

Treatment with Pterostilbene Ameliorates the Antioxidant Status of Bovine Spermatozoa and Modulates Cell Death Pathways

 ,
,  ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Semen Samples
2.2. Measurement of Lipid Peroxidation
2.3. Determination of Total Antioxidant Capacity (TAC)
2.4. Determination of Intracellular Glutathione (GSH)
2.5. Determination of ATP and AMP
2.6. SDS-PAGE/Immunoblot and Dot Blot Analysis
2.7. Statistics
3. Results
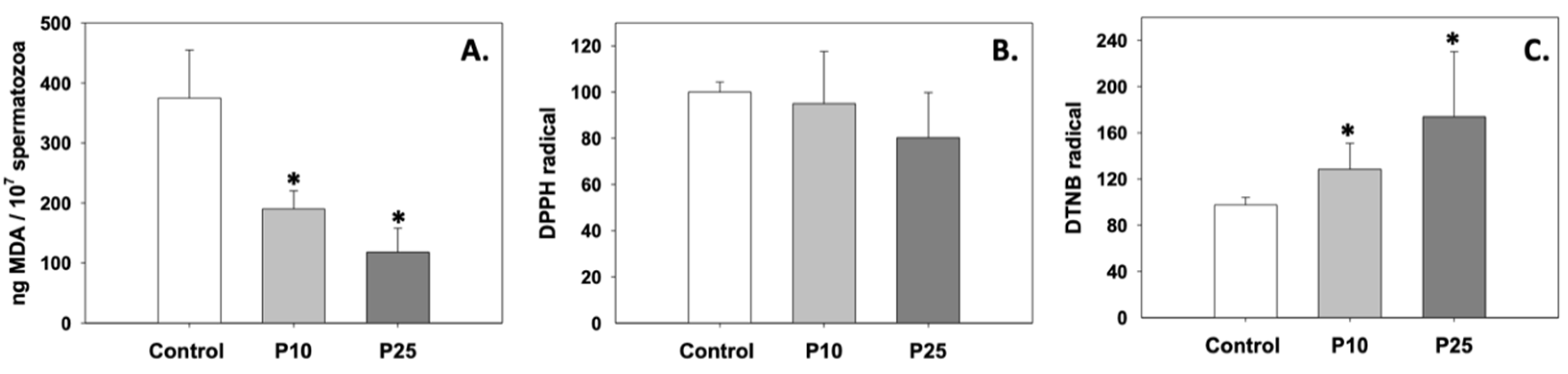
3.1. Oxidative Stress/Antioxidant Status
3.2. Apoptosis
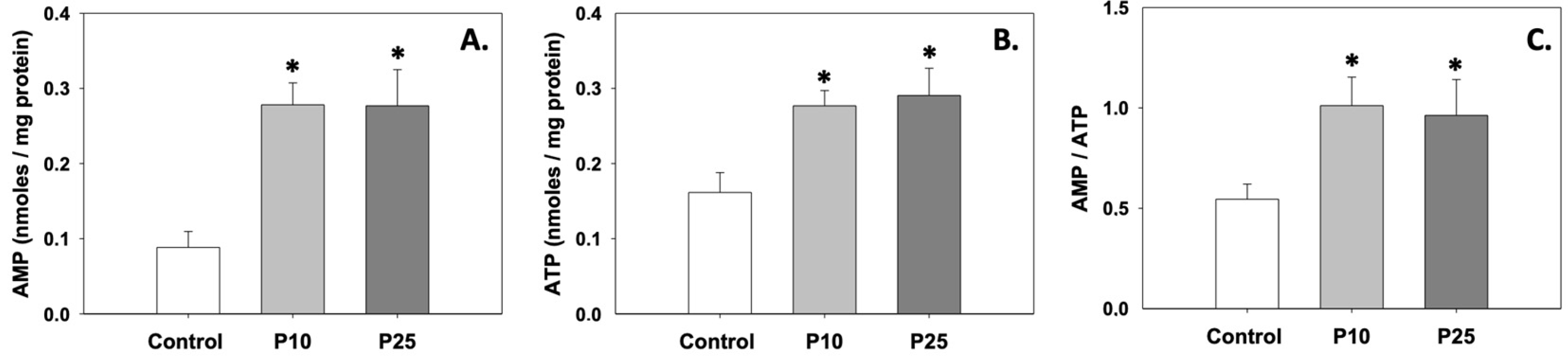
3.3. Energy Content
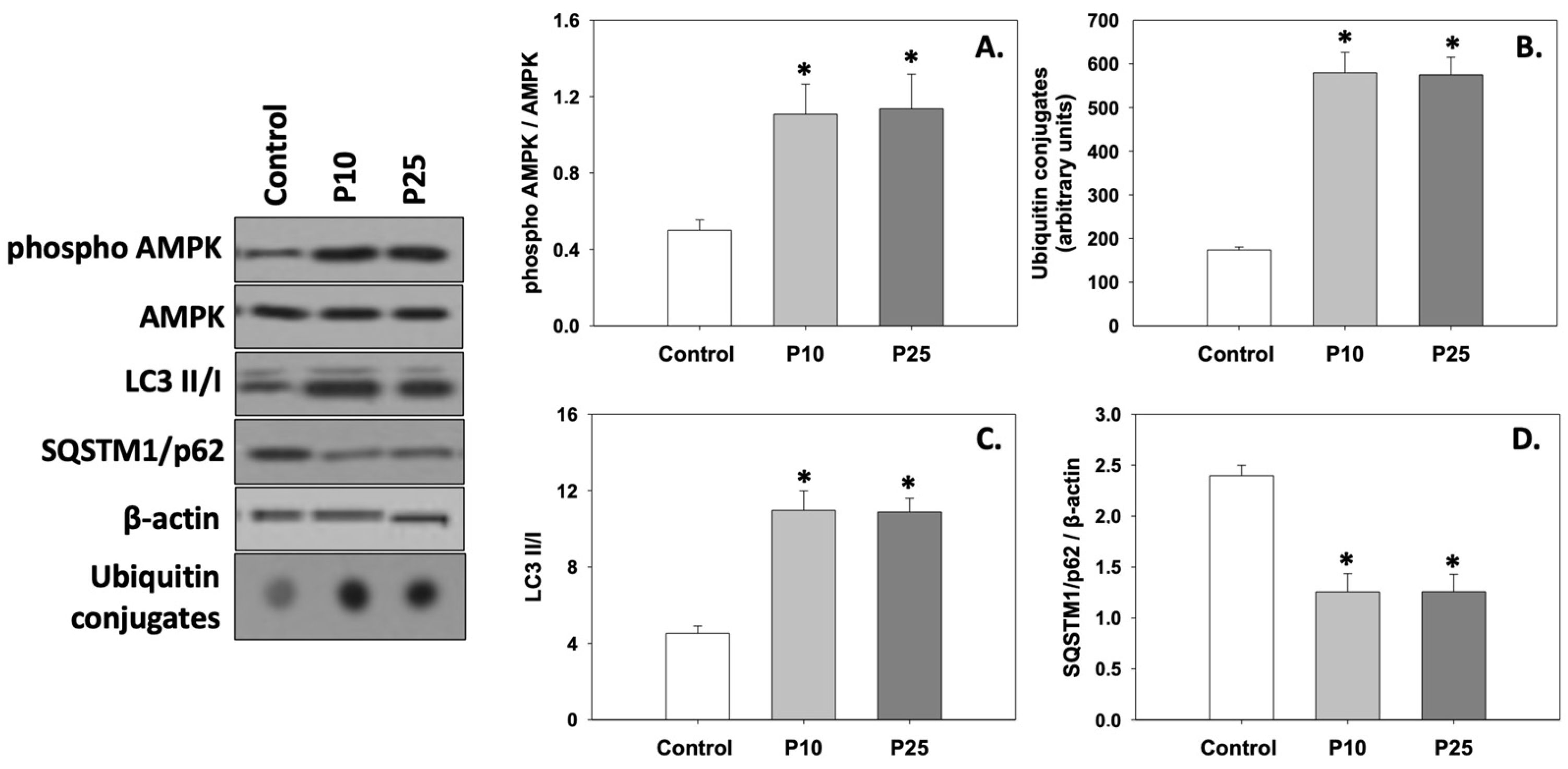
3.4. AMPK Phosphorylation
3.5. Autophagy
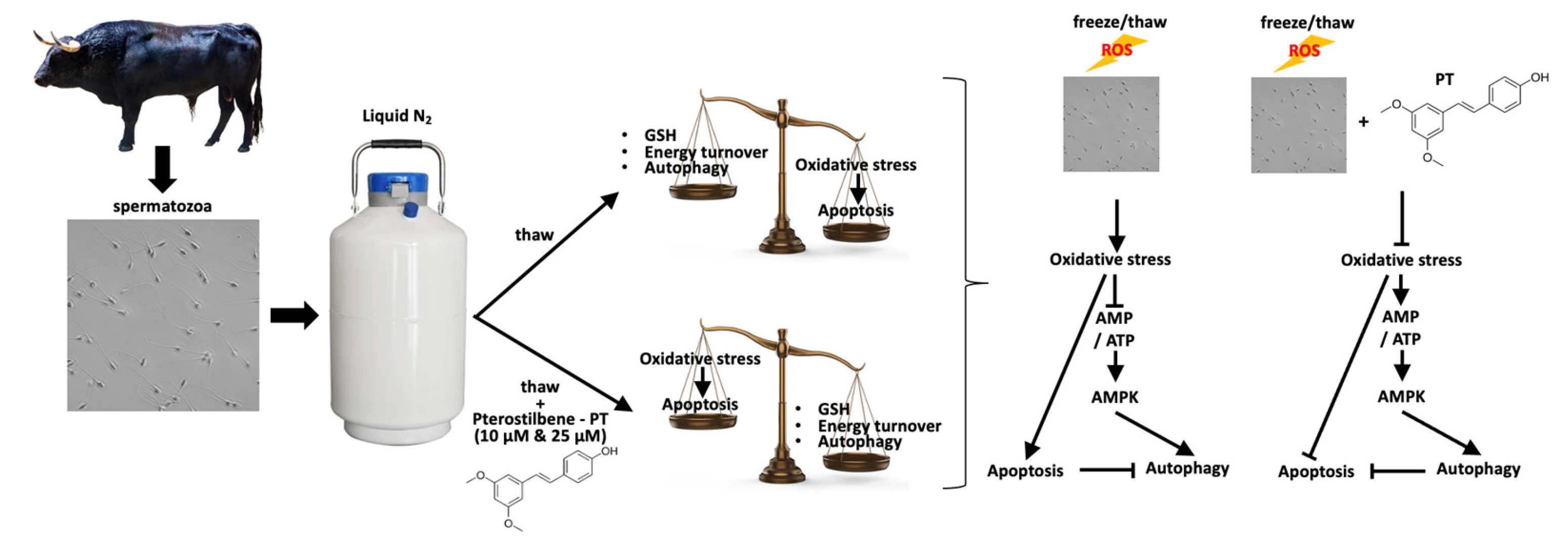
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Flaherty, C.; Beorlegui, N.; Beconi, M.T. Participation of superoxide anion in the capacitation of cryopreserved bovine sperm. Int. J. Androl. 2003, 26, 109–114. [Google Scholar] [CrossRef]
- Visconti, P.E. Understanding the molecular basis of sperm capacitation through kinase design. Proc. Natl. Acad. Sci. USA 2009, 10, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Dodson, M.; Darley-Usmar, V.; Zhang, J. Cellular Metabolic and Autophagic Pathways: Traffic Control by Redox Signaling. Free Rad. Biol. Med. 2013, 63, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Hardie, G.; Schaffer, B.; Brunet, A. AMPK: An energy-sensing pathway with multiple inputs and outputs. Trends Cell. Biol. 2016, 26, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, X. Melatonin and male reproduction. Clin. Chim. Acta 2015, 446, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.P.; Majzoub, A.; Agarwal, A. Antioxidants Use and Sperm DNA Damage. In Male Infertility; Parekattil, S., Esteves, S., Agarwal, A., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Sapanidou, V.; Tsantarliotou, M.P.; Lavrentiadou, S.N. A review of the use of antioxidants in bovine sperm preparation protocols. Anim. Reprod. Sci. 2023, 251, 107215. [Google Scholar] [CrossRef]
- Sapanidou, V.; Tsantarliotou, M.; Lavrentiadou, S.; Tzekaki, E.; Efraimidis, I.; Lialiaris, T.; Asimakopoulos, B. The beneficial effects of pterostilbene on post-thawed bovine spermatozoa. Animals 2023, 13, 2713. [Google Scholar] [CrossRef]
- Sapanidou, V.G.; Asimakopoulos, B.; Lialiaris, T.; Lavrentiadou, S.N.; Feidantsis, K.; Kourousekos, G.; Tsantarliotou, M.P. The Role of Erythropoietin in Bovine Sperm Physiology. Animals 2024, 14, 2175. [Google Scholar] [CrossRef]
- Niizuma, K.; Yoshioka, H.; Chen, H.; Kim, G.S.; Jung, J.E.; Katsu, M.; Okami, N.; Chan, P.H. Mitochondrial and apoptotic neuronal death signaling pathways in cerebral ischemia. Biochim. Biophys. Acta 2010, 1802, 92–99. [Google Scholar] [CrossRef]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; et al. A ubiquitin-like system mediates protein lipidation. Nature 2000, 408, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.A.; Outzen, H.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–24145. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, M.; Waguri, S.; Koike, M.; Sou, Y.S.; Ueno, T.; Hara, T.; Mizushima, N.; Iwata, J.; Ezaki, J.; Murata, S.; et al. Homeostatic levels of p62 control cytoplasmic inclusion body formation in autophagy-deficient mice. Cell 2007, 31, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial membrane permeabilization in cell death. Physiol. Rev. 2007, 87, 99–163. [Google Scholar] [CrossRef] [PubMed]
- Hockenbery, D.M. bcl-2, a novel regulator of cell death. BioEssays 1995, 17, 631–638. [Google Scholar] [CrossRef]
- Takle, H.; Andersen, Ø. Caspases and apoptosis in fish. J. Fish. Biol. 2007, 71, 326–349. [Google Scholar] [CrossRef]
- Riedl, S.J.; Shi, Y. Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol. 2004, 5, 897–907. [Google Scholar] [CrossRef]
- Aitken, R.J.; Koppers, A.J. Apoptosis and DNA damage in human spermatozoa. Asian J. Androl. 2011, 13, 36. [Google Scholar] [CrossRef]
- Aparicio, I.M.; Espino, J.; Bejarano, I.; Gallardo-Soler, A.; Campo, M.L.; Salido, G.M.; Pariente, J.A.; Peña, F.J.; Tapia, J.A. Autophagy-related proteins are functionally active in human spermatozoa and may be involved in the regulation of cell survival and motility. Sci. Rep. 2016, 6, 33647. [Google Scholar] [CrossRef]
- Gallardo Bolaños, J.M.; Miró Morán, Á.; Balao da Silva, C.M.; Morillo Rodríguez, A.; Plaza Dávila, M.; Aparicio, I.M.; Tapia, J.A.; Ferrusola, C.O.; Peña, F.J. Autophagy and apoptosis have a role in the survival or death of stallion spermatozoa during conservation in refrigeration. PLoS ONE 2012, 7, e30688. [Google Scholar] [CrossRef]
- Uribe, P.; Merino, J.; Matus, C.E.; Schulz, M.; Zambrano, F.; Villegas, J.V.; Conejeros, I.; Taubert, A.; Hermosilla, C.; Sánchez, R. Autophagy is activated in human spermatozoa subjected to oxidative stress and its inhibition impairs sperm quality and promotes cell death. Hum. Reprod. 2022, 37, 680–695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jiang, M.; Bi, Y.; Zhu, H.; Zhou, Z.; Sha, J. Autophagy and apoptosis act as partners to induce germ cell death after heat stress in mice. PLoS ONE 2012, 7, e41412. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Zhang, Y.; Wang, Q.; Yuan, L. Autophagy: A double-edged sword in male reproduction. Int. J. Mol. Sci. 2022, 23, 15273. [Google Scholar] [CrossRef] [PubMed]
- Correa, J.R.; Pace, M.M.; Zavos, P.M. Relationships among frozen-thawed sperm characteristics assessed via the routine semen analysis, sperm functional tests and fertility of bulls in an artificial insemination program. Theriogenology 1997, 48, 721–731. [Google Scholar] [CrossRef]
- Hoshi, H. In vitro production of bovine embryos and their application for embryo transfer. Theriogenology 2003, 59, 675–685. [Google Scholar] [CrossRef]
- Chatterjee, S.; Gagnon, C. Production of reactive oxygen species by spermatozoa undergoing cooling, freezing, and thawing. Mol. Reprod. Dev. 2001, 59, 451–458. [Google Scholar] [CrossRef]
- Al-Gubory, K.H.; Fowler, P.A.; Garrel, C. The roles of cellular reactive oxygen species, oxidative stress and antioxidants in pregnancy outcomes. Int. J. Biochem. Cell Biol. 2010, 42, 1634–1650. [Google Scholar] [CrossRef]
- Chaudière, J. Some chemical and biochemical constraints of oxidative stress in living cells. In New Comprehensive Biochemistry, Free Radical Damage and Its Control; Rice-Evans, C.A., Burdon, R.H., Eds.; Elsevier: Amsterdam, The Netherlands, 1994; Volume 28, pp. 25–66. [Google Scholar]
- Sanocka, D.; Kurpisz, M. Reactive oxygen species and sperm cells. Reprod. Biol. Endocrinol. 2004, 2, 12. [Google Scholar] [CrossRef]
- Remsberg, C.M.; Yáñez, J.A.; Ohgami, Y.; Vega-Villa, K.R.; Rimando, A.M.; Davies, N.M. Pharmacometrics of pterostilbene: Preclinical pharmacokinetics and metabolism, anticancer, antiinflammatory, antioxidant and analgesic activity. Phytother. Res. 2008, 22, 169–179. [Google Scholar] [CrossRef]
- Perecko, T.; Jancinova, V.; Drábiková, K.; Nosal’, R.; Harmatha, J. Structure-efficiency relationship in derivatives of stilbene. Comparison of resveratrol, pinosylvin and pterostilbene. Neuroendocrinol. Lett. 2008, 29, 802. [Google Scholar]
- Amarnath Satheesh, M.; Pari, L. The antioxidant role of pterostilbene in streptozotocin-nicotinamide-induced type 2 diabetes mellitus in Wistar rats. J. Pharm. Pharmacol. 2006, 58, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- McCormack, D.; McFadden, D. A review of pterostilbene antioxidant activity and disease modification. Oxidative Med. Cell. Longev. 2013, 2013, 575482. [Google Scholar] [CrossRef] [PubMed]
- Robb, E.L.; Stuart, J.A. The stilbenes resveratrol, pterostilbene and piceid affect growth and stress resistance in mammalian cells via a mechanism requiring estrogen receptor beta and the induction of Mn-superoxide dismutase. Phytochemistry 2014, 98, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Messina, F.; Guglielmini, G.; Curini, M.; Orsini, S.; Gresele, P.; Marcotullio, M.C. Effect of substituted stilbenes on platelet function. Fitoterapia 2015, 105, 228–233. [Google Scholar] [CrossRef]
- Li, J.; Deng, R.; Hua, X.; Zhang, L.; Lu, F.; Coursey, T.G.; Pflugfelder, S.C.; Li, D.Q. Blueberry Component Pterostilbene Protects Corneal Epithelial Cells from Inflammation via Anti-oxidative Pathway. Sci. Rep. 2016, 6, 19408. [Google Scholar] [CrossRef]
- Zhou, J.; Ci, X.; Ma, X.; Yu, Q.; Cui, Y.; Zhen, Y.; Li, S. Pterostilbene Activates the Nrf2-Dependent Antioxidant Response to Ameliorate Arsenic-Induced Intracellular Damage and Apoptosis in Human Keratinocytes. Front. Pharmacol. 2019, 10, 497. [Google Scholar] [CrossRef]
- Sosa, F.; Romo, S.; Kjelland, M.E.; Álvarez-Gallardo, H.; Pérez-Reynozo, S.; Urbán-Duarte, D.; De La Torre-Sánchez, J.F. Effect of pterostilbene on development, equatorial lipid accumulation and reactive oxygen species production of in vitro-produced bovine embryos. Reprod. Domest. Anim. 2020, 55, 1490–1500. [Google Scholar] [CrossRef]
- Yoshida, M. Conservation of sperms: Current status and new trends. Anim. Reprod. Sci. 2000, 60, 349–355. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, sensitive, and specific thiobarbituric acid method for measuring lipid peroxidation in animal tissue, food, and feedstuff samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Janaszewska, A.; Bartosz, G. Assay of total antioxidant capacity: Comparison of four methods as applied to human blood plasma. Scand. J. Clin. Lab Investig. 2002, 62, 231–236. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Petrotos, K.; Stagos, D.; Gerasopoulos, K.; Maimaris, A.; Makris, H.; Kafantaris, I.; Makri, S.; Kerasioti, E.; Halabalaki, M.; et al. Enhancement of Antioxidant Mechanisms and Reduction of Oxidative Stress in Chickens after the Administration of Drinking Water Enriched with Polyphenolic Powder from Olive Mill Waste Waters. Oxidative Med. Cell. Longev. 2017, 2017, 8273160. [Google Scholar] [CrossRef]
- Söderquist, L.; Stålhammar, E.M. Relationship between ATP content and motility in bovine spermatozoa with reference to the effects of the bull and the AI centre. Acta Vet. Scand. 1991, 32, 353. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, G.; Yang, L.; Gajewski, C.D.; Mattiazzi, M. Measurements of ATP in mammalian cells. Methods 2002, 26, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Bock, C.; Pörtner, H.O. Resistance to freshwater exposure in White Sea littorina spp. I: Anaerobic metabolism and energetics. J. Comp. Physiol. B 2000, 170, 91–103. [Google Scholar] [CrossRef]
- Dickinson, G.H.; Ivanina, A.V.; Matoo, O.B.; Pörtner, H.O.; Lannig, G.; Bock, C.; Beniash, E.; Sokolova, I.M. Interactive effects of salinity and elevated CO2 levels on juvenile eastern oysters, Crassostrea virginica. J. Exp. Biol. 2012, 215, 29–43. [Google Scholar] [CrossRef]
- Adam, H. Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Passonneau, J.V.; Lowry, O.H. Enzymatic Analysis: A Practical Guide; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, Ι.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Zervos, I.; Tsantarliotou, M. Antioxidant effect of crocin on bovine sperm quality and in vitro fertilization. Theriogenology 2015, 84, 1273–1282. [Google Scholar] [CrossRef]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Lavrentiadou, S.; Tsantarliotou, M. Protective effect of crocetin on bovine spermatozoa against oxidative stress during in vitro fertilization. Andrology 2016, 4, 1138–1149. [Google Scholar] [CrossRef]
- Lavrentiadou, S.N.; Sapanidou, V.; Tzekaki, E.E.; Margaritis, I.; Tsantarliotou, M.P. Melatonin protects bovine spermatozoa by reinforcing their antioxidant defenses. Animals 2023, 13, 3219. [Google Scholar] [CrossRef]
- Baumber, J.; Ball, B.A.; Gravance, C.G.; Medina, V.; Davies-Morel, M.C. The effect of reactive oxygen species on equine sperm motility, viability, acrosomal integrity, mitochondrial membrane potential, and membrane lipid peroxidation. J. Androl. 2000, 21, 895–902. [Google Scholar] [CrossRef]
- Gürler, H.; Malama, E.; Heppelmann, M.; Calisici, O.; Leiding, C.; Kastelic, J.P.; Bollwein, H. Effects of cryopreservation on sperm viability, synthesis of reactive oxygen species, and DNA damage of bovine sperm. Theriogenology 2016, 86, 562–571. [Google Scholar] [CrossRef]
- Yánez-Ortiz, I.; Catalán, J.; Rodríguez-Gil, J.E.; Miró, J.; Yeste, M. Advances in sperm cryopreservation in farm animals: Cattle, horse, pig and sheep. Anim. Reprod. Sci. 2022, 246, 106904. [Google Scholar] [CrossRef] [PubMed]
- Hammerstedt, R.H. Maintenance of bioenergetic balance in sperm and prevention of lipid peroxidation: A review of the effect on design of storage preservation systems. Reprod. Fertil. Dev. 1993, 5, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Mammalian fertilization. Physiol. Reprod. 1994, 1, 273–279. [Google Scholar]
- Sapanidou, V.; Lavrentiadou, S.N.; Errico, M.; Panagiotidis, I.; Fletouris, D.; Efraimidis, I.; Zervos, I.; Taitzoglou, I.; Gasparrini, B.; Tsantarliotou, M. The addition of crocin in the freezing medium extender improves post-thaw semen quality. Reprod. Domest. Anim. 2022, 57, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, J.F.; Blanchette, S.; Gagnon, C.; Sirard, M.A. Thiols prevent H2O2-mediated loss of sperm motility in cryopreserved bull semen. Theriogenology 2001, 56, 275–286. [Google Scholar] [CrossRef]
- Gadea, J.; Sellés, E.; Marco, M.A.; Coy, P.; Matás, C.; Romar, R.; Ruiz, S. Decrease in glutathione content in boar sperm after cryopreservation: Effect of the addition of reduced glutathione to the freezing and thawing extenders. Theriogenology 2004, 62, 690–701. [Google Scholar] [CrossRef]
- Aurich, J.E.; Schönherr, U.; Hoppe, H.; Aurich, C. Effects of antioxidants on motility and membrane integrity of chilled-stored stallion semen. Theriogenology 1997, 48, 185–192. [Google Scholar] [CrossRef]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef]
- Gerlach, M.; Ben-Shachar, D.; Riederer, P.; Youdim, M.B.H. Altered brain metabolism of iron as a cause of neurodegenerative diseases? J. Neurochem. 1994, 63, 793–807. [Google Scholar] [CrossRef]
- Tiwari, B.S.; Belenghi, B.; Levine, A. Oxidative stress increased respiration and generation of reactive oxygen species, resulting in ATP depletion, opening of mitochondrial permeability transition, and programmed cell death. Plant Physiol. 2002, 128, 1271–1281. [Google Scholar] [CrossRef]
- Cardoso, S.M.; Pereira, C.; Oliveira, C.R. Mitochondrial function is differentially affected upon oxidative stress. Free Radic. Biol. Med. 1999, 26, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.; Czapski, G.; Cohen, H.; Meyerstein, D. Free radicals induced peptide damage in the presence of transition metal ions: A plausible pathway for biological deleterious processes. Free Radic. Biol. Med. 1994, 17, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Feidantsis, K.; Georgoulis, I.; Giantsis, I.A.; Michaelidis, B. Treatment with ascorbic acid normalizes the aerobic capacity, antioxidant defence, and cell death pathways in thermally stressed Mytilus galloprovincialis. Comp. Biochem. Physiol. B 2021, 255, 110611. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.M.; Pereira, C.; Oliveira, C.R. The protective effect of vitamin E, idebenone and reduced glutathione on free radical mediated injury in rat brain synaptosomes. Biochem. Biophys. Res. Commun. 1998, 246, 703–710. [Google Scholar] [CrossRef]
- Abdolmaleki, A.; Ghayour, M.B.; Behnam-Rassouli, M. Protective effects of acetyl-l-carnitine against serum and glucose deprivation-induced apoptosis in rat adipose-derived mesenchymal stem cells. Cell Tissue Bank. 2020, 21, 655–666. [Google Scholar] [CrossRef]
- Vashisht, A.; Gahlay, G.K. Using miRNAs as diagnostic biomarkers for male infertility: Opportunities and challenges. Mol. Hum. Reprod. 2020, 26, 199–214. [Google Scholar] [CrossRef]
- Hai, E.; Li, B.; Zhang, J.; Zhang, J. Sperm freezing damage: The role of regulated cell death. Cell Death Discov. 2024, 10, 239. [Google Scholar] [CrossRef]
- Min, C.G.; Ma, X.; Wang, Y.C.; Zhong, C.K.; Yuan, C.S.; Zhang, K.Y.; Zhan, C.L.; Hou, S.K.; Wang, X.H.; Wang, J.; et al. The effects of repeated freezing and thawing on bovine sperm morphometry and function. Cryobiology 2024, 115, 104892. [Google Scholar] [CrossRef]
- Söderquist, L.; Rodriguez−Martinez, H.; Janson, L. Post−thaw motility, ATP content and cytochrome C oxidase activity of AI bull spermatozoa in relation to fertility. J. Vet. Med. A 1991, 38, 165–174. [Google Scholar] [CrossRef]
- Pua, H.H.; Guo, J.; Komatsu, M.; He, Y.W. Autophagy is essential for mitochondrial clearance in mature T lymphocytes. J. Immunol. 2009, 182, 4046–4055. [Google Scholar] [CrossRef]
- Cassel, M.; Camargo, M.D.P.; Oliveira De Jesus, L.W.; Borella, M.I. Involution processes of follicular atresia and post-ovulatory complex in a characid fish ovary: A study of apoptosis and autophagy pathways. J. Mol. Histol. 2017, 48, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Feidantsis, K.; Mellidis, K.; Galatou, E.; Sinakos, Z.; Lazou, A. Treatment with crocin improves cardiac dysfunction by normalizing autophagy and inhibiting apoptosis in STZ-induced diabetic cardiomyopathy. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.F. Till death do us part: The marriage of autophagy and apoptosis. Oxidative Med. Cell. Longev. 2018, 2018, 4701275. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, I.M.; Munoz, P.M.; Salido, G.M.; Pena, F.J.; Tapia, J.A. The autophagy-related protein LC3 is processed in stallion spermatozoa during short-and long-term storage and the related stressful conditions. Animals 2016, 10, 1182–1191. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chavas, C.; Sapanidou, V.G.; Feidantsis, K.; Lavrentiadou, S.N.; Mavrogianni, D.; Zarogoulidou, I.; Fletouris, D.J.; Tsantarliotou, M.P. Treatment with Pterostilbene Ameliorates the Antioxidant Status of Bovine Spermatozoa and Modulates Cell Death Pathways. Antioxidants 2024, 13, 1437. https://doi.org/10.3390/antiox13121437
Chavas C, Sapanidou VG, Feidantsis K, Lavrentiadou SN, Mavrogianni D, Zarogoulidou I, Fletouris DJ, Tsantarliotou MP. Treatment with Pterostilbene Ameliorates the Antioxidant Status of Bovine Spermatozoa and Modulates Cell Death Pathways. Antioxidants. 2024; 13(12):1437. https://doi.org/10.3390/antiox13121437
Chicago/Turabian StyleChavas, Christos, Vasiliki G. Sapanidou, Konstantinos Feidantsis, Sophia N. Lavrentiadou, Despoina Mavrogianni, Ioanna Zarogoulidou, Dimitrios J. Fletouris, and Maria P. Tsantarliotou. 2024. "Treatment with Pterostilbene Ameliorates the Antioxidant Status of Bovine Spermatozoa and Modulates Cell Death Pathways" Antioxidants 13, no. 12: 1437. https://doi.org/10.3390/antiox13121437
APA StyleChavas, C., Sapanidou, V. G., Feidantsis, K., Lavrentiadou, S. N., Mavrogianni, D., Zarogoulidou, I., Fletouris, D. J., & Tsantarliotou, M. P. (2024). Treatment with Pterostilbene Ameliorates the Antioxidant Status of Bovine Spermatozoa and Modulates Cell Death Pathways. Antioxidants, 13(12), 1437. https://doi.org/10.3390/antiox13121437