role of Seminal Oxidative Stress in Recurrent Pregnancy Loss. Antioxidants 2023, 12, 723. https://doi.org/10.3390/antiox12030723
Davies R, Jayasena CN, Rai R, Minhas S. The Role of Seminal Oxidative Stress in Recurrent Pregnancy Loss. Antioxidants. 2023; 12(3):723. https://doi.org/10.3390/antiox12030723
Chicago/Turabian StyleDavies, Rhianna, Channa N. Jayasena, Raj Rai, and Suks Minhas. 2023. "The Role of Seminal Oxidative Stress in Recurrent Pregnancy Loss" Antioxidants 12, no. 3: 723. https://doi.org/10.3390/antiox12030723
APA StyleDavies, R., Jayasena, C. N., Rai, R., & Minhas, S. (2023). The Role of Seminal Oxidative Stress in Recurrent Pregnancy Loss. Antioxidants, 12(3), 723. https://doi.org/10.3390/antiox12030723