
In Vitro Bioactivities of Commonly Consumed Cereal, Vegetable, and Legume Seeds as Related to Their Bioactive Components: An Untargeted Metabolomics Approach Using UHPLC–QTOF-MS2

 ,
,
Abstract
:1. Introduction
2. Materials and Methodology
2.1. Plant Materials Collection
2.2. Chemicals
2.3. Preparation of Ethanol Extract
2.4. Total Phenolic, Flavonoids, and Saponin Contents
2.5. DPPH and ABTS Radical Inhibition Assays
2.6. Pancreatic Lipase Inhibition Assay
2.7. Alpha-Glucosidase Inhibitory Assay
2.8. Inhibition of AGEs Formation
2.9. UHPLC–QTOF-MS/MS2 Metabolite Identification
2.10. Statistical Analysis
3. Results
3.1. Total Phenolic (TPC), Flavonoid (TFC), Saponin (TSC) Contents
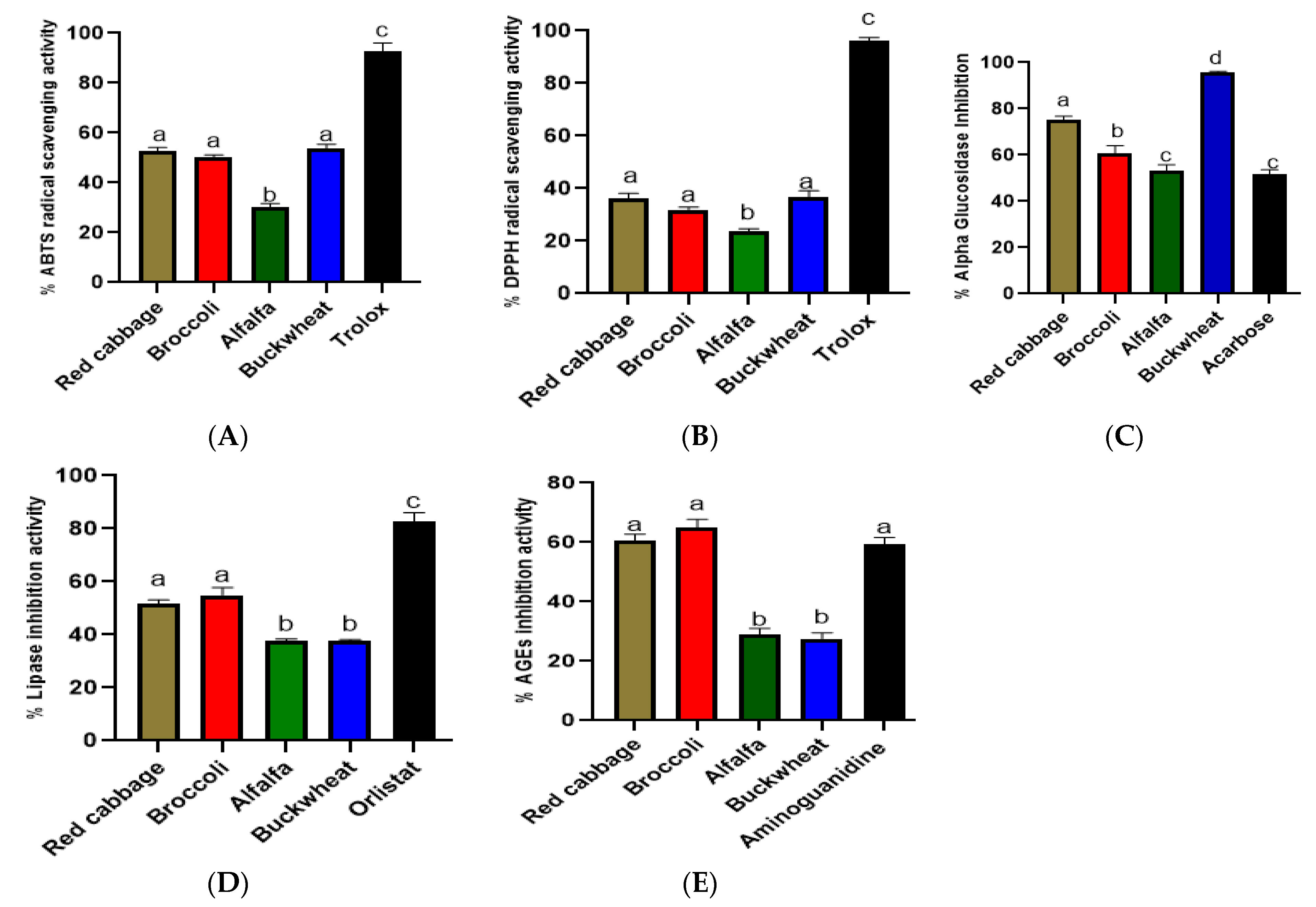
3.2. In Vitro Antioxidant, AGEs Formation Inhibition, Alpha-Glucosidase Inhibition, and Pancreatic Lipase Inhibition Activities
3.3. Comprehensive Profiling of Possible Bioactive Compounds in Alfalfa, Buckwheat, Broccoli, and Red Cabbage Extracts Using UHPLC–QTOF-MS/MS2
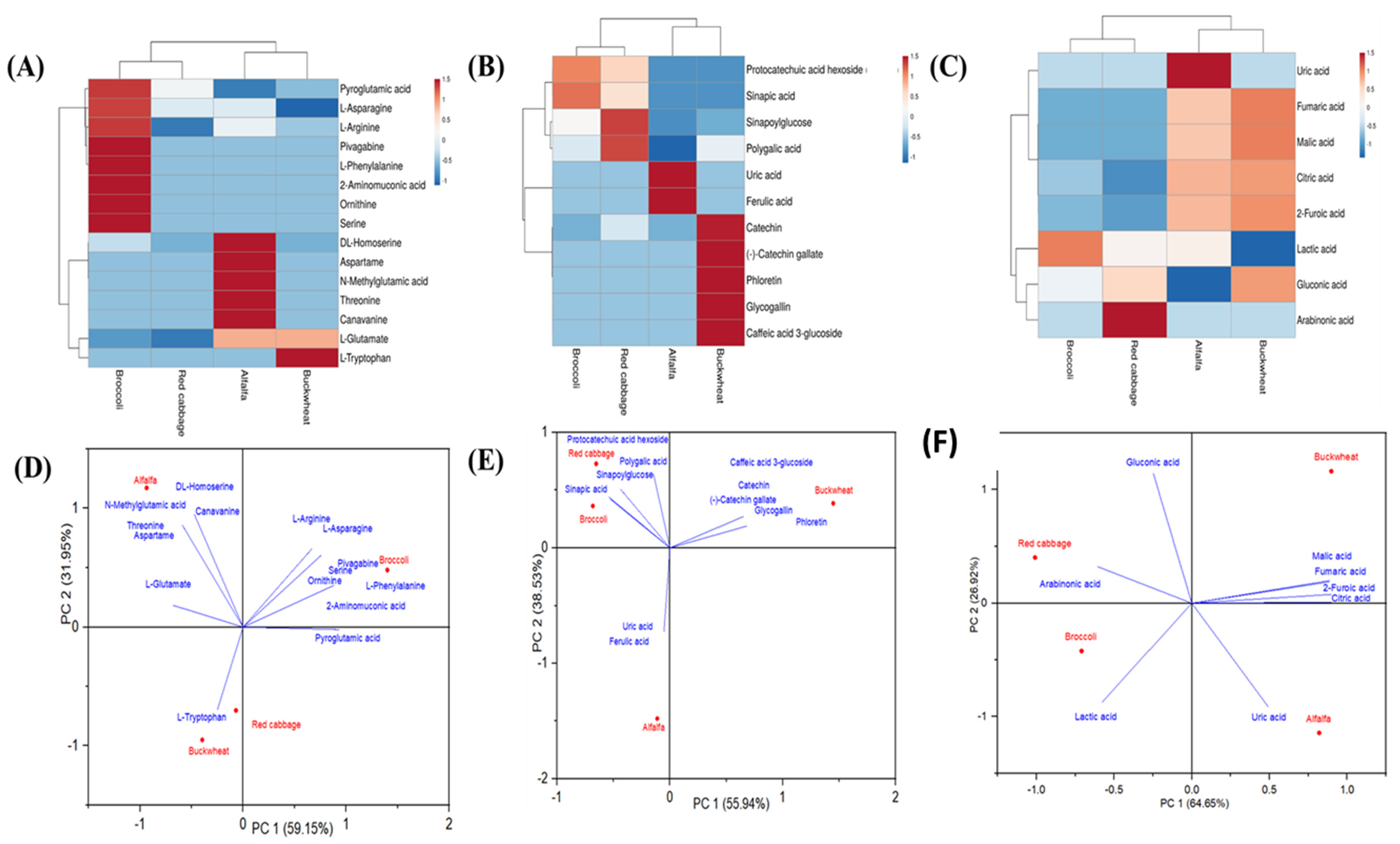
3.4. Heatmap Visualization and Principal Component Analysis of Tentatively Identified Metabolites
3.5. Exploring the Contribution of Amino Acids, Polyphenols, and Organic Acids to the Functional Properties of Red Cabbage, Broccoli, Alfalfa and Buckwheat
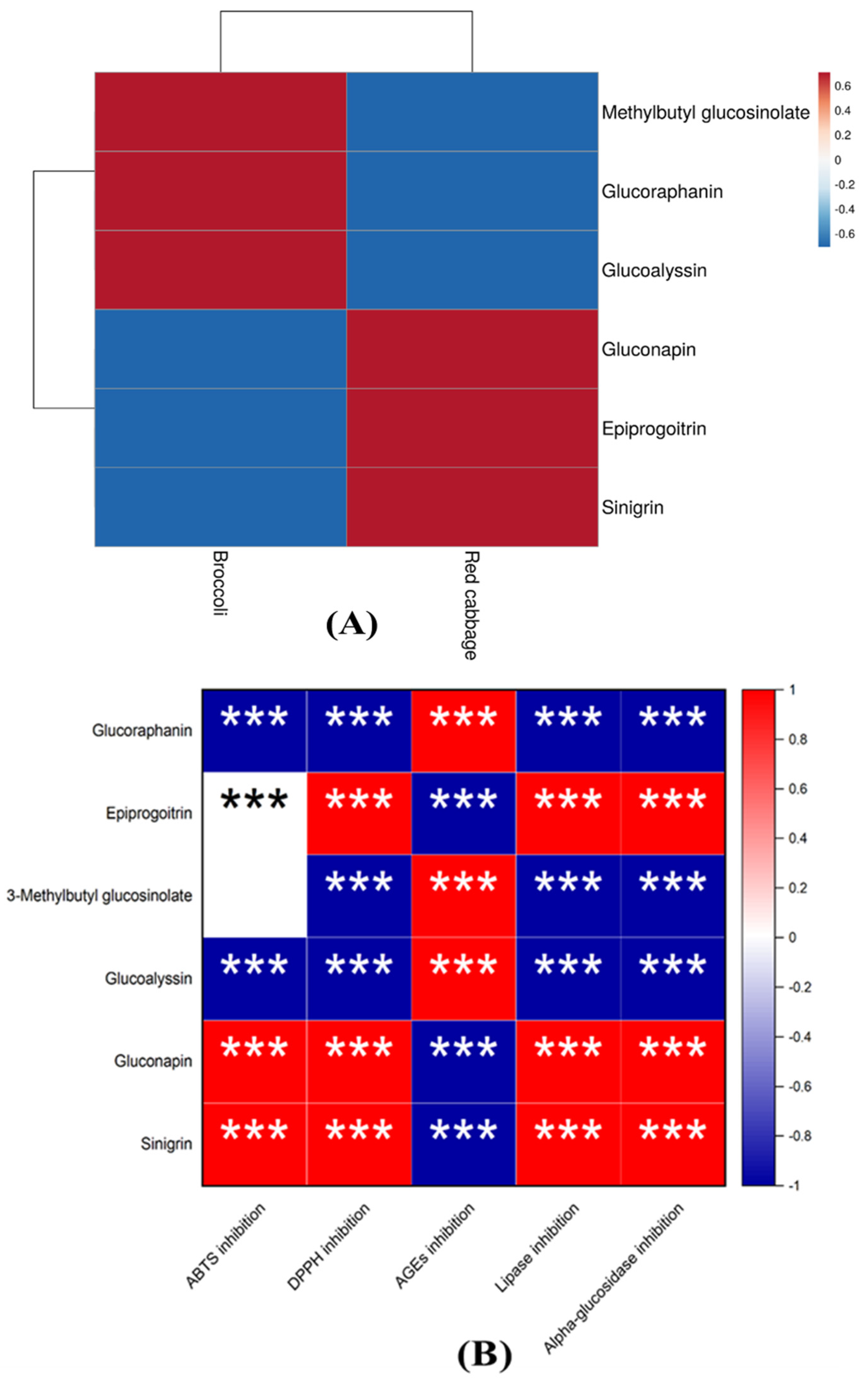
3.6. Correlation between Glucosinolates Identified in Red Cabbage and Broccoli Seeds and Functional Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABTS | 2, 2-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid |
| AGE | Advanced glycation end produs |
| CE | Catechin equivalent |
| FAE | Ferrulic acid equivalent |
| SSBE | Soysaponin B equivalents |
| DPPH | 2-diphenyl-1-picrylhydrazyl |
| OD | Optical density |
| TPC | Total phenolic content |
| TFC | Total flavonoid content |
| TSC | Total saponin content |
| 4-MU | 4-Methylumbelliferyl |
References
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef]
- Salehi, B.; Quispe, C.; Butnariu, M.; Sarac, I.; Marmouzi, I.; Kamle, M.; Tripathi, V.; Kumar, P.; Bouyahya, A.; Capanoglu, E. Phytotherapy and food applications from Brassica genus. Phytother. Res. 2021, 35, 3590–3609. [Google Scholar] [CrossRef]
- Hafidh, R.; Abdulamir, A.; Bakar, F.A.; Jalilian, F.; Jahanshiri, F.; Abas, F.; Sekawi, Z. Novel anticancer activity and anticancer mechanisms of Brassica oleracea L. var. capitata f. rubra. Eur. J. Integr. Med. 2013, 5, 450–464. [Google Scholar] [CrossRef]
- Mohammed, A.; Mohammed, H.A. Beneficial role of broccoli and its active ingredient, sulforaphane in the treatment of diabetes. Phytomedicine Plus 2023, 3, 100431. [Google Scholar] [CrossRef]
- Gimenez-Bastida, J.A.; Zielinski, H. Buckwheat as a functional food and its effects on health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef]
- Aloo, S.-O.; Ofosu, F.-K.; Oh, D.-H. Effect of Germination on Alfalfa and Buckwheat: Phytochemical Profiling by UHPLC-ESI-QTOF-MS/MS, Bioactive Compounds, and In-Vitro Studies of Their Diabetes and Obesity-Related Functions. Antioxidants 2021, 10, 1613. [Google Scholar] [CrossRef]
- Morishita, T.; Yamaguchi, H.; Degi, K. The contribution of polyphenols to antioxidative activity in common buckwheat and tartary buckwheat grain. Plant Prod. Sci. 2007, 10, 99–104. [Google Scholar] [CrossRef]
- Zhang, Z.-L.; Zhou, M.-L.; Tang, Y.; Li, F.-L.; Tang, Y.-X.; Shao, J.-R.; Xue, W.-T.; Wu, Y.-M. Bioactive compounds in functional buckwheat food. Food Res. Int. 2012, 49, 389–395. [Google Scholar] [CrossRef]
- Gebrelibanos, M.; Tesfaye, D.; Raghavendra, Y.; Sintayeyu, B. Nutritional and health implications of legumes. Int. J. Pharm. Sci. Res. 2013, 4, 1269. [Google Scholar]
- Gholami, A.; De Geyter, N.; Pollier, J.; Goormachtig, S.; Goossens, A. Natural product biosynthesis in Medicago species. Nat. Prod. Rep. 2014, 31, 356–380. [Google Scholar] [CrossRef]
- Soto-Zarazúa, M.G.; Bah, M.; Costa, A.S.G.; Rodrigues, F.; Pimentel, F.B.; Rojas-Molina, I.; Rojas, A.; Oliveira, M.B.P.P. Nutraceutical potential of new alfalfa (Medicago sativa) ingredients for beverage preparations. J. Med. Food 2017, 20, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, M.; Palacios, J.C.O.; McClements, D.J.; Mahfouzi, M.; Moreno, A. Alfalfa as a sustainable source of plant-based food proteins. Trends Food Sci. Technol. 2023, 135, 202–214. [Google Scholar] [CrossRef]
- Chun, O.K.; Smith, N.; Sakagawa, A.; Lee, C.Y. Antioxidant properties of raw and processed cabbages. Int. J. Food Sci. Nutr. 2004, 55, 191–199. [Google Scholar] [CrossRef]
- Guo, X.-D.; Ma, Y.-J.; Parry, J.; Gao, J.-M.; Yu, L.-L.; Wang, M. Phenolics content and antioxidant activity of tartary buckwheat from different locations. Molecules 2011, 16, 9850–9867. [Google Scholar] [CrossRef] [PubMed]
- Raeeszadeh, M.; Beheshtipour, J.; Jamali, R.; Akbari, A. The antioxidant properties of Alfalfa (Medicago sativa L.) and its biochemical, antioxidant, anti-inflammatory, and pathological effects on nicotine-induced oxidative stress in the rat liver. Oxidative Med. Cell. Longev. 2022, 2022, 2691577. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liang, K.; Jin, Y.; Zhang, M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Ofosu, F.K.; Chelliah, R.; Kim, J.-H.; Kim, J.-R.; Yoo, D.; Oh, D.-H. Untargeted metabolomics of fermented rice using UHPLC Q-TOF MS/MS reveals an abundance of potential antihypertensive compounds. Foods 2020, 9, 1007. [Google Scholar] [CrossRef]
- Ribeiro, P.V.; Tavares, J.F.; Costa, M.A.; Mattar, J.B.; Alfenas, R.C. Effect of reducing dietary advanced glycation end products on obesity-associated complications: A systematic review. Nutr. Rev. 2019, 77, 725–734. [Google Scholar] [CrossRef]
- Benito, S.; Sánchez, A.; Unceta, N.; Andrade, F.; Aldámiz-Echevarria, L.; Goicolea, M.A.; Barrio, R.J. LC-QTOF-MS-based targeted metabolomics of arginine-creatine metabolic pathway-related compounds in plasma: Application to identify potential biomarkers in pediatric chronic kidney disease. Anal. Bioanal. Chem. 2016, 408, 747–760. [Google Scholar] [CrossRef]
- Molina-Calle, M.; Priego-Capote, F.; de Castro, M.L. Characterization of Stevia leaves by LC–QTOF MS/MS analysis of polar and non-polar extracts. Food Chem. 2017, 219, 329–338. [Google Scholar] [CrossRef]
- Bence, A.K.; Worthen, D.R.; Adams, V.R.; Crooks, P.A. The antiproliferative and immunotoxic effects of L-canavanine and L-canaline. Anti-Cancer Drugs 2002, 13, 313–320. [Google Scholar] [CrossRef]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P. UPLC/ESI-Q-TOF-MS analysis of (poly) phenols, tocols and amino acids in Chaenomeles leaves versus in vitro anti-enzyme activities. Ind. Crops Prod. 2022, 181, 114829. [Google Scholar] [CrossRef]
- Aloo, S.-O.; Ofosu, F.-K.; Daliri, E.-B.-M.; Oh, D.-H. UHPLC-ESI-QTOF-MS/MS Metabolite Profiling of the Antioxidant and Antidiabetic Activities of Red Cabbage and Broccoli Seeds and Sprouts. Antioxidants 2021, 10, 852. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Vives-Peris, V.; Gómez-Cadenas, A.; Arbona, V. A fast and precise method to identify indolic glucosinolates and camalexin in plants by combining mass spectrometric and biological information. J. Agric. Food Chem. 2012, 60, 8648–8658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wei, D.; He, L.; Wang, D.; Wang, L.; Tang, D.; Zhao, R.; Ye, X.; Wu, C.; Peng, W. Application of response surface methodology (RSM) for optimization of the supercritical CO2 extract of oil from Zanthoxylum bungeanum pericarp: Yield, composition and gastric protective effect. Food Chem. X 2022, 15, 100391. [Google Scholar] [CrossRef] [PubMed]
- de la Luz Cádiz-Gurrea, M.; Fernández-Arroyo, S.; Joven, J.; Segura-Carretero, A. Comprehensive characterization by UHPLC-ESI-Q-TOF-MS from an Eryngium bourgatii extract and their antioxidant and anti-inflammatory activities. Food Res. Int. 2013, 50, 197–204. [Google Scholar] [CrossRef]
- Wang, H.; Ye, Y.H.; Wang, H.H.; Liu, J.; Liu, Y.J.; Jiang, B.W. HPLC-QTOF-MS/MS profiling, antioxidant, and α-glucosidase inhibitory activities of Pyracantha fortuneana fruit extracts. J. Food Biochem. 2019, 43, e12821. [Google Scholar] [CrossRef]
- Li, H.; Zhang, D.; Tan, L.-H.; Yu, B.; Zhao, S.-P.; Cao, W.-G. Comparison of the antioxidant properties of various solvent extracts from Dipsacus asperoides and identification of phenolic compounds by LC-ESI-QTOF-MS–MS. S. Afr. J. Bot. 2017, 109, 1–8. [Google Scholar] [CrossRef]
- Nadeem, M.; Mumtaz, M.W.; Danish, M.; Rashid, U.; Mukhtar, H.; Irfan, A.; Anwar, F.; Saari, N. UHPLC-QTOF-MS/MS metabolites profiling and antioxidant/antidiabetic attributes of Cuscuta reflexa grown on Casearia tomentosa: Exploring phytochemicals role via molecular docking. Int. J. Food Prop. 2020, 23, 918–940. [Google Scholar] [CrossRef]
- Maciejewska-Turska, M.; Zgórka, G. In-depth phytochemical and biological studies on potential AChE inhibitors in red and zigzag clover dry extracts using reversed–phase liquid chromatography (RP-LC) coupled with photodiode array (PDA) and electron spray ionization-quadrupole/time of flight-mass spectrometric (ESI-QToF/MS-MS) detection and thin-layer chromatography-bioautography. Food Chem. 2022, 375, 131846. [Google Scholar]
- Oszmiański, J.; Kolniak-Ostek, J.; Wojdyło, A. Application of ultra performance liquid chromatography-photodiode detector-quadrupole/time of flight-mass spectrometry (UPLC-PDA-Q/TOF-MS) method for the characterization of phenolic compounds of Lepidium sativum L. sprouts. Eur. Food Res. Technol. 2013, 236, 699–706. [Google Scholar] [CrossRef]
- Tsednee, M.; Mak, Y.W.; Chen, Y.R.; Yeh, K.C. A sensitive LC-ESI-Q-TOF-MS method reveals novel phytosiderophores and phytosiderophore–iron complexes in barley. New Phytol. 2012, 195, 951–961. [Google Scholar] [CrossRef]
- Yao, W.; Li, H.; Liu, Q.; Gao, Y.; Dai, J.; Bao, B.; Zhang, L.; Ding, A. Cellular metabolomics revealed the cytoprotection of amentoflavone, a natural compound, in lipopolysaccharide-induced injury of human umbilical vein endothelial cells. Int. J. Mol. Sci. 2016, 17, 1514. [Google Scholar] [CrossRef]
- Guo, Q.; Sun, Y.; Tang, Q.; Zhang, H.; Cheng, Z. Isolation, identification, biological estimation, and profiling of glucosinolates in Isatis indigotica roots. J. Liq. Chromatogr. Relat. Technol. 2020, 43, 645–656. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Wei, S.; Li, Y.; Li, W.; Yan, M.; Deng, Y.; Zhang, B.; Cai, H. Identification and analysis of components in Shen-Fu-Shu granule extract and in rat plasma after oral administration by UPLC-ESI/Q-TOF-MS. J. Pharm. Biomed. Anal. 2019, 169, 159–169. [Google Scholar] [CrossRef]
- Song, J.; Liu, C.; Li, D.; Gu, Z. Evaluation of sugar, free amino acid, and organic acid compositions of different varieties of vegetable soybean (Glycine max [L.] Merr). Ind. Crops Prod. 2013, 50, 743–749. [Google Scholar] [CrossRef]
- Salunkhe, D.; Jadhav, S.; Kadam, S.; Chavan, J.; Luh, B. Chemical, biochemical, and biological significance of polyphenols in cereals and legumes. Crit. Rev. Food Sci. Nutr. 1983, 17, 277–305. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Singh, B.; Gupta, M. Antioxidant and prebiotic potential of Murraya koenigii and Brassica oleracea var. botrytis leaves as food ingredient. J. Agric. Food Res. 2020, 2, 100069. [Google Scholar] [CrossRef]
- Zhang, Y.; Hao, R.; Chen, J.; Li, S.; Huang, K.; Cao, H.; Farag, M.A.; Battino, M.; Daglia, M.; Capanoglu, E. Health benefits of saponins and its mechanisms: Perspectives from absorption, metabolism, and interaction with gut. Crit. Rev. Food Sci. Nutr. 2023, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Tabart, J.; Pincemail, J.; Kevers, C.; Defraigne, J.-O.; Dommes, J. Processing effects on antioxidant, glucosinolate, and sulforaphane contents in broccoli and red cabbage. Eur. Food Res. Technol. 2018, 244, 2085–2094. [Google Scholar] [CrossRef]
- Şengül, M.; Yildiz, H.; Kavaz, A. The effect of cooking on total polyphenolic content and antioxidant activity of selected vegetables. Int. J. Food Prop. 2014, 17, 481–490. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical activity and polyphenol composition of local Brassicaceae edible varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Qin, P.; Wang, Q.; Shan, F.; Hou, Z.; Ren, G. Nutritional composition and flavonoids content of flour from different buckwheat cultivars. Int. J. Food Sci. Technol. 2010, 45, 951–958. [Google Scholar] [CrossRef]
- Graziano, S.; Agrimonti, C.; Marmiroli, N.; Gullì, M. Utilisation and limitations of pseudocereals (quinoa, amaranth, and buckwheat) in food production: A review. Trends Food Sci. Technol. 2022, 125, 154–165. [Google Scholar] [CrossRef]
- Szumacher-Strabel, M.; Stochmal, A.; Cieslak, A.; Kozłowska, M.; Kuznicki, D.; Kowalczyk, M.; Oleszek, W. Structural and quantitative changes of saponins in fresh alfalfa compared to alfalfa silage. J. Sci. Food Agric. 2019, 99, 2243–2250. [Google Scholar] [CrossRef] [PubMed]
- Radünz, M.; Camargo, T.M.; dos Santos Hackbart, H.C.; Blank, J.P.; Hoffmann, J.F.; Stefanello, F.M.; da Rosa Zavareze, E. Encapsulation of broccoli extract by electrospraying: Influence of in vitro simulated digestion on phenolic and glucosinolate contents, and on antioxidant and antihyperglycemic activities. Food Chem. 2021, 339, 128075. [Google Scholar] [CrossRef]
- Živković, A.; Polak, T.; Cigić, B.; Požrl, T. Germinated buckwheat: Effects of dehulling on phenolics profile and antioxidant activity of buckwheat seeds. Foods 2021, 10, 740. [Google Scholar] [CrossRef]
- Liu, S.; Li, D.; Huang, B.; Chen, Y.; Lu, X.; Wang, Y. Inhibition of pancreatic lipase, α-glucosidase, α-amylase, and hypolipidemic effects of the total flavonoids from Nelumbo nucifera leaves. J. Ethnopharmacol. 2013, 149, 263–269. [Google Scholar] [CrossRef]
- Qin, P.; Wu, L.; Yao, Y.; Ren, G. Changes in phytochemical compositions, antioxidant and α-glucosidase inhibitory activities during the processing of tartary buckwheat tea. Food Res. Int. 2013, 50, 562–567. [Google Scholar] [CrossRef]
- Podsedek, A.; Majewska, I.; Kucharska, A.Z. Inhibitory potential of red cabbage against digestive enzymes linked to obesity and type 2 diabetes. J. Agric. Food Chem. 2017, 65, 7192–7199. [Google Scholar] [CrossRef]
- Sotokawauchi, A.; Ishibashi, Y.; Matsui, T.; Yamagishi, S.-I. Aqueous extract of glucoraphanin-rich broccoli sprouts inhibits formation of advanced glycation end products and attenuates inflammatory reactions in endothelial cells. Evid.-Based Complement. Altern. Med. 2018, 2018, 9823141. [Google Scholar] [CrossRef]
- Buko, V.; Zavodnik, I.; Kanuka, O.; Belonovskaya, E.; Naruta, E.; Lukivskaya, O.; Kirko, S.; Budryn, G.; Żyżelewicz, D.; Oracz, J. Antidiabetic effects and erythrocyte stabilization by red cabbage extract in streptozotocin-treated rats. Food Funct. 2018, 9, 1850–1863. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, Y.; Wang, X.; Dai, W.; Zhang, L.; Piao, C. Flavonoids derived from buckwheat hull can break advanced glycation end-products and improve diabetic nephropathy. Food Funct. 2021, 12, 7161–7170. [Google Scholar] [CrossRef]
- Starowicz, M.; Zieliński, H. Inhibition of advanced glycation end-product formation by high antioxidant-leveled spices commonly used in European cuisine. Antioxidants 2019, 8, 100. [Google Scholar] [CrossRef]
- Bunning, B.J.; Contrepois, K.; Lee-McMullen, B.; Dhondalay, G.K.R.; Zhang, W.; Tupa, D.; Raeber, O.; Desai, M.; Nadeau, K.C.; Snyder, M.P. Global metabolic profiling to model biological processes of aging in twins. Aging cell 2020, 19, e13073. [Google Scholar] [CrossRef]
- Quiroga, P.R.; Nepote, V.; Baumgartner, M.T. Contribution of organic acids to α-terpinene antioxidant activity. Food Chem. 2019, 277, 267–272. [Google Scholar] [CrossRef]
- Yoshinari, O.; Igarashi, K. Anti-diabetic effect of pyroglutamic acid in type 2 diabetic Goto-Kakizaki rats and KK-Ay mice. Br. J. Nutr. 2011, 106, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, Y.-L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Zukalová, H.; Vasak, J. The role and effects of glucosinolates of Brassica species-a review. Rostl. Vyrob. 2002, 48, 175–180. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Metabolites | RT Per Min | [M − H]− (m/z) | Molecular Formula | Alfalfa | Buckwheat | Broccoli | Red Cabbage | Fragment Pattern | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Amino acids | ||||||||||
| 1 | L-Arginine | 0.7 | 173.10 | C6H14N4O2 | 77,000 | 65,000 | 110,000 | 52,000 | 173 | [20] |
| 2 | Threonine | 0.69 | 118.05 | C4H9NO3 | 190,000 | 0.00 | 0.00 | 0.00 | 56, 74 | [21] |
| 3 | Canavanine | 0.7 | 175.08 | C5H12N4O3 | 310,000 | 0.00 | 0.00 | 0.00 | 86, 118, 130 | [22] |
| 4 | Ornithine | 0.71 | 131.08 | C5H12N2O2 | 0.00 | 0.00 | 63,000 | 0.00 | 131, 88 | [23] |
| 5 | Serine | 0.77 | 104.04 | C3H7NO3 | 0.00 | 0.00 | 29,000 | 0.00 | 44, 60 | [21] |
| 6 | DL-Homoserine | 0.81 | 118.05 | C4H9NO3 | 190,000 | 0.00 | 29,000 | 0.00 | 74, 100 | [24] |
| 7 | L-Glutamate | 0.82 | 146.04 | C5H9NO4 | 220,000 | 220,000 | 24,000 | 0.00 | 102, 146 | [23] |
| 8 | L-Asparagine | 0.78 | 131.05 | C4H8N2O3 | 54,000.00 | 0.00 | 130,000 | 54,000 | 58, 113 | [23] |
| 9 | Pyroglutamic acid | 1.05 | 128.04 | C5H7NO3 | 48,000 | 69,000 | 160,000 | 100,000 | 289, 72 | METLIN database |
| 10 | N-Methylglutamic acid | 1.18 | 160.06 | C6H11NO4 | 45,000 | 0.00 | 0.00 | 0.00 | 58, 142 | METLIN database |
| 11 | 2-Aminomuconic acid | 1.21 | 240.05 | C6H7N7O4 | 0.00 | 0.00 | 27,000 | 0.00 | 128 | METLIN databases |
| 12 | L-Phenylalanine | 2.98 | 164.07 | C9H11NO2 | 0.00 | 0.00 | 72,000 | 0.00 | 77, 103 | [23] |
| 13 | L-Tryptophan | 5.9 | 203.08 | C11H12N2O2 | 0.00 | 68,000 | 0.00 | 0.00 | 205, 188, 118, 91, 116, 142 | [23,25] |
| 14 | Aspartame | 8.99 | 293.11 | C14H18N2O5 | 470,000 | 0.00 | 0.00 | 0.00 | - | METLIN databases |
| 15 | Pivagabine | 15.34 | 186.11 | C9H17NO3 | 0.00 | 0.00 | 78,000 | 0.00 | [26] | |
| Polyphenols | ||||||||||
| 16 | Glycogallin | 2.48 | 331.07 | C13H16O10 | 0.00 | 53,000 | 0.00 | 0.00 | 168 | [27] |
| 17 | Protocatechuic acid hexoside (isomer) | 4.03 | 315.07 | C13H16O9 | 0.00 | 0.00 | 130,000 | 100,000 | 163, 152 | [28] |
| 18 | Catechin | 9.68 | 289.07 | C15H14O6 | 0.00 | 350,000 | 0.00 | 68,000 | 245, 203, 151, 137, 109 | [28] |
| 19 | Ferulic acid | 10.89 | 193.05 | C10H10O4 | 79,000 | 0.00 | 0.00 | 0.00 | 193, 151 | [29] |
| 20 | Caffeic acid 3-glucoside | 11.09 | 341.09 | C15H18O9 | 0.00 | 190,000 | 0.00 | 0.00 | 179, 281, 221, 161, 135 | [30] |
| 21 | Sinapoylglucose | 13.66 | 385.11 | C17H22O10 | 0.00 | 120,000 | 620,000 | 1,300,000 | 223, 208, 179 164 | [28] |
| 22 | Phloretin | 15.99 | 273.08 | C15H14O5 | 0.00 | 50,000 | 0.00 | 0.00 | 255, 149 137 | [31] |
| 23 | (-)-Catechin gallate | 16.24 | 441.08 | C22H18O10 | 0.00 | 310,000 | 0.00 | 0.00 | 125, 124, 145, 303 | [24] |
| 24 | Polygalic acid | 16.67 | 487.31 | C29H44O6 | 250,000 | 330,000 | 320,000 | 420,000 | 61, 174, 239 | METLIN database |
| 25 | Sinapic acid | 16.75 | 223.061 | C11H12O5 | 0.00 | 0.00 | 110,000 | 75,000 | 121, 149, 164, 179, 223, 193, 208 | [32] |
| Organic acid | ||||||||||
| 26 | Gluconic acid | 0.83 | 195.05 | C6H12O7 | 0.00 | 640,000 | 370,000 | 540,000 | 177, 159, 129 | [27] |
| 27 | Arabinonic acid | 0.85 | 165.04 | C5H10O6 | 0.00 | 0.00 | 0.00 | 180,000 | - | - |
| 28 | Fumaric acid | 1.04 | 115.00 | C4H4O4 | 96,000 | 120,000 | 0.00 | 0.00 | - | - |
| 29 | Malic acid | 1.05 | 134.02 | C4H6O5 | 300,000 | 380,000 | 0.00 | 0.00 | 71, 133, 59 | METLIN database |
| 30 | Uric acid | 1.21 | 168.03 | C5H4N4O3 | 150,000 | 0.00 | 0.00 | 0.00 | 168 | METLIN database |
| 31 | Lactic acid | 1.21 | 89.02 | C3H6O3 | 45,000 | 0.00 | 70,000 | 43,000 | 89 | METLIN database |
| 32 | Citric acid | 1.22 | 191.02 | C6H8O7 | 770,000 | 790,000 | 570,000 | 510,000 | 173,155, 131, 129. 127, 111 191 | [30] |
| 33 | 2-Furoic acid | 1.23 | 111.01 | C5H4O3 | 250,000 | 260,000 | 180,000 | 170,000 | 65, 70 | [24] |
| Other compounds | ||||||||||
| 34 | Nicotianamine | 0.76 | 303.14 | C12H21N3O6 | 0.00 | 0.00 | 0.00 | 24,000 | 305, 287, 241, 169, 114 | [33] |
| 35 | Camalexin | 0.77 | 200.04 | C11H8N2S | 0.00 | 0.00 | 45,000 | 0.00 | 201, 160, 142, 59 | [25] |
| 36 | Dihydrouracil | 0.77 | 114.04 | C4H6N2O2 | 0.00 | 0.00 | 48,000 | 0.00 | 111, 112, 113 | [34] |
| 37 | Glucoraphanin | 1.02 | 436.04 | C12H23NO10S3 | 0.00 | 0.00 | 19,000,000 | 0.00 | 291, 275, 259, 195, 97 | [35] |
| 38 | Epiprogoitrin | 1.08 | 388.04 | C11H19NO10S2 | 0.00 | 0.00 | 2,600,000 | 14,000,000 | 96, 74, 95, 274, 90, 135,259, 149 | [35] |
| 39 | Robinose | 1.09 | 326.12 | C12H22O10 | 0.00 | 100,000 | 0.00 | 0.00 | - | - |
| 40 | Sinigrin | 1.12 | 358.03 | C10H17NO9S2 | 0.00 | 0.00 | 1,200,000 | 5,300,000 | 74, 95, 96, 274 | [35] |
| 41 | Uridine | 1.22 | 243.06 | C9H12N2O6 | 100,000 | 0.00 | 0.00 | 0.00 | 110, 82, 122, 66 | |
| 42 | Glucoalyssin | 1.21 | 450.06 | C13H25NO10S3 | 0.00 | 0.00 | 200,000 | 0.00 | 96, 95, 192, 165, 256, 386 | [35] |
| 43 | Adenosine | 1.23 | 268.10 | C10H13N5O4 | 560,000 | 370,000 | 0.00 | 0.00 | 136, 268 | |
| 44 | Gluconapin | 1.9 | 372.04 | C11H19NO9S2 | 0.00 | 0.00 | 590,000 | 2,600,000 | 74, 96, 119, 174, 274, 372 | [35] |
| 45 | 4-Sulfophthalic anhydride | 2.31 | 227.97 | C8H4O6S | 920 | 0.00 | 2600 | 5000 | - | - |
| 46 | 2-Benzoylmalononitrile | 2.93 | 170.05 | C10H6N2O | 0.00 | 0.00 | 0.00 | 290,000 | - | - |
| 47 | 2 or 3-Methylbutyl glucosinolate | 6.14 | 388.07 | C12H23NO9S2 | 0.00 | 0.00 | 470,000 | 0.00 | 74, 95, 96, 259, 388 | [35] |
| 48 | Emiglitate | 10.42 | 355.16 | C17H25NO7 | 0.00 | 0.00 | 2,400,000 | 2,300,000 | 93, 119,147, 355 | METLIN database |
| 49 | Actinonin | 13.33 | 384.25 | C19H35N3O5 | 0.00 | 420,000 | 380,000 | 380,000 | 111, 180, 224 | [24] |
| 50 | 7-Caffeoylsedoheptulose | 14.88 | 372.11 | C16H20O10 | 0.00 | 400,000 | 0.00 | 0.00 | 77, 121, 175, 249 | METLIN database |
| 51 | Capparidisine | 16.11 | 495.24 | C27H33N3O6 | 0.00 | 0.00 | 830,000 | 0.00 | - | - |
| 52 | Quinadoline A | 16.27 | 485.21 | C27H27N5O4 | 0.00 | 0.00 | 56,000 | 0.00 | - | - |
| 53 | 1-Hexadecylamine | 17.00 | 242.29 | C16H35N | 490,000 | 540,000 | 0.00 | 0.00 | 136, 268 | [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aloo, S.O.; Ofosu, F.K.; Muchiri, M.N.; Vijayalakshmi, S.; Pyo, C.-G.; Oh, D.-H. In Vitro Bioactivities of Commonly Consumed Cereal, Vegetable, and Legume Seeds as Related to Their Bioactive Components: An Untargeted Metabolomics Approach Using UHPLC–QTOF-MS2. Antioxidants 2023, 12, 1501. https://doi.org/10.3390/antiox12081501
Aloo SO, Ofosu FK, Muchiri MN, Vijayalakshmi S, Pyo C-G, Oh D-H. In Vitro Bioactivities of Commonly Consumed Cereal, Vegetable, and Legume Seeds as Related to Their Bioactive Components: An Untargeted Metabolomics Approach Using UHPLC–QTOF-MS2. Antioxidants. 2023; 12(8):1501. https://doi.org/10.3390/antiox12081501
Chicago/Turabian StyleAloo, Simon Okomo, Fred Kwame Ofosu, Mary Njeri Muchiri, Selvakumar Vijayalakshmi, Choi-Geun Pyo, and Deog-Hwan Oh. 2023. "In Vitro Bioactivities of Commonly Consumed Cereal, Vegetable, and Legume Seeds as Related to Their Bioactive Components: An Untargeted Metabolomics Approach Using UHPLC–QTOF-MS2" Antioxidants 12, no. 8: 1501. https://doi.org/10.3390/antiox12081501