
Osthole Prevents Heart Damage Induced by Diet-Induced Metabolic Syndrome: Role of Fructokinase (KHK)

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Protocol
2.3. Body Weight and Feeding
2.4. Systolic Blood Pressure (SBP)
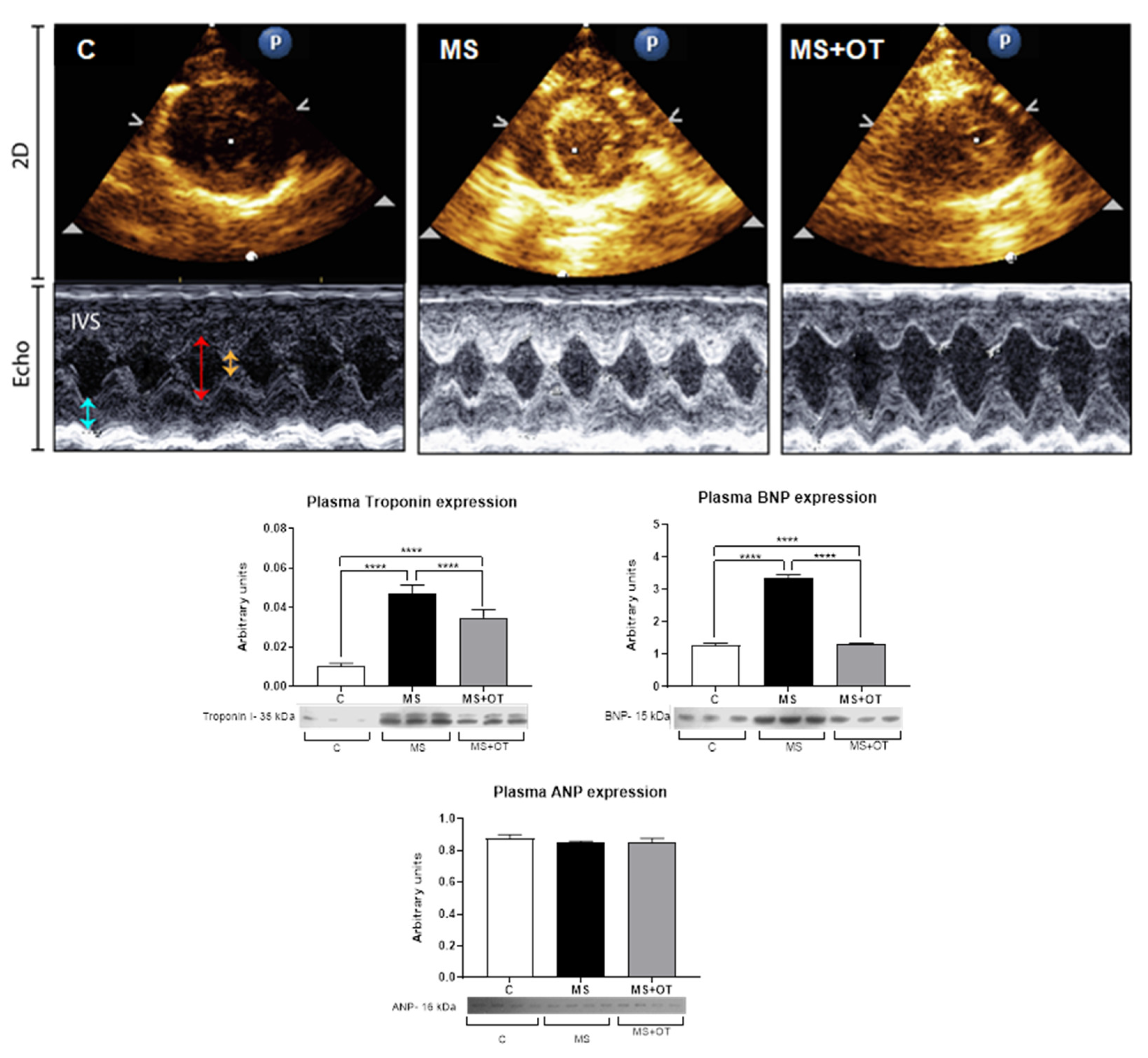
Evaluation of Cardiac Function by Echocardiography
2.5. Systemic Parameters
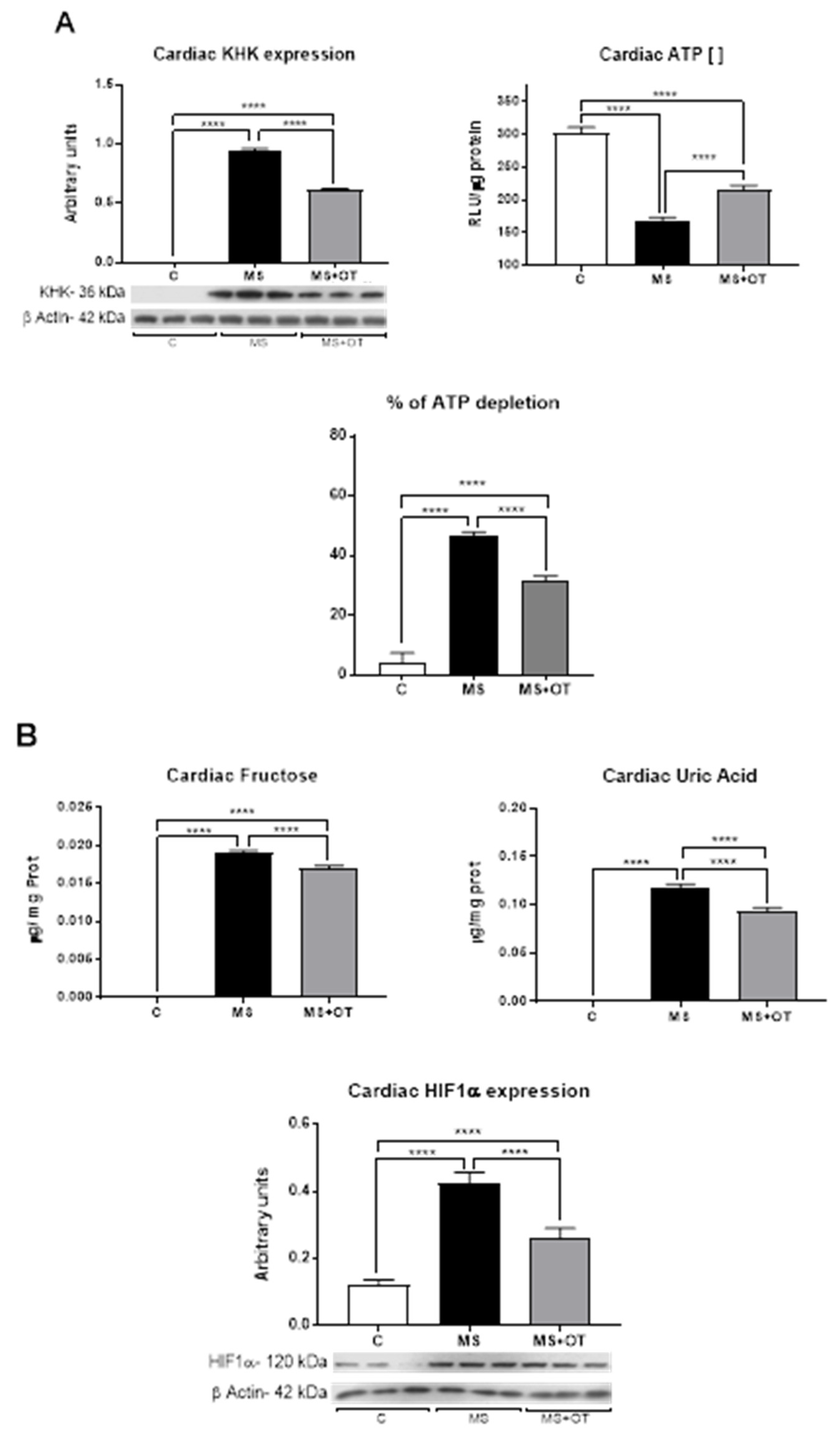
2.6. Heart Fructokinase (KHK) Activity Assessment
2.7. Heart Fructose and Uric Acid Content
2.8. Western Blot Analysis
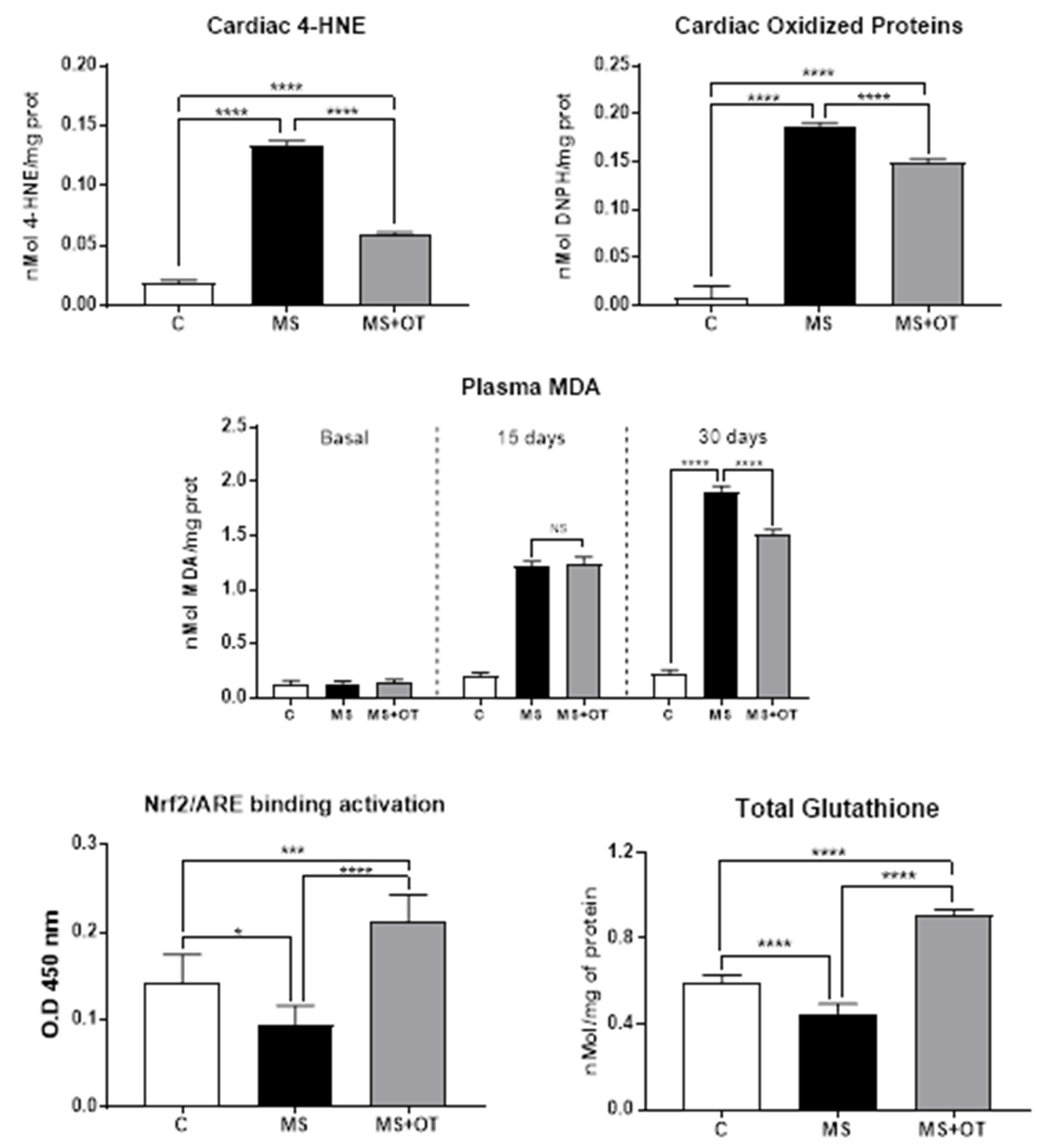
2.9. Oxidative Stress Markers
2.10. Nrf2 Factor Transcription Assay by Trans-Activation
2.11. Plasma Extracellular Vesicles Isolation
2.12. Histological Analysis
2.13. Immunohistochemistry
2.14. Statistical Analysis
3. Results
3.1. Body Weight and Fluid and Food Consumption
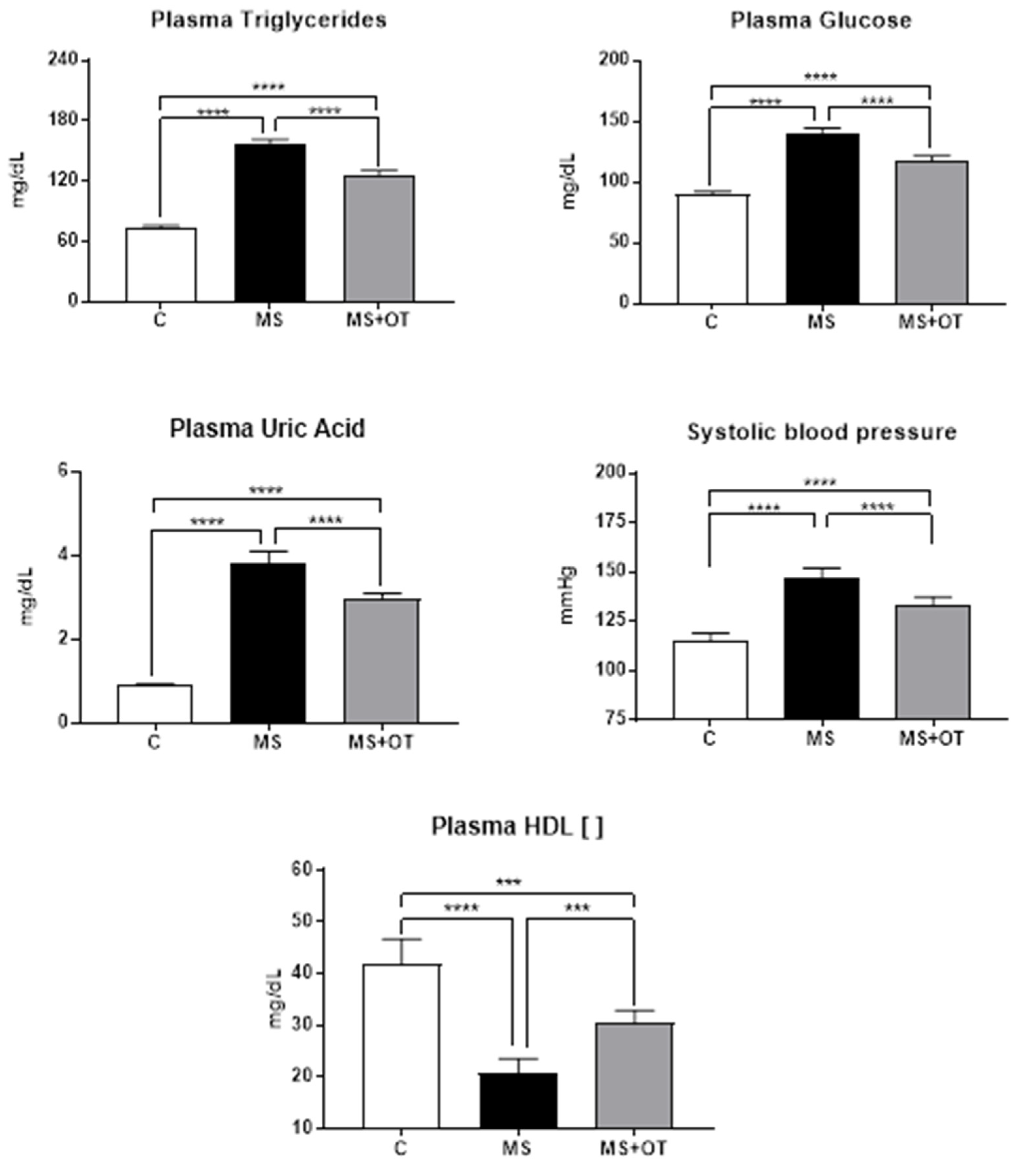
3.2. Markers of Metabolic Syndrome
3.3. Evaluation of Cardiac Function by Echocardiography and Systemic Markers of Cardiac Damage
3.4. Cardiac Oxidative Stress Assessment and Nrf2 Activation
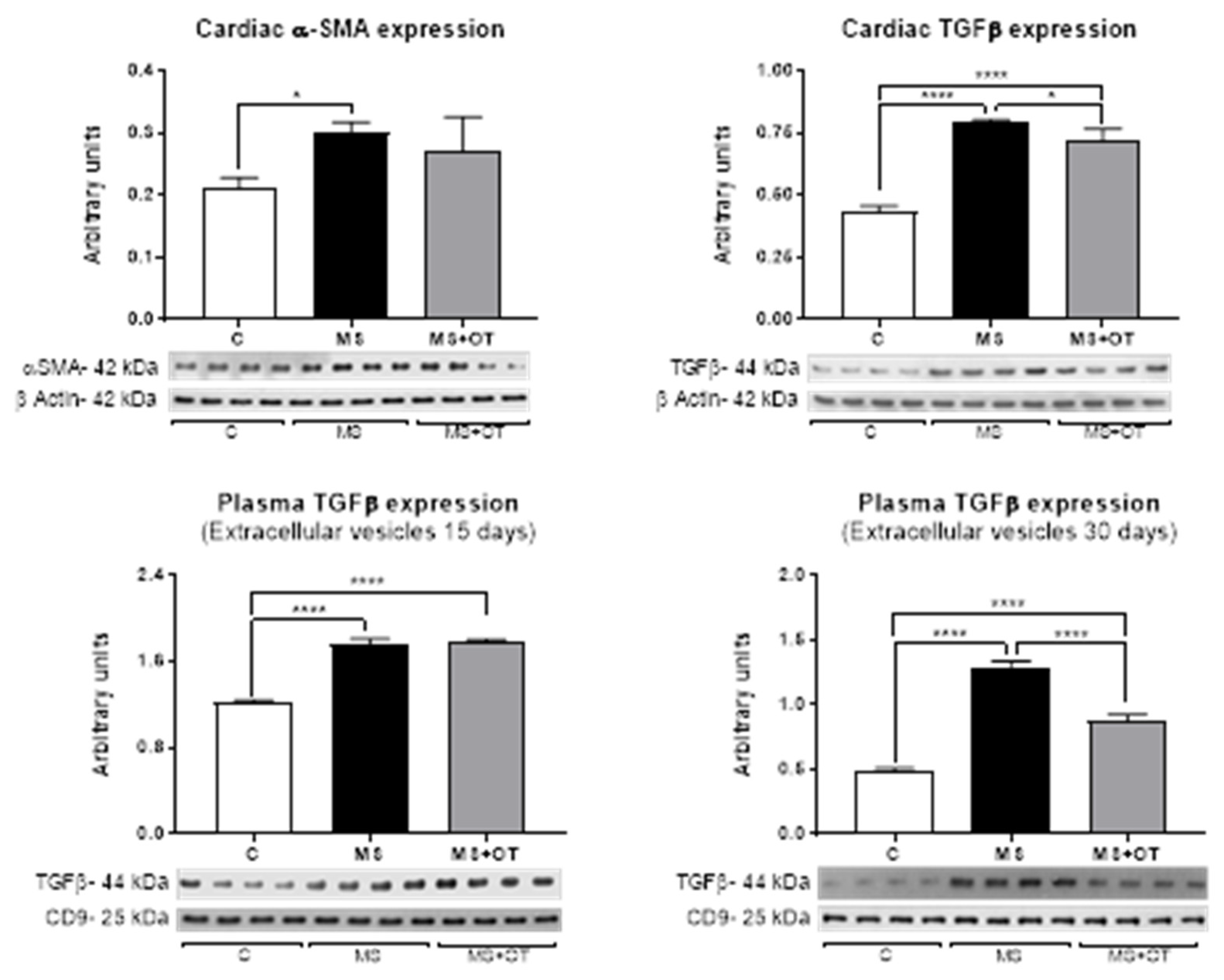
3.5. Fibrosis Assessment in Cardiac Tissue and Extracellular Vesicles
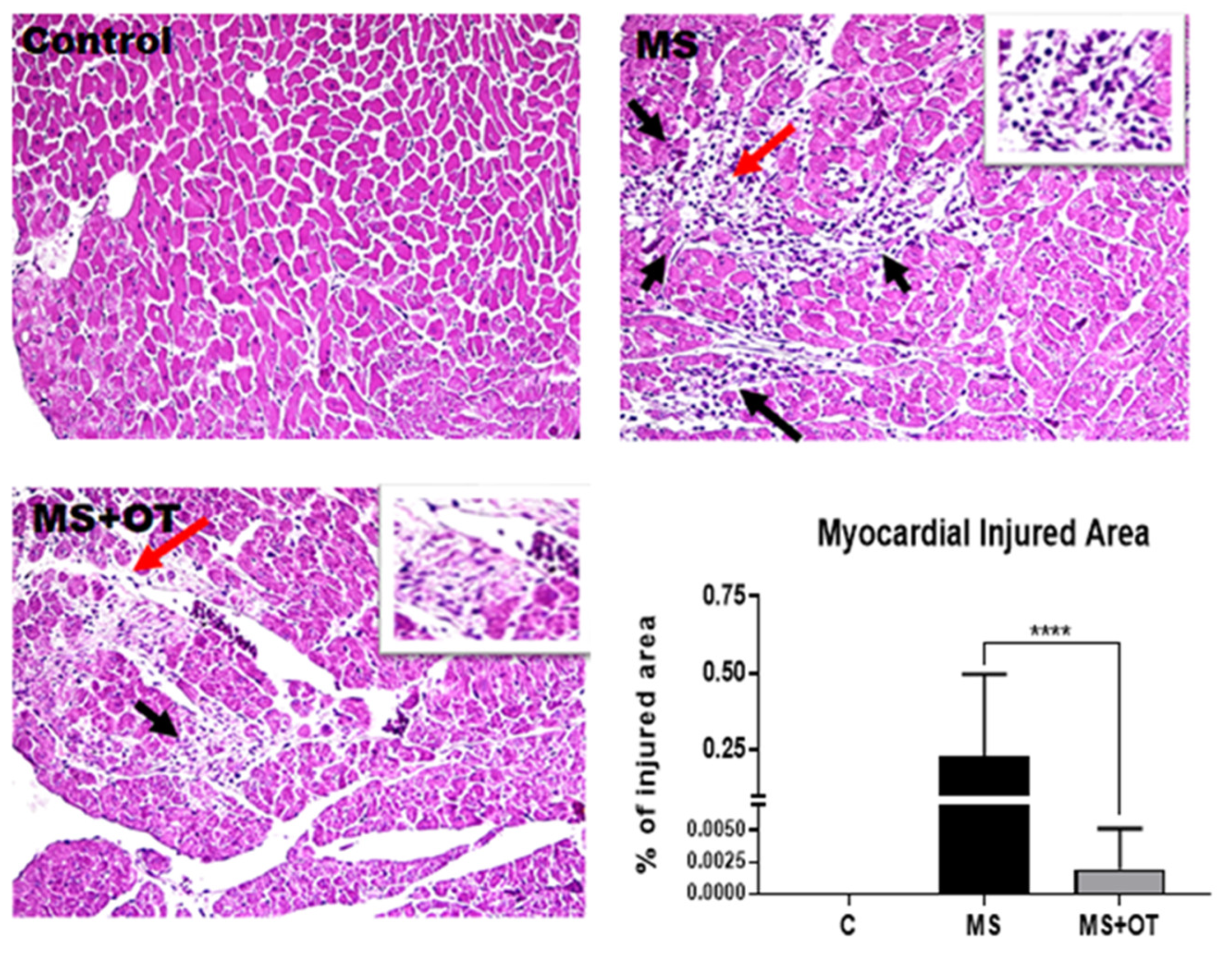
3.6. Histological Analysis
3.7. KHK Immunohistochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tune, J.D.; Goodwill, A.G.; Sassoon, D.J.; Mather, K.J. Cardiovascular consequences of metabolic syndrome. Transl. Res. 2017, 183, 57–70. [Google Scholar] [CrossRef]
- Annandale, M.; Daniels, L.J.; Li, X.; Neale, J.P.H.; Chau, A.H.L.; Ambalawanar, H.A.; James, S.L.; Koutsifeli, P.; Delbridge, L.M.D.; Mellor, K.M. Fructose Metabolism and Cardiac Metabolic Stress. Front. Pharmacol. 2021, 12, 695486. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.; Abdelmalek, M.F.; Sullivan, S.; Nadeau, K.J.; Green, M.; Roncal, C.; Nakagawa, T.; Kuwabara, M.; Sato, Y.; Kang, D.H.; et al. Fructose and sugar: A major mediator of non-alcoholic fatty liver disease. J. Hepatol. 2018, 68, 1063–1075. [Google Scholar] [CrossRef]
- Mellor, K.; Ritchie, R.H.; Meredith, G.; Woodman, O.L.; Morris, M.J.; Delbridge, L.M. High-fructose diet elevates myocardial superoxide generation in mice in the absence of cardiac hypertrophy. Nutrition 2010, 26, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Szucs, G.; Soja, A.; Peter, M.; Sarkozy, M.; Bruszel, B.; Siska, A.; Foldesi, I.; Szabo, Z.; Janaky, T.; Vigh, L.; et al. Prediabetes Induced by Fructose-Enriched Diet Influences Cardiac Lipidome and Proteome and Leads to Deterioration of Cardiac Function prior to the Development of Excessive Oxidative Stress and Cell Damage. Oxid. Med. Cell. Longev. 2019, 2019, 3218275. [Google Scholar] [CrossRef] [PubMed]
- Mirtschink, P.; Krek, W. Hypoxia-driven glycolytic and fructolytic metabolic programs: Pivotal to hypertrophic heart disease. Biochim. Biophys. Acta 2016, 1863, 1822–1828. [Google Scholar] [CrossRef]
- Mirtschink, P.; Krishnan, J.; Grimm, F.; Sarre, A.; Horl, M.; Kayikci, M.; Fankhauser, N.; Christinat, Y.; Cortijo, C.; Feehan, O.; et al. HIF-driven SF3B1 induces KHK-C to enforce fructolysis and heart disease. Nature 2015, 522, 444–449. [Google Scholar] [CrossRef]
- García-Arroyo, F.E.; Gonzaga-Sánchez, G.; Tapia, E.; Muñoz-Jiménez, I.; Manterola-Romero, L.; Osorio-Alonso, H.; Arellano-Buendía, A.S.; Pedraza-Chaverri, J.; Roncal-Jiménez, C.A.; Lanaspa, M.A.; et al. Osthol Ameliorates Kidney Damage and Metabolic Syndrome Induced by a High-Fat/High-Sugar Diet. Int. J. Mol. Sci. 2021, 22, 2431. [Google Scholar] [CrossRef]
- Hidalgo, I.; Nájera, N.; Meaney, E.; Pérez-Durán, J.; Valdespino-Vazquez, Y.; Villarreal, F.; Ceballos, G. Effects of (-)-epicatechin on the time course of the expression of perilipins in a diet-induced model of nonalcoholic steatohepatitis. J. Nutr. Biochem. 2020, 77, 108296. [Google Scholar] [CrossRef]
- Silva-Palacios, A.; Ostolga-Chavarria, M.; Buelna-Chontal, M.; Garibay, C.; Hernandez-Resendiz, S.; Roldan, F.J.; Flores, P.L.; Luna-Lopez, A.; Konigsberg, M.; Zazueta, C. 3-NP-induced Huntington’s-like disease impairs Nrf2 activation without loss of cardiac function in aged rats. Exp. Gerontol. 2017, 96, 89–98. [Google Scholar] [CrossRef]
- Stein, A.B.; Tiwari, S.; Thomas, P.; Hunt, G.; Levent, C.; Stoddard, M.F.; Tang, X.L.; Bolli, R.; Dawn, B. Effects of anesthesia on echocardiographic assessment of left ventricular structure and function in rats. Basic Res. Cardiol. 2007, 102, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Halhoul, M.N.; Kleinberg, I. Differential determination of glucose and fructose, and glucose- and fructose-yielding substances with anthrone. Anal. Biochem. 1972, 50, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, P.; Gersch, M.S.; Mu, W.; Scherer, P.M.; Kim, K.M.; Gesualdo, L.; Henderson, G.N.; Johnson, R.J.; Sautin, Y.Y. Ketohexokinase-dependent metabolism of fructose induces proinflammatory mediators in proximal tubular cells. J. Am. Soc. Nephrol. 2009, 20, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Kirby, E.D.; Kuwahara, A.A.; Messer, R.L.; Wyss-Coray, T. Adult hippocampal neural stem and progenitor cells regulate the neurogenic niche by secreting VEGF. Proc. Natl. Acad. Sci. USA 2015, 112, 4128–4133. [Google Scholar] [CrossRef] [PubMed]
- Molina-Jijon, E.; Tapia, E.; Zazueta, C.; El Hafidi, M.; Zatarain-Barron, Z.L.; Hernandez-Pando, R.; Medina-Campos, O.N.; Zarco-Marquez, G.; Torres, I.; Pedraza-Chaverri, J. Curcumin prevents Cr(VI)-induced renal oxidant damage by a mitochondrial pathway. Free Radic. Biol. Med. 2011, 51, 1543–1557. [Google Scholar] [CrossRef]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Daniels, L.J.; Annandale, M.; Koutsifeli, P.; Li, X.; Bussey, C.T.; van Hout, I.; Bunton, R.W.; Davis, P.J.; Coffey, S.; Katare, R.; et al. Elevated myocardial fructose and sorbitol levels are associated with diastolic dysfunction in diabetic patients, and cardiomyocyte lipid inclusions in vitro. Nutr. Diabetes 2021, 11, 8. [Google Scholar] [CrossRef]
- Daiber, A.; Hahad, O.; Andreadou, I.; Steven, S.; Daub, S.; Munzel, T. Redox-related biomarkers in human cardiovascular disease-Classical footprints and beyond. Redox Biol. 2021, 42, 101875. [Google Scholar] [CrossRef]
- Vileigas, D.F.; de Souza, S.L.B.; Correa, C.R.; Silva, C.; de Campos, D.H.S.; Padovani, C.R.; Cicogna, A.C. The effects of two types of Western diet on the induction of metabolic syndrome and cardiac remodeling in obese rats. J. Nutr. Biochem. 2021, 92, 108625. [Google Scholar] [CrossRef]
- Ishimoto, T.; Lanaspa, M.A.; Rivard, C.J.; Roncal-Jimenez, C.A.; Orlicky, D.J.; Cicerchi, C.; McMahan, R.H.; Abdelmalek, M.F.; Rosen, H.R.; Jackman, M.R.; et al. High fat and high sucrose (Western) diet induce steatohepatitis that is dependent on fructokinase. Hepatology 2013, 58, 1632–1643. [Google Scholar] [CrossRef]
- Janssen, J. The Impact of Westernization on the Insulin/IGF-I Signaling Pathway and the Metabolic Syndrome: It Is Time for Change. Int. J. Mol. Sci. 2023, 24, 4551. [Google Scholar] [CrossRef]
- Huang, W.C.; Liao, P.C.; Huang, C.H.; Hu, S.; Huang, S.C.; Wu, S.J. Osthole attenuates lipid accumulation, regulates the expression of inflammatory mediators, and increases antioxidants in FL83B cells. Biomed. Pharmacother. 2017, 91, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.Y.; Chou, T.C. Osthole Exerts Inhibitory Effects on Hypoxic Colon Cancer Cells via EIF2α Phosphorylation-mediated Apoptosis and Regulation of HIF-1α. Am. J. Chin. Med. 2022, 50, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Poulianiti, K.P.; Kaltsatou, A.; Mitrou, G.I.; Jamurtas, A.Z.; Koutedakis, Y.; Maridaki, M.; Stefanidis, I.; Sakkas, G.K.; Karatzaferi, C. Systemic Redox Imbalance in Chronic Kidney Disease: A Systematic Review. Oxid. Med. Cell. Longev. 2016, 2016, 8598253. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef]
- Lu, S.C. Regulation of glutathione synthesis. Mol. Asp. Med. 2009, 30, 42–59. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- Gutierrez, J.A.; Liu, W.; Perez, S.; Xing, G.; Sonnenberg, G.; Kou, K.; Blatnik, M.; Allen, R.; Weng, Y.; Vera, N.B.; et al. Pharmacologic inhibition of ketohexokinase prevents fructose-induced metabolic dysfunction. Mol. Metab. 2021, 48, 101196. [Google Scholar] [CrossRef]
- Sharma, N.; Okere, I.C.; Duda, M.K.; Chess, D.J.; O’Shea, K.M.; Stanley, W.C. Potential impact of carbohydrate and fat intake on pathological left ventricular hypertrophy. Cardiovasc. Res. 2007, 73, 257–268. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter/ Group | C | MS | MS+OT |
|---|---|---|---|
| IVS (mm) | 0.18 ± 0.019 | 0.205 ± 0.026 | 0.19 ± 0.012 |
| LVEDd (mm) | 4.96 ± 0.44 | 4.88 ± 0.39 | 4.62 ± 0.57 |
| LVEDs (mm) | 2.33 ± 0.53 | 2.06 ± 0.37 | 2.02 ± 0.43 |
| LVPW (mm) | 0.17 ± 0.019 | 0.19 ± 0.019 | 0.18 ± 0.016 |
| EF (%) | 81.4 ± 5.62 | 84.4 ± 6.1 | 83.3 ± 6.5 |
| FS (%) | 53.4 ± 7.36 | 57.7 ± 7.3 | 56.3 ± 8.5 |
| HR (bpm) | 424.5 ± 34.72 | 378.9 ± 51.4 | 412.5 ± 32.1 |
| Body weight (g) | 292.5 ± 5.82 | 336.4 ± 14.30 | 314 ± 15.9 |
| Heart weight (HW, g) | 1.05 ± 0.07 | 1.37 ± 0.05 a | 1.26 ± 0.03 b |
| Lung weight (LW, g) | 1.90 ± 0.13 | 3.83 ± 0.39 a | 2.91 ± 0.20 b |
| Tibial length (TL, cm) | 3.92 ± 0.14 | 4.20 ± 0.20 | 3.84 ± 0.83 |
| HW/TL (g/cm) | 0.27 ± 0.02 | 0.33 ± 0.02 | 0.35 ± 0.11 |
| LW/TL (g/cm) | 0.49 ± 0.04 | 0.91 ± 0.10 a | 0.82 ± 0.32 a |
| HW/BW (g/kg) | 3.52 ± 0.20 | 4.56 ± 0.24 a | 4.38 ± 0.29 a |
| LW/BW (g/kg) | 6.38 ± 0.55 | 12.72 ± 1.38 a | 10.16 ± 1.26 a,b |
| Functional parameters | (n = 7) | (n = 8) | (n = 7) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Arroyo, F.E.; Gonzaga-Sánchez, G.; Silva-Palacios, A.; Roldán, F.J.; Loredo-Mendoza, M.L.; Alvarez-Alvarez, Y.Q.; de los Santos Coyotl, J.A.; Vélez Orozco, K.A.; Tapia, E.; Osorio-Alonso, H.; et al. Osthole Prevents Heart Damage Induced by Diet-Induced Metabolic Syndrome: Role of Fructokinase (KHK). Antioxidants 2023, 12, 1023. https://doi.org/10.3390/antiox12051023
García-Arroyo FE, Gonzaga-Sánchez G, Silva-Palacios A, Roldán FJ, Loredo-Mendoza ML, Alvarez-Alvarez YQ, de los Santos Coyotl JA, Vélez Orozco KA, Tapia E, Osorio-Alonso H, et al. Osthole Prevents Heart Damage Induced by Diet-Induced Metabolic Syndrome: Role of Fructokinase (KHK). Antioxidants. 2023; 12(5):1023. https://doi.org/10.3390/antiox12051023
Chicago/Turabian StyleGarcía-Arroyo, Fernando E., Guillermo Gonzaga-Sánchez, Alejandro Silva-Palacios, Francisco Javier Roldán, María L. Loredo-Mendoza, Yamnia Quetzal Alvarez-Alvarez, Jesus A. de los Santos Coyotl, Kevin A. Vélez Orozco, Edilia Tapia, Horacio Osorio-Alonso, and et al. 2023. "Osthole Prevents Heart Damage Induced by Diet-Induced Metabolic Syndrome: Role of Fructokinase (KHK)" Antioxidants 12, no. 5: 1023. https://doi.org/10.3390/antiox12051023