
Regulation of Redox Profile and Genomic Instability by Physical Exercise Contributes to Neuroprotection in Mice with Experimental Glioblastoma
 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Bioethical Procedures
2.2. Training Protocol
2.3. Experimental Glioblastoma Protocol
2.4. Body Mass, Water, and Food Control
2.5. Rotarod Test
2.6. Open-Field Test
2.7. Euthanasia and Sample Preparation
2.8. Redox Parameter Assays
2.9. Comet Test
2.10. Micronucleus Test
2.11. Data Analysis
3. Results
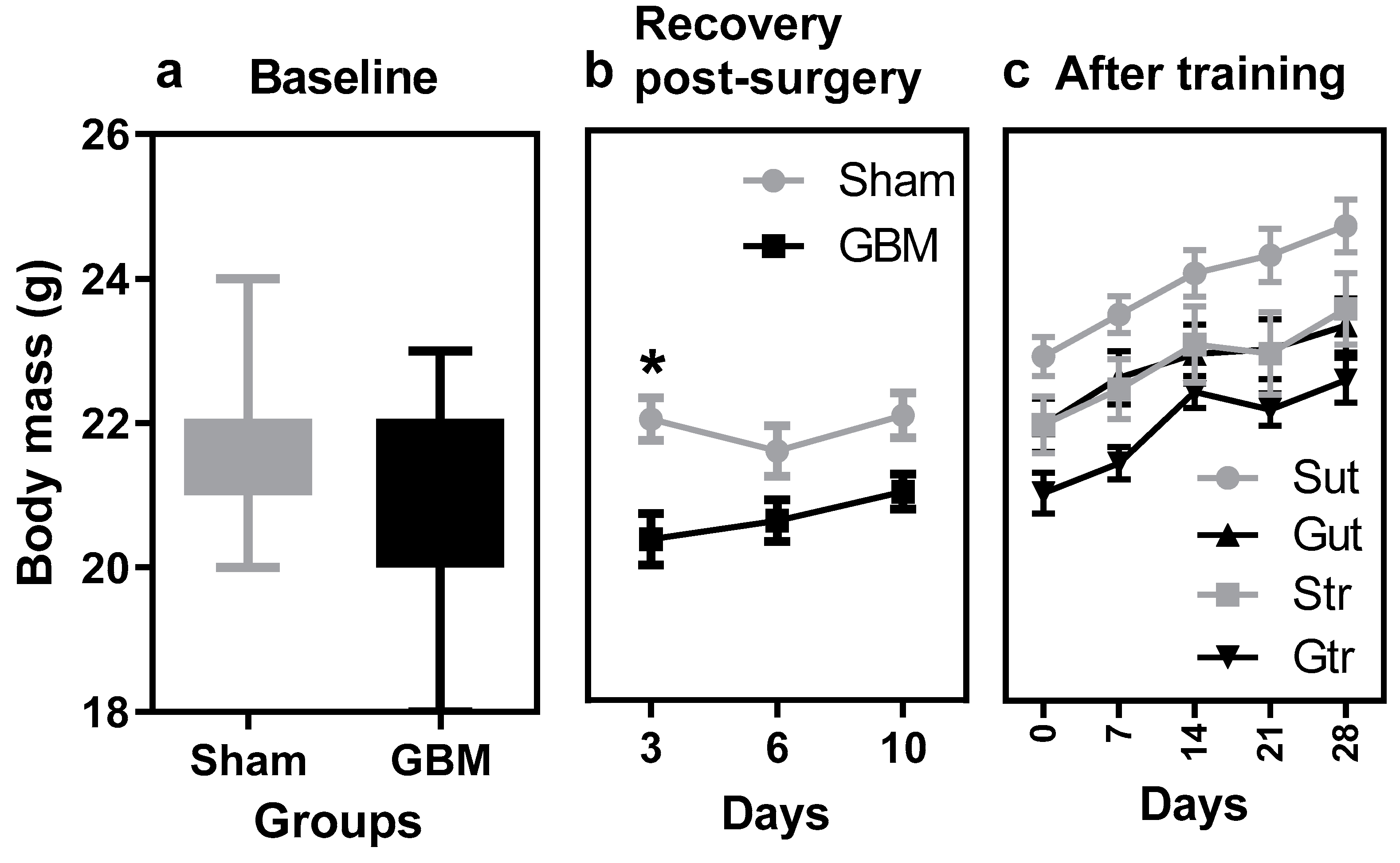
3.1. Body Mass Control
3.2. Control of Water and Food Consumption
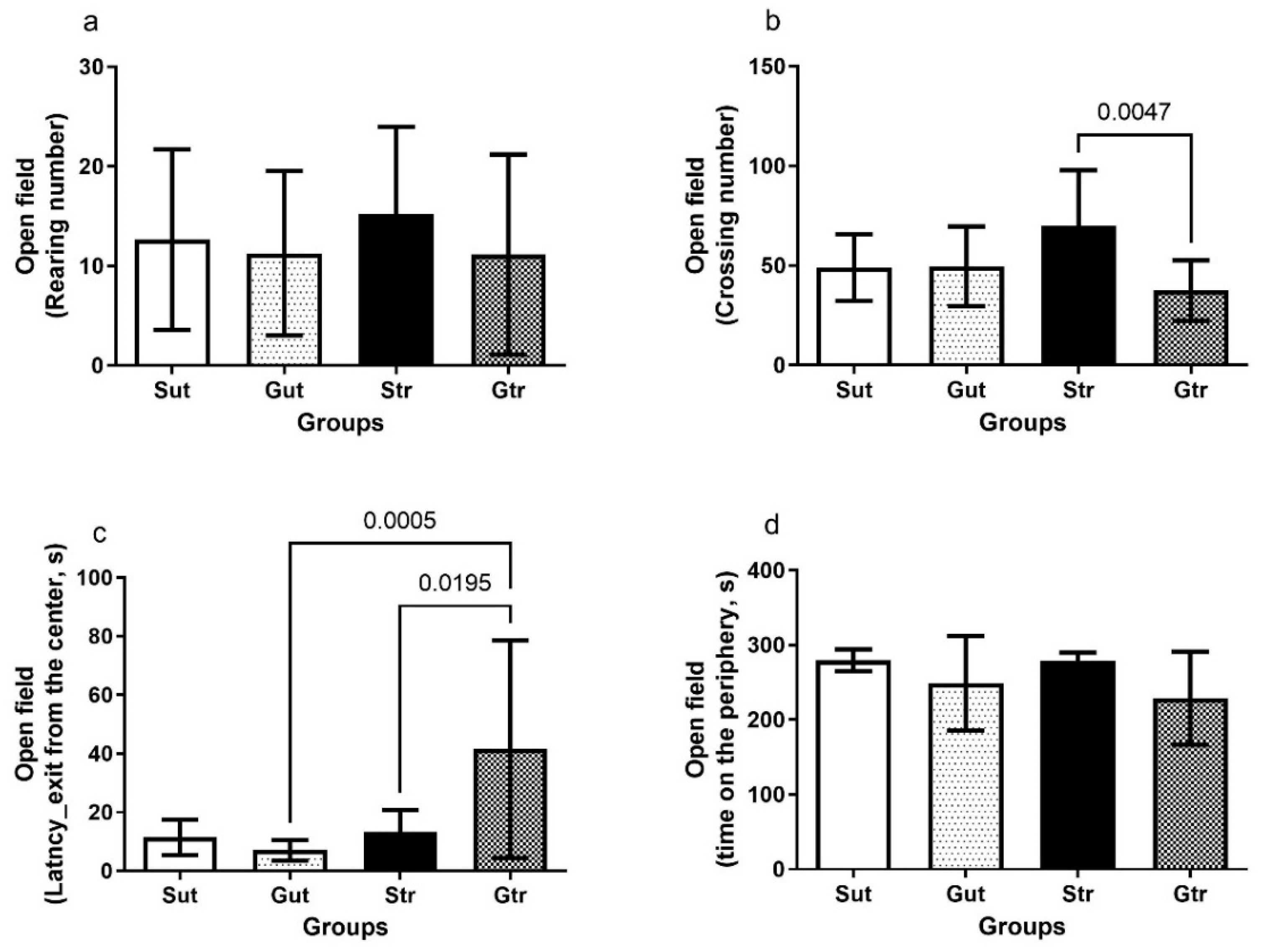
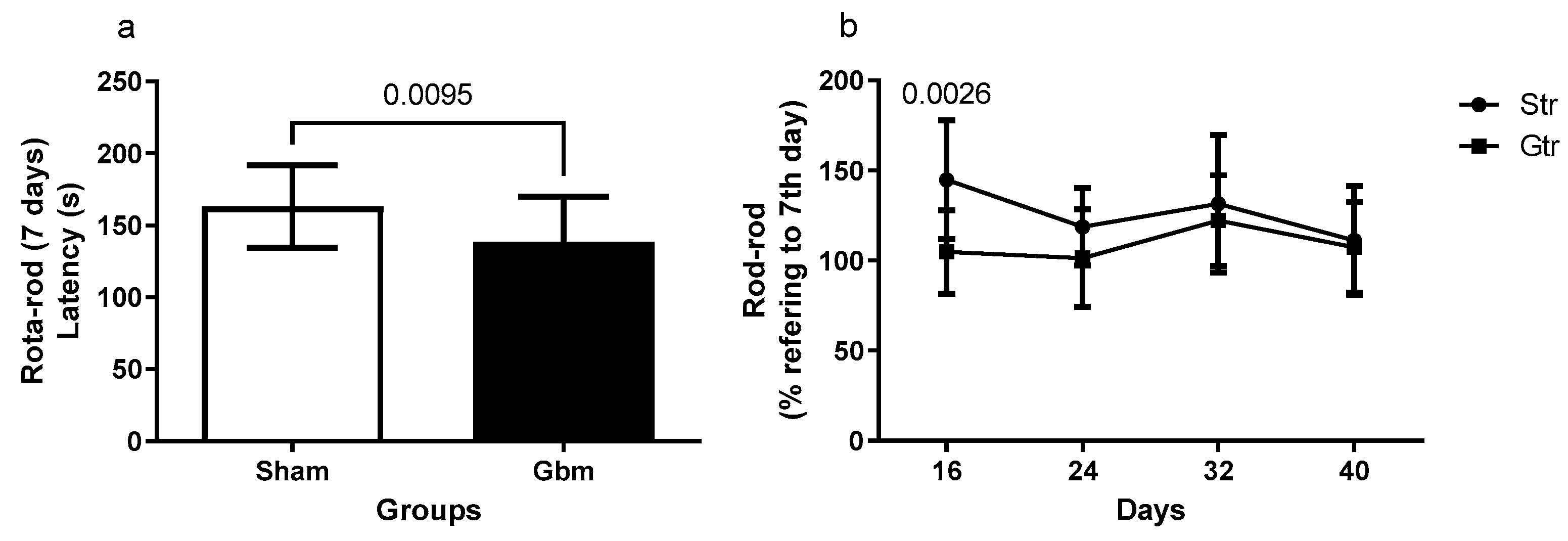
3.3. Behavioral Parameters
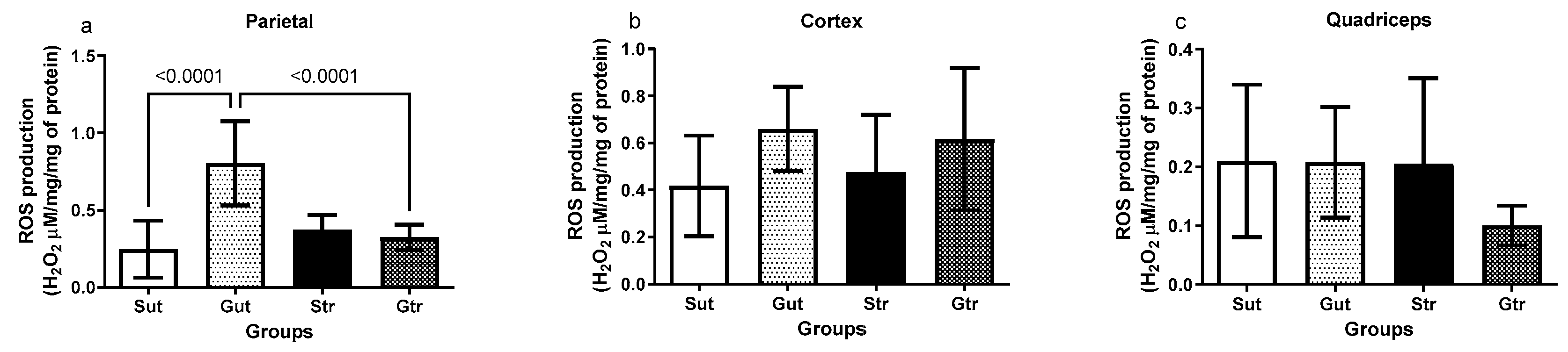
3.4. Production of Reactive Oxygen Species
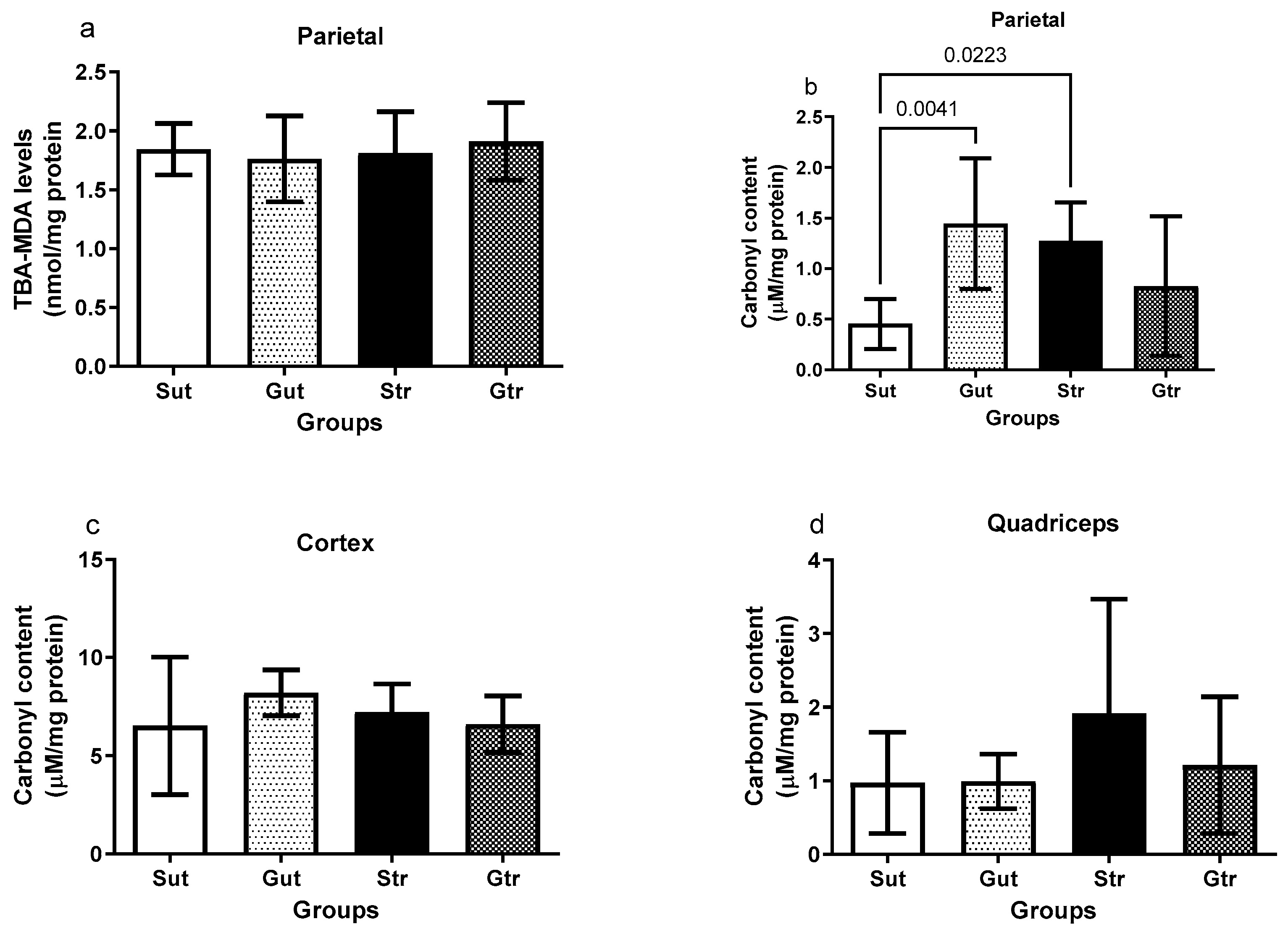
3.5. Oxidative Damage
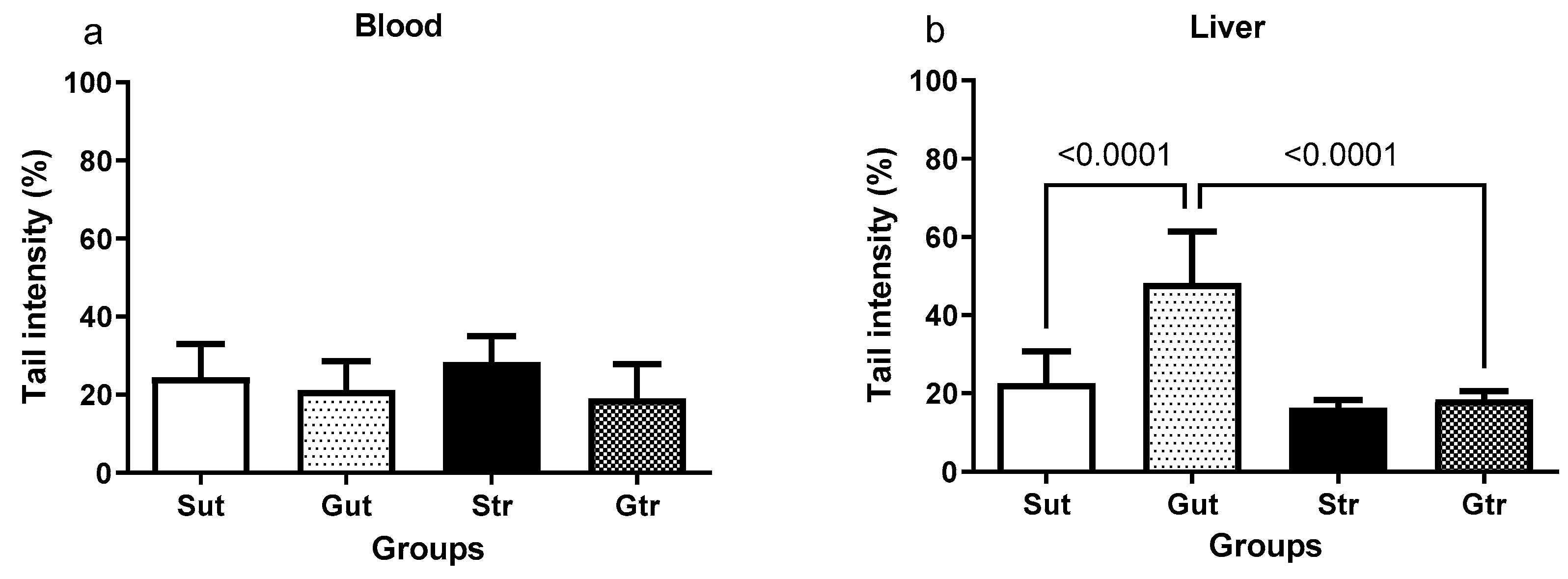
3.6. DNA Damage
3.7. Micronucleus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kristensen, B.W.; Priesterbach-Ackley, L.P.; Petersen, J.K.; Wesseling, P. Molecular Pathology of Tumors of the Central Nervous System. Ann. Oncol. 2019, 30, 1265–1278. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Stetson, L.; Virk, S.M.; Barnholtz-Sloan, J.S. Epidemiology of Gliomas. Cancer Treat. Res. 2015, 163, 1–14. [Google Scholar] [CrossRef]
- Wirsching, H.G.; Galanis, E.; Weller, M. Glioblastoma. Handb. Clin. Neurol. 2016, 134, 381–397. [Google Scholar] [CrossRef]
- Zhou, C.; Zhao, H.; Yang, F.; Huangfu, L.; Dong, C.; Wang, S.; Zhang, J. Clinical and Genetic Features of Brainstem Glioma in Adults: A Report of 50 Cases in a Single Center. J. Clin. Neurol. 2021, 17, 220–228. [Google Scholar] [CrossRef]
- Marques-Aleixo, I.; Beleza, J.; Sampaio, A.; Stevanović, J.; Coxito, P.; Gonçalves, I.; Ascensão, A.; Magalhães, J. Preventive and Therapeutic Potential of Physical Exercise in Neurodegenerative Diseases. Antioxid. Redox Signal. 2021, 34, 674–693. [Google Scholar] [CrossRef]
- Harris, G.; Jayamanne, D.; Wheeler, H.; Gzell, C.; Kastelan, M.; Schembri, G.; Brazier, D.; Cook, R.; Parkinson, J.; Khasraw, M.; et al. Survival Outcomes of Elderly Patients with Glioblastoma Multiforme in Their 75th Year or Older Treated with Adjuvant Therapy. Int. J. Radiat. Oncol. Biol. Phys. 2017, 98, 802–810. [Google Scholar] [CrossRef]
- Akmal, M.; Hasnain, N.; Rehan, A.; Iqbal, U.; Hashmi, S.; Fatima, K.; Farooq, M.Z.; Khosa, F.; Siddiqi, J.; Khan, M.K. Glioblastome Multiforme: A Bibliometric Analysis. World Neurosurg. 2020, 136, 270–282. [Google Scholar] [CrossRef]
- Lemke, D.; Pledl, H.W.; Zorn, M.; Jugold, M.; Green, E.; Blaes, J.; Löw, S.; Hertenstein, A.; Ott, M.; Sahm, F.; et al. Slowing down Glioblastoma Progression in Mice by Running or the Anti-Malarial Drug Dihydroartemisinin? Induction of Oxidative Stress in Murine Glioblastoma Therapy. Oncotarget 2016, 7, 56713–56725. [Google Scholar] [CrossRef]
- du Sert, N.P.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting Animal Research: Explanation and Elaboration for the Arrive Guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar]
- Souza, P.S.; Gonçalves, E.D.; Pedroso, G.S.; Farias, H.R.; Junqueira, S.C.; Marcon, R.; Tuon, T.; Cola, M.; Silveira, P.C.L.; Santos, A.R.; et al. Physical Exercise Attenuates Experimental Autoimmune Encephalomyelitis by Inhibiting Peripheral Immune Response and Blood-Brain Barrier Disruption. Mol. Neurobiol. 2017, 54, 4723–4737. [Google Scholar] [CrossRef]
- Lorico, A.; Mercapide, J.; Soloduschko, V.; Alexeyev, M.; Fodstad, O.; Rappa, G. Primary Neural Stem/Progenitor Cells Expressing Endostatin or Cytochrome P450 for Gene Therapy of Glioblastoma. Cancer Gene Ther. 2008, 15, 605–615. [Google Scholar] [CrossRef]
- Godoy, M.C.M.; Fighera, M.R.; Souza, F.R.; Flores, A.E.; Rubin, M.A.; Oliveira, M.R.; Zanatta, N.; Martins, M.A.P.; Bonacorso, H.G.; Mello, C.F. A2-Adrenoceptors and 5-HT Receptors Mediate the Antinociceptive Effect of New Pyrazolines, but Not of Dipyrone. Eur. J. Pharmacol. 2004, 496, 93–97. [Google Scholar] [CrossRef]
- Archer, J. Tests for Emotionality in Rats and Mice: A Review. Anim. Behav. 1973, 21, 205–235. [Google Scholar] [CrossRef]
- Wei, J.; Xie, C.-Z.; Yuan, Q. Improvements of Motor and Cognitive Functions in the Rats with Permanent Middle Cerebral Artery Occlusion Treated with Acupuncture and Rota-Rod Training. Zhongguo Zhen Jiu 2019, 39, 748–754. [Google Scholar]
- Colombo, G.; Clerici, M.; Garavaglia, M.E.; Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. A Step-by-Step Protocol for Assaying Protein Carbonylation in Biological Samples. J. Chromatogr. B 2016, 1019, 178–190. [Google Scholar] [CrossRef]
- Ernst, O.; Zor, T. Linearization of the Bradford Protein Assay. J. Vis. Exp. 2010, 38, 1918. [Google Scholar] [CrossRef]
- Valencia, L.C.; García, A.; Ramírez-Pinilla, M.P.; Fuentes, J.L. Estimates of DNA Damage by the Comet Assay in the Direct-Developing Frog Eleutherodactylus johnstonei (Anura, Eleutherodactylidae). Genet. Mol. Biol. 2011, 34, 681–688. [Google Scholar] [CrossRef]
- Krishna, G.; Hayashi, M. In Vivo Rodent Micronucleus Assay: Protocol, Conduct and Data Interpretation. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2000, 455, 155–166. [Google Scholar] [CrossRef]
- Karremann, M.; Gielen, G.H.; Hoffmann, M.; Wiese, M.; Colditz, N.; Warmuth-Metz, M.; Bison, B.; Claviez, A.; Van Vuurden, D.G.; Von Bueren, A.O.; et al. Diffuse High-Grade Gliomas with H3 K27M Mutations Carry a Dismal Prognosis Independent of Tumor Location. Neuro Oncol. 2018, 20, 123–131. [Google Scholar] [CrossRef]
- Ladomersky, E.; Genet, M.; Zhai, L.; Gritsina, G.; Lauing, K.L.; Lulla, R.R.; Fangusaro, J.; Lenzen, A.; Kumthekar, P.; Raizer, J.J.; et al. Improving Vaccine Efficacy against Malignant Glioma. Oncoimmunology 2016, 5, e1196311. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to Measure Locomotor and Anxiety-like Behavior in Mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef]
- Shoji, H.; Miyakawa, T. Age-Related Behavioral Changes from Young to Old Age in Male Mice of a C57BL/6J Strain Maintained under a Genetic Stability Program. Neuropsychopharmacol. Rep. 2019, 39, 100–118. [Google Scholar] [CrossRef]
- Tantillo, E.; Colistra, A.; Baroncelli, L.; Costa, M.; Caleo, M.; Vannini, E. Voluntary Physical Exercise Reduces Motor Dysfunction and Hampers Tumor Cell Proliferation in a Mouse Model of Glioma. Int. J. Environ. Res. Public. Health 2020, 17, 5667. [Google Scholar] [CrossRef]
- Assi, M.; Dufresne, S.; Rébillard, A. Exercise Shapes Redox Signaling in Cancer. Redox Biol. 2020, 35, 101439. [Google Scholar] [CrossRef]
- Burtscher, J.; Millet, G.P.; Place, N.; Kayser, B.; Zanou, N. The Muscle-Brain Axis and Neurodegenerative Diseases: The Key Role of Mitochondria in Exercise-Induced Neuroprotection. Int. J. Mol. Sci. 2021, 22, 6479. [Google Scholar] [CrossRef]
- Prasad, S.; Gupta, S.C.; Tyagi, A.K. Reactive Oxygen Species (ROS) and Cancer: Role of Antioxidative Nutraceuticals. Cancer Lett. 2017, 387, 95–105. [Google Scholar] [CrossRef]
- Bailey, D.M. Oxygen, Evolution and Redox Signalling in the Human Brain; Quantum in the Quotidian. J. Physiol. 2019, 597, 15–28. [Google Scholar] [CrossRef]
- Jelinek, M.; Jurajda, M.; Duris, K. Oxidative Stress in the Brain: Basic Concepts and Treatment Strategies in Stroke. Antioxidants 2021, 10, 1886. [Google Scholar] [CrossRef]
- Adler, V.; Yin, Z.; Tew, K.D.; Ronai, Z. Role of Redox Potential and Reactive Oxygen Species in Stress Signaling. Oncogene 1999, 18, 6104–6111. [Google Scholar] [CrossRef]
- Salazar-Ramiro, A.; Ramírez-Ortega, D.; de La Cruz, V.P.; Hérnandez-Pedro, N.Y.; González-Esquivel, D.F.; Sotelo, J.; Pineda, B. Role of Redox Status in Development of Glioblastoma. Front. Immunol. 2016, 7, 156. [Google Scholar] [CrossRef]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Eustress and Distress in Redox Homeostasis; Elsevier, Inc.: Amsterdam, The Netherlands, 2019; ISBN 9780128131466. [Google Scholar]
- Souza, J.; da Silva, R.A.; da Luz Scheffer, D.; Penteado, R.; Solano, A.; Barros, L.; Budde, H.; Trostchansky, A.; Latini, A. Physical-Exercise-Induced Antioxidant Effects on the Brain and Skeletal Muscle. Antioxidants 2022, 11, 826. [Google Scholar] [CrossRef]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A Mutual Interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef]
- Reczek, C.R.; Chandel, N.S. ROS-Dependent Signal Transduction. Curr. Opin. Cell. Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef]
- Sies, H. Hydrogen Peroxide as a Central Redox Signaling Molecule in Physiological Oxidative Stress: Oxidative Eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Meitzler, J.L.; Konaté, M.M.; Doroshow, J.H. Hydrogen Peroxide-Producing NADPH Oxidases and the Promotion of Migratory Phenotypes in Cancer. Arch. Biochem. Biophys. 2019, 675, 108076. [Google Scholar] [CrossRef]
- Takashi, Y.; Tomita, K.; Kuwahara, Y.; Roudkenar, M.H.; Roushandeh, A.M.; Igarashi, K.; Nagasawa, T.; Nishitani, Y.; Sato, T. Mitochondrial Dysfunction Promotes Aquaporin Expression That Controls Hydrogen Peroxide Permeability and Ferroptosis. Free Radic. Biol. Med. 2020, 161, 60–70. [Google Scholar] [CrossRef]
- Schoenfeld, J.D.; Sibenaller, Z.A.; Mapuskar, K.A.; Wagner, B.A.; Cramer-Morales, K.L.; Furqan, M.; Sandhu, S.; Carlisle, T.L.; Smith, M.C.; Abu Hejleh, T.; et al. O2− and H2O2-Mediated Disruption of Fe Metabolism Causes the Differential Susceptibility of NSCLC and GBM Cancer Cells to Pharmacological Ascorbate. Cancer Cell 2017, 31, 487–500.e8. [Google Scholar] [CrossRef]
- Pinho, R.A.; Aguiar, A.S.; Radák, Z. Effects of Resistance Exercise on Cerebral Redox Regulation and Cognition: An Interplay between Muscle and Brain. Antioxidants 2019, 8, 529. [Google Scholar] [CrossRef]
- Cabiscol, E.; Tamarit, J.; Ros, J. Protein Carbonylation: Proteomics, Specificity and Relevance to Aging. Mass. Spectrom. Rev. 2014, 33, 21–48. [Google Scholar] [CrossRef]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein Carbonylation as a Major Hallmark of Oxidative Damage: Update of Analytical Strategies. Mass. Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Kryndushkin, D.; Wu, W.W.; Venna, R.; Norcross, M.A.; Shen, R.F.; Rao, V.A. Complex Nature of Protein Carbonylation Specificity After Metal-Catalyzed Oxidation. Pharm. Res. 2017, 34, 765–779. [Google Scholar] [CrossRef] [PubMed]
- Hardiany, N.S.; Mulyawan, W.; Wanandi, S.I. Correlation between Oxidative Stress and Tumor Grade in Glioma Cells from Patients in Jakarta. Med. J. Indones. 2012, 21, 122–127. [Google Scholar] [CrossRef]
- Liu, W.; Kuang, H.; Xia, Y.; Pope, Z.C.; Wang, Z.; Tang, C.; Yin, D. Regular Aerobic Exercise-Ameliorated Troponin I Carbonylation to Mitigate Aged Rat Soleus Muscle Functional Recession. Exp. Physiol. 2019, 104, 715–728. [Google Scholar] [CrossRef]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.H.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.M.; Gallia, G.L.; et al. An Integrated Genomic Analysis of Human Glioblastoma Multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef]
- Møller, P. Genotoxicity of Environmental Agents Assessed by the Alkaline Comet Assay. Basic. Clin. Pharmacol. Toxicol. 2005, 96, 1–42. [Google Scholar] [CrossRef]
- Ali, M.M.; Sahar, T.; Firyal, S.; Ijaz, M.; Majeed, K.A.; Awan, F.; Adil, M.; Akbar, H.; Rashid, M.I.; Ciğerci, İ.H. Assessment of Cytotoxic, Genotoxic, and Oxidative Stress of Dibutyl Phthalate on Cultured Bovine Peripheral Lymphocytes. Oxid. Med. Cell. Longev. 2022, 2022, 9961513. [Google Scholar] [CrossRef]
- Saleh, E.M.; Hamdy, G.M.; Hassan, R.E. Neuroprotective Effect of Sodium Alginate against Chromium-Induced Brain Damage in Rats. PLoS ONE 2022, 17, e0266898. [Google Scholar] [CrossRef]
- Netto, A.O.; Gelaleti, R.B.; Corvino, S.B.; Serrano, R.G.; Hernández, S.C.; Volpato, G.T.; Rudge, M.V.C.; Braz, M.G.; Damasceno, D.C. Small-for-Pregnancy-Age Rats Submitted to Exercise: DNA Damage in Mothers and Newborns, Measured by the Comet Assay. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 835, 11–15. [Google Scholar] [CrossRef]
- Tryfidou, D.V.; McClean, C.; Nikolaidis, M.G.; Davison, G.W. DNA Damage Following Acute Aerobic Exercise: A Systematic Review and Meta-Analysis. Sport. Med. 2020, 50, 103–127. [Google Scholar] [CrossRef]
- Sommer, S.; Buraczewska, I.; Kruszewski, M. Micronucleus Assay: The State of Art, and Future Directions. Int. J. Mol. Sci. 2020, 21, 1534. [Google Scholar] [CrossRef]
- Decordier, I.; Kirsch-Volders, M. The in Vitro Micronucleus Test: From Past to Future. Mutat. Res. 2006, 607, 2–4. [Google Scholar] [CrossRef]
- Vijitkul, P.; Kongsema, M.; Toommakorn, T.; Bullangpoti, V. Investigation of Genotoxicity, Mutagenicity, and Cytotoxicity in Erythrocytes of Nile Tilapia (Oreochromis niloticus) after Fluoxetine Exposure. Toxicol. Rep. 2022, 9, 588–596. [Google Scholar] [CrossRef]
- Damiani, A.P.; Garcez, M.L.; Letieli de Abreu, L.; Tavares, T.H.; Rodrigues Boeck, C.; Moraes de Andrade, V. A Reduction in DNA Damage in Neural Tissue and Peripheral Blood of Old Mice Treated with Caffeine. J. Toxicol. Environ. Health A 2017, 80, 621–629. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Average Water Consumption (Mean ± Standard Deviation) | % Change * | Average Food Intake (Mean ± Standard Deviation) | % Change * | ||||
|---|---|---|---|---|---|---|---|---|
| Post-Surgery | Day 7 | Day 48 | Post-Surgery | Day 7 | Day 48 | |||
| Sut | 2.8 ± 0.9 | 4.8 ± 0.9 | 4.7 ± 1.1 | −1.3 | 2.7 ± 0.2 | 4.8 ± 0.8 | 4.7 ± 1.1 | −1.3 |
| Gut | 1.1 ± 0.3 | 3.1 ± 1.1 | 3.5 ± 0.1 | 16.5 | 1.1 ± 0.3 | 3.0 ± 0.9 | 3.5 ± 0.2 | 16.6 |
| Str | 1.6 ± 0.9 | 5.1 ± 0.5 | 4.3 ± 1.3 | −14.7 | 1.6 ± 0.9 | 5.0 ± 0.5 | 4.3 ± 1.1 | −14.3 |
| Gtr | 1.0 ± 0.4 | 3.9 ± 0.1 | 3.3 ± 0.0 | −13.7 | 0.9 ± 0.1 | 3.7 ± 0.1 | 3.4 ± 0.1 | −8.1 |
| Groups | MN-PCE | MN-MCE | PCE/NCE |
|---|---|---|---|
| Sut | 2.71 ± 1.11 | 0.71 ± 1.11 | 0.56 ± 0.05 |
| Str | 3.25 ± 0.95 | 1.57 ± 1.81 | 0.50 ± 0.05 |
| Gut | 4.33 ± 1.03 * | 1.10 ± 1.37 | 0.52 ± 0.05 |
| Gtr | 6.57 ± 1.90 #& | 1.90 ± 1.85 | 0.56 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marqueze, L.F.B.; Costa, A.K.; Pedroso, G.S.; Vasconcellos, F.F.; Pilger, B.I.; Kindermann, S.; Andrade, V.M.; Alves, A.C.B.; Nery, T.; Silva, A.A., Jr.; et al. Regulation of Redox Profile and Genomic Instability by Physical Exercise Contributes to Neuroprotection in Mice with Experimental Glioblastoma. Antioxidants 2023, 12, 1343. https://doi.org/10.3390/antiox12071343
Marqueze LFB, Costa AK, Pedroso GS, Vasconcellos FF, Pilger BI, Kindermann S, Andrade VM, Alves ACB, Nery T, Silva AA Jr., et al. Regulation of Redox Profile and Genomic Instability by Physical Exercise Contributes to Neuroprotection in Mice with Experimental Glioblastoma. Antioxidants. 2023; 12(7):1343. https://doi.org/10.3390/antiox12071343
Chicago/Turabian StyleMarqueze, Luis F. B., Amanda K. Costa, Giulia S. Pedroso, Franciane F. Vasconcellos, Bruna I. Pilger, Schellen Kindermann, Vanessa M. Andrade, Ana C. B. Alves, Tatyana Nery, Aderbal A. Silva, Jr., and et al. 2023. "Regulation of Redox Profile and Genomic Instability by Physical Exercise Contributes to Neuroprotection in Mice with Experimental Glioblastoma" Antioxidants 12, no. 7: 1343. https://doi.org/10.3390/antiox12071343
APA StyleMarqueze, L. F. B., Costa, A. K., Pedroso, G. S., Vasconcellos, F. F., Pilger, B. I., Kindermann, S., Andrade, V. M., Alves, A. C. B., Nery, T., Silva, A. A., Jr., Carvalhal, S. R. S., Zazula, M. F., Naliwaiko, K., Fernandes, L. C., Radak, Z., & Pinho, R. A. (2023). Regulation of Redox Profile and Genomic Instability by Physical Exercise Contributes to Neuroprotection in Mice with Experimental Glioblastoma. Antioxidants, 12(7), 1343. https://doi.org/10.3390/antiox12071343