
Redox Homeostasis and Nrf2-Regulated Mechanisms Are Relevant to Male Infertility

 ,
,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
A Brief General Overview of Male Infertility
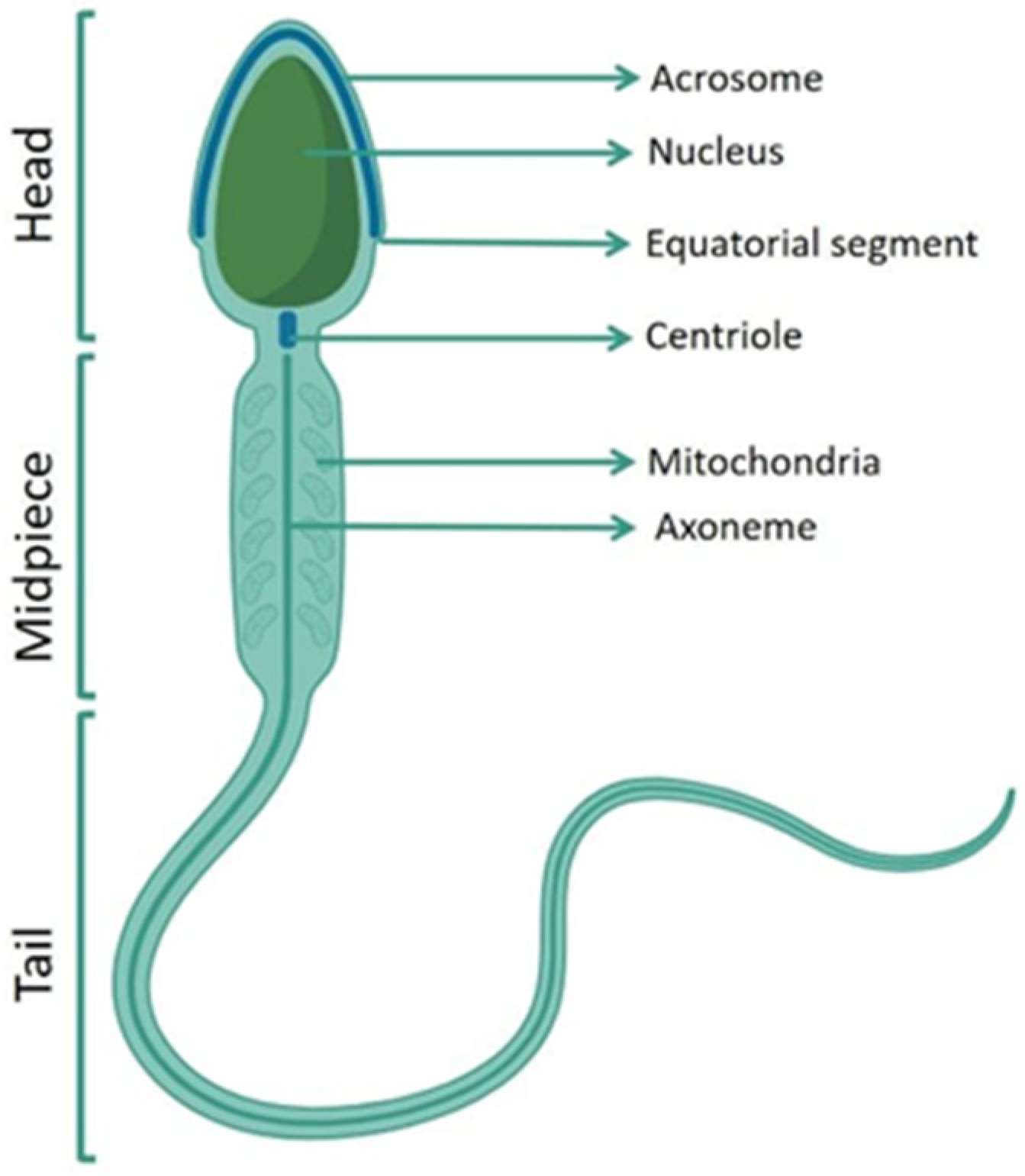
2. Male Infertility and Oxidative Stress
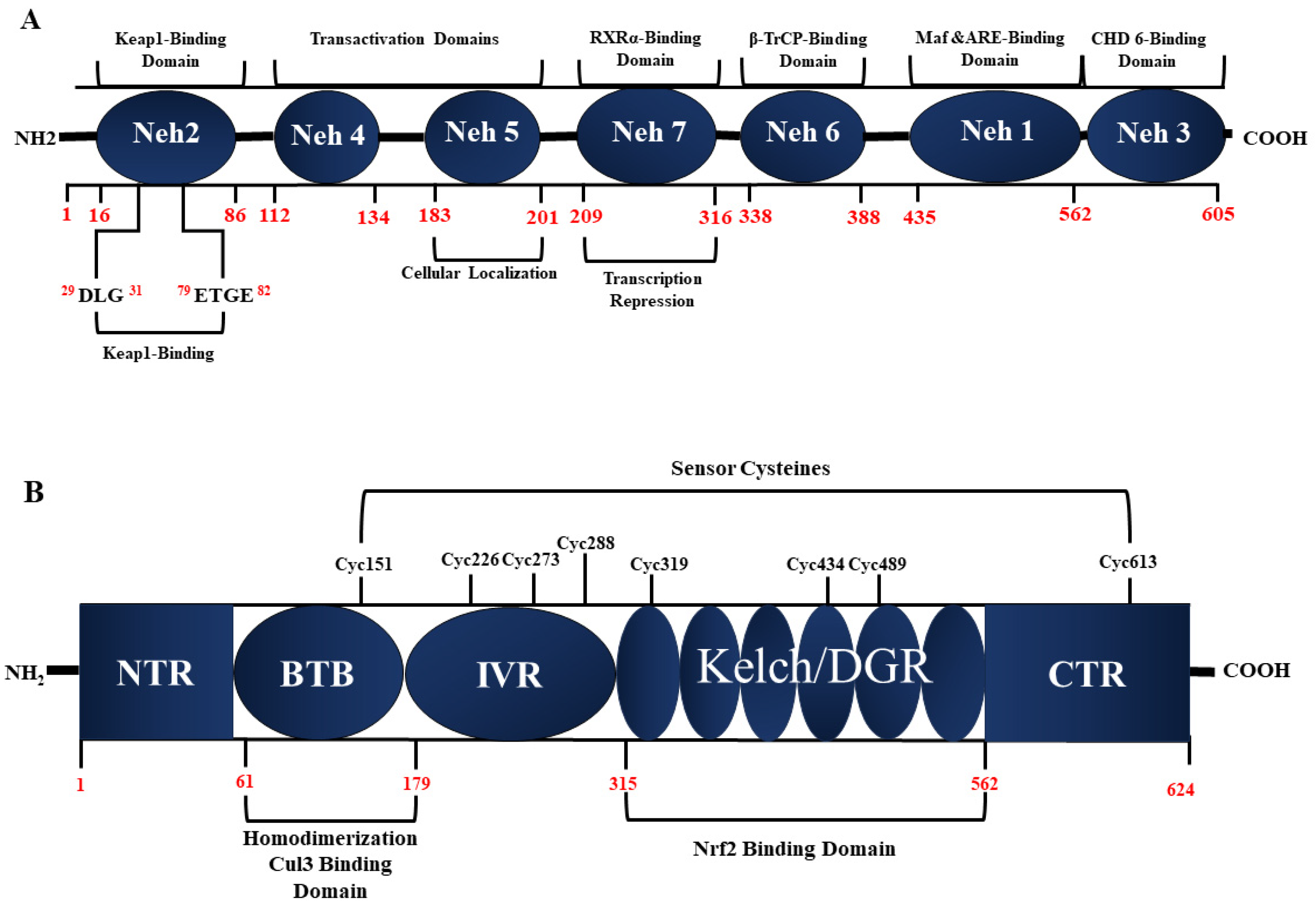
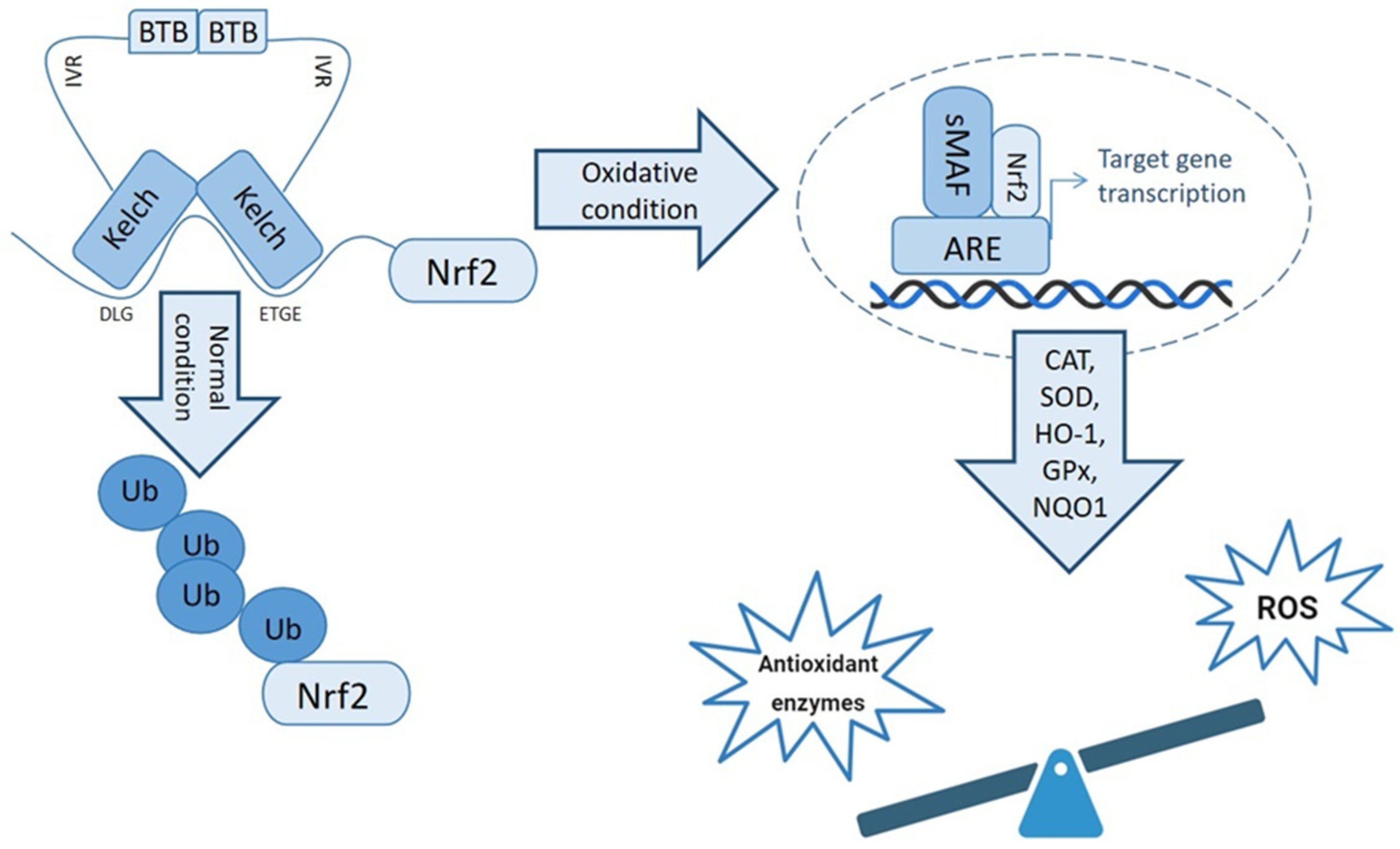
3. Keap1-Nrf2 Antioxidant Pathway
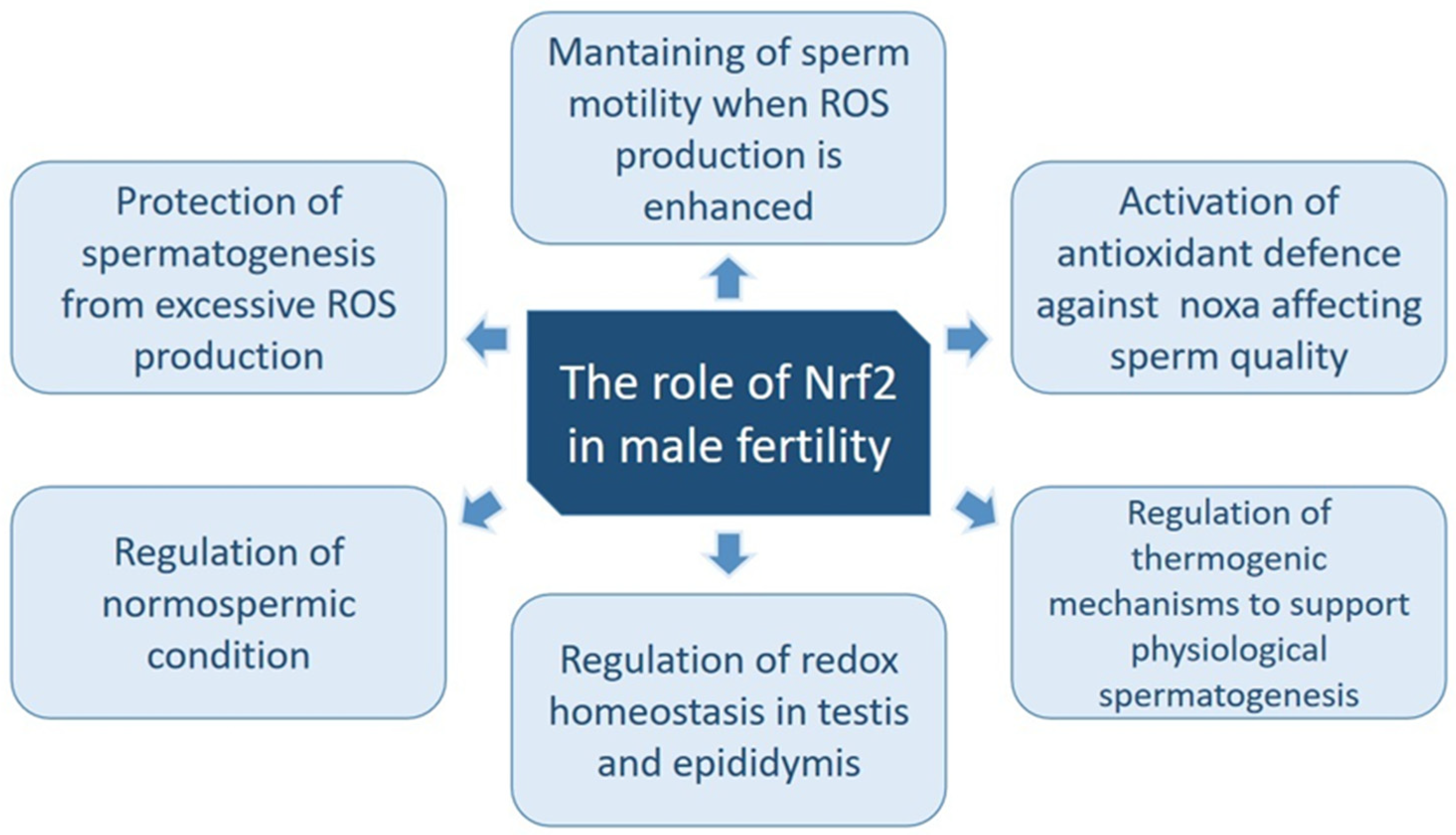
4. Role of Nrf2 in Male Fertility
4.1. Oxidative Stress and Nrf2-Related Gene Regulation
4.2. Effects of Nrf2 Gene Deletion on Spermatogenesis
4.3. Thermoregulation Mechanisms of Nrf2 in Spermatogenesis
4.4. Role of Nrf2 in Long-Term Storage of Sperm
5. Compounds Targeting Nrf2 and Influencing Male Fertility
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; WHO Press: Geneva, Switzerland, 2021. [Google Scholar]
- Leslie, S.W.; Soon-Sutton, T.L.; Khan, M.A.B. Male infertility. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Drevet, J.R.; Aitken, R.J. Oxidative Damage to Sperm DNA: Attack and Defense. Adv. Exp. Med. Biol. 2019, 1166, 107–117. [Google Scholar] [PubMed]
- Agarwal, A.; Majzoub, A.; Baskaran, S.; Panner Selvam, M.K.; Cho, C.L.; Henkel, R.; Finelli, R.; Leisegang, K.; Sengupta, P.; Barbarosie, C.; et al. Sperm DNA Fragmentation: A New Guideline for Clinicians. World J. Mens Health 2020, 38, 412–471. [Google Scholar] [CrossRef]
- Han, P.; Wang, X.; Zhou, T.; Cheng, J.; Wang, C.; Sun, F.; Zhao, X. Inhibition of ferroptosis attenuates oligospermia in male Nrf2 knockout mice. Free Radic. Biol. Med. 2022, 193, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Ghareghomi, S.; Habibi-Rezaei, M.; Arese, M.; Saso, L.; Moosavi-Movahedi, A.A. Nrf2 modulation in breast cancer. Biomedicines 2022, 10, 2668. [Google Scholar] [CrossRef]
- Krausz, C. Male infertility: Pathogenesis and clinical diagnosis. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 271–285. [Google Scholar] [CrossRef]
- Corsini, C.; Boeri, L.; Candela, L.; Pozzi, E.; Belladelli, F.; Capogrosso, P.; Fallara, G.; Schifano, N.; Cignoli, D.; Ventimiglia, E.; et al. Is There a Relevant Clinical Impact in Differentiating Idiopathic versus Unexplained Male Infertility? World J Mens Health 2023, 41, 354–362. [Google Scholar] [CrossRef]
- Neto, F.T.; Bach, P.V.; Najari, B.B.; Li, P.S.; Goldstein, M. Spermatogenesis in humans and its affecting factors. Semin. Cell Dev. Biol. 2016, 59, 10–26. [Google Scholar] [CrossRef]
- O’Donnell, L.; Smith, L.B.; Rebourcet, D. Sertoli cells as key drivers of testis function. Semin. Cell Dev. Biol. 2022, 121, 2–9. [Google Scholar] [CrossRef]
- Björndahl, L.; Brown, J.K. The sixth edition of the WHO Laboratory Manual for the Examination and Processing of Human Semen: Ensuring quality and standardization in basic examination of human ejaculates. Fertil. Steril. 2022, 117, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Dave, P.; Farber, N.; Vij, S. Conventional semen analysis and advanced sperm function tests in diagnosis and management of varicocele. Andrologia 2021, 53, e13629. [Google Scholar] [CrossRef] [PubMed]
- Llavanera, M.; Delgado-Bermúdez, A.; Ribas-Maynou, J.; Salas-Huetos, A.; Yeste, M. A systematic review identifying fertility biomarkers in semen: A clinical approach through Omics to diagnose male infertility. Fertil. Steril. 2022, 118, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. On the history of oxidative stress: Concept and some aspects of current development. Curr. Opin. Toxicol. 2018, 7, 122–126. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Clarendon Press: Oxford, UK, 2015. [Google Scholar]
- Mannucci, A.; Argento, F.R.; Fini, E.; Coccia, M.E.; Taddei, N.; Becatti, M.; Fiorillo, C. The Impact of Oxidative Stress in Male Infertility. Front. Mol. Biosci. 2022, 8, 799294. [Google Scholar] [CrossRef] [PubMed]
- Moretti, E.; Signorini, C.; Corsaro, R.; Giamalidi, M.; Collodel, G. Human Sperm as an In Vitro Model to Assess the Efficacy of Antioxidant Supplements during Sperm Handling: A Narrative Review. Antioxidants 2023, 12, 1098. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary evidence on the physiological role of reactive oxygen species in human sperm function. J. Assist. Reprod. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef]
- Takei, G.L.; Tourzani, D.A.; Paudel, B.; Visconti, P.E. Activation of cAMP-dependent phosphorylation pathways is independent of ROS production during mouse sperm capacitation. Mol. Reprod. Dev. 2021, 8, 544–557. [Google Scholar] [CrossRef]
- Barati, E.; Nikzad, H.; Karimian, M. Oxidative stress and male infertility: Current knowledge of pathophysiology and role of antioxidant therapy in disease management. Cell. Mol. Life Sci. 2020, 77, 93–113. [Google Scholar] [CrossRef]
- Tvrdá, E.; Kňažická, Z.; Bárdos, L.; Massányi, P.; Lukáč, N. Impact of oxidative stress on male fertility—A review. Acta Vet. Hung. 2011, 59, 465–484. [Google Scholar] [CrossRef]
- O’Flaherty, C. Protective role of peroxiredoxins against reactive oxygen species in neonatal rat testicular gonocytes. Antioxidants 2019, 9, 32. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, H.W. Seminal leukocytes: Passengers, terrorists or good samaritans? Hum. Reprod. 1995, 10, 1736–1739. [Google Scholar] [CrossRef]
- Alvarez, J.G. Nurture vs nature: How can we optimize sperm quality? J. Androl. 2003, 24, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ni, M.; Xing, S.; Yu, Y.; Zhou, Y.; Yang, S.; Li, H.; Zhu, R.; Han, M. Reactive Oxygen Species Secreted by Leukocytes in Semen Induce Self-Expression of Interleukin-6 and Affect Sperm Quality. Am. J. Mens Health 2020, 14, 1557988320970053. [Google Scholar] [CrossRef]
- Micheli, L.; Cerretani, D.; Collodel, G.; Menchiari, A.; Moltoni, L.; Fiaschi, A.I.; Moretti, E. Evaluation of enzymatic and non-enzymatic antioxidants in seminal plasma of men with genitourinary infections, varicocele, and idiopathic infertility. Andrology 2016, 4, 456–464. [Google Scholar] [CrossRef]
- Henkel, R.R. Leukocytes and oxidative stress: Dilemma for sperm function and male fertility. Asian J. Androl. 2011, 13, 43. [Google Scholar] [CrossRef]
- Castleton, P.E.; Deluao, J.C.; Sharkey, D.J.; McPherson, N.O. Measuring reactive oxygen species in semen for male preconception care: A scientist perspective. Antioxidants 2022, 11, 264. [Google Scholar] [CrossRef] [PubMed]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Moretti, E.; Cerretani, D.; Noto, D.; Signorini, C.; Iacoponi, F.; Collodel, G. Relationship between semen IL-6, IL-33 and malondialdehyde generation in human seminal plasma and spermatozoa. Reprod. Sci. 2021, 28, 2136–2143. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Collodel, G. Role of isoprostanes in human male infertility. Syst. Biol. Reprod. Med. 2020, 66, 291–299. [Google Scholar] [CrossRef]
- Naderi Noreini, S.; Malmir, M.; Ghafarizadeh, A.; Faraji, T.; Bayat, R. Protective effect of L-carnitine on apoptosis, DNA fragmentation, membrane integrity and Lipid peroxidation of spermatozoa in the asthenoteratospermic men. Andrologia 2021, 53, e13932. [Google Scholar] [CrossRef]
- Wathes, D.C.; Abayasekara, D.R.E.; Aitken, R.J. Polyunsaturated fatty acids in male and female reproduction. Biol. Reprod. 2007, 77, 190–201. [Google Scholar] [CrossRef]
- delBarco-Trillo, J.; Roldan, E.R. Effects of metabolic rate and sperm competition on the fatty-acid composition of mammalian sperm. J. Evol. Biol. 2014, 27, 55–62. [Google Scholar] [CrossRef]
- Crisóstomo, L.; Videira, R.A.; Jarak, I.; Starčević, K.; Mašek, T.; Rato, L.; Raposo, J.F.; Batterham, R.L.; Oliveira, P.F.; Alves, M.G. Diet during early life defines testicular lipid content and sperm quality in adulthood. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E1061–E1073. [Google Scholar] [CrossRef]
- Suliga, E.; Głuszek, S. The relationship between diet, energy balance and fertility in men. Int. J. Vitam. Nutr. Res. 2020, 90, 514–5262019. [Google Scholar] [CrossRef]
- Kljajic, M.; Hammadeh, M.E.; Wagenpfeil, G.; Baus, S.; Sklavounos, P.; Solomayer, E.F.; Kasoha, M. Impact of the vegan diet on sperm quality and sperm oxidative stress values: A preliminary study. J. Hum. Reprod. Sci. 2021, 14, 365. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Diet and male fertility: The impact of nutrients and antioxidants on sperm energetic metabolism. Int. J. Mol. Sci. 2022, 23, 2542. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P.; Slama, P.; Roychoudhury, S. Oxidative stress, testicular inflammatory pathways, and male reproduction. Int. J. Mol. Sci. 2021, 22, 10043. [Google Scholar] [CrossRef] [PubMed]
- Takalani, N.B.; Monageng, E.M.; Mohlala, K.; Monsees, T.K.; Henkel, R.; Opuwari, C.S. Role of oxidative stress in male infertility. Reprod. Fertil. 2023, 4, e230024. [Google Scholar] [CrossRef] [PubMed]
- Arya, D.; Balasinor, N.; Singh, D. Varicocoele-associated male infertility: Cellular and molecular perspectives of pathophysiology. Andrology 2022, 10, 1463–1483. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E. Revisiting the impact of varicocele and its treatments on male fertility. Reprod. Biomed. Online 2022, 45, 1061–1063. [Google Scholar] [CrossRef]
- Bisconti, M.; Simon, J.F.; Grassi, S.; Leroy, B.; Martinet, B.; Arcolia, V.; Isachenko, V.; Hennebert, E. Influence of risk factors for male infertility on sperm protein composition. Int. J. Mol. Sci. 2021, 22, 13164. [Google Scholar] [CrossRef]
- Minas, A.; Fernandes, A.C.C.; Maciel Júnior, V.L.; Adami, L.; Intasqui, P.; Bertolla, R.P. Influence of physical activity on male fertility. Andrologia 2022, 54, e14433. [Google Scholar] [CrossRef]
- Kaur, P.; Rai, U.; Singh, R. Genotoxic Risks to Male Reproductive Health from Radiofrequency Radiation. Cells 2023, 12, 59456. [Google Scholar] [CrossRef]
- Paulis, G. Inflammatory mechanisms and oxidative stress in prostatitis: The possible role of antioxidant therapy. Res. Rep. Urol. 2018, 10, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Rana, M.; Qiu, E.; AlBunni, H.; Bui, A.D.; Henkel, R. Role of oxidative stress, infection and inflammation in male infertility. Andrologia 2018, 50, e13126. [Google Scholar] [CrossRef] [PubMed]
- Hærvig, K.K.; Kierkegaard, L.; Lund, R.; Bruunsgaard, H.; Osler, M.; Schmidt, L. Is male factor infertility associated with midlife low-grade inflammation? A population based study. Hum. Fertil. 2018, 21, 146–154. [Google Scholar] [CrossRef]
- Motrich, R.D.; Salazar, F.C.; Breser, M.L.; Mackern-Oberti, J.P.; Godoy, G.J.; Olivera, C.; Paira, D.A.; Rivero, V.E. Implications of prostate inflammation on male fertility. Andrologia 2018, 50, e13093. [Google Scholar] [CrossRef] [PubMed]
- Leisegang, K.; Sengupta, P.; Agarwal, A.; Henkel, R. Obesity and male infertility: Mechanisms and management. Andrologia 2021, 53, e13617. [Google Scholar] [CrossRef]
- Collodel, G.; Signorini, C.; Nerucci, F.; Gambera, L.; Iacoponi, F.; Moretti, E. Semen biochemical components in varicocele, leukocytospermia, and idiopathic infertility. Reprod. Sci. 2021, 28, 91–101. [Google Scholar] [CrossRef]
- Koppenol, W.H.; Hider, R. Iron and redox cycling. Do’s and don’ts. Free Radic. Biol. Med. 2019, 133, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Seiwert, N.; Adam, J.; Steinberg, P.; Wirtz, S.; Schwerdtle, T.; Adams-Quack, P.; Hövelmeyer, N.; Kaina, B.; Foersch, S.; Fahrer, J. Chronic intestinal inflammation drives colorectal tumor formation triggered by dietary heme iron in vivo. Arch. Toxicol. 2021, 95, 2507–2522. [Google Scholar] [CrossRef] [PubMed]
- Marques, O.; Weiss, G.; Muckenthaler, M.U. The role of iron in chronic inflammatory diseases: From mechanisms to treatment options in anemia of inflammation. Blood 2022, 140, 2011–2023. [Google Scholar] [CrossRef]
- Jing, T.; Wang, P.; Liu, Y.; Zhao, J.; Niu, X.; Wang, X. Pathological changes in Sertoli cells and dysregulation of divalent metal transporter 1 with iron responsive element in the testes of idiopathic azoospermia patients. Andrologia 2018, 50, e12878. [Google Scholar] [CrossRef]
- Ammar, O.; Houas, Z.; Mehdi, M. The association between iron, calcium, and oxidative stress in seminal plasma and sperm quality. Environ. Sci. Pollut. Res. Int. 2019, 26, 14097–14105. [Google Scholar] [CrossRef]
- Li, L.; Hao, Y.; Zhao, Y.; Wang, H.; Zhao, X.; Jiang, Y.; Gao, F. Ferroptosis is associated with oxygen-glucose deprivation/reoxygenation-induced Sertoli cell death. Int. J. Mol. Med. 2018, 41, 3051–3062. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, H.; Cui, J.G.; Wang, J.X.; Chen, M.S.; Wang, H.R.; Li, X.N.; Li, J.L. Ferroptosis is critical for phthalates driving the blood-testis barrier dysfunction via targeting transferrin receptor. Redox Biol. 2023, 59, 102584. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Noto, D.; Micheli, L.; Ponchia, R.; Collodel, G. Fatty Acid Oxidation and Pro-Resolving Lipid Mediators Are Related to Male Infertility. Antioxidants 2022, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Chiang, N.; Serhan, C.N. Specialized pro-resolving mediator network: An update on production and actions. Essays Biochem. 2020, 64, 443–462. [Google Scholar] [PubMed]
- Lo, S.C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1: Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J. 2006, 25, 3605–3617. [Google Scholar] [CrossRef] [PubMed]
- Nam, L.B.; Keum, Y.S. Binding partners of NRF2: Functions and regulatory mechanisms. Arch. Biochem. Biophys. 2019, 678, 108184. [Google Scholar] [CrossRef] [PubMed]
- Namani, A.; Li, Y.; Wang, X.J.; Tang, X. Modulation of NRF2 signaling pathway by nuclear receptors: Implications for cancer. Biochim. Biophys. Acta. 2014, 1843, 1875–1885. [Google Scholar] [CrossRef] [PubMed]
- Krajka-Kuźniak, V.; Paluszczak, J.; Baer-Dubowska, W. The Nrf2-ARE signaling pathway: An update on its regulation and possible role in cancer prevention and treatment. Pharmacol. Rep. 2017, 69, 393–402. [Google Scholar] [CrossRef]
- Keum, Y.S.; Choi, B.Y. Molecular and chemical regulation of the Keap1-Nrf2 signaling pathway. Molecules 2014, 19, 10074–10089. [Google Scholar] [CrossRef]
- Xiang, M.; Namani, A.; Wu, S.; Wang, X. Nrf2: Bane or blessing in cancer? J. Cancer Res. Clin. Oncol. 2014, 140, 1251–1259. [Google Scholar] [CrossRef]
- Bai, X.; Chen, Y.; Hou, X.; Huang, M.; Jin, J. Emerging role of NRF2 in chemoresistance by regulating drug-metabolizing enzymes and efflux transporters. Drug Metab. Rev. 2016, 48, 541–567. [Google Scholar] [CrossRef]
- Wei, S.; Pei, Y.; Wang, Y.; Guan, H.; Huang, Y.; Xing, T.; Johnson, R.W.; Wang, D. Role of human Keap1 S53 and S293 residues in modulating the binding of Keap1 to Nrf2. Biochimie 2019, 158, 73–81. [Google Scholar] [CrossRef]
- Kang, M.I.; Kobayashi, A.; Wakabayashi, N.; Kim, S.G.; Yamamoto, M. Scaffolding of Keap1 to the actin cytoskeleton controls the function of Nrf2 as key regulator of cytoprotective phase 2 genes. Proc. Natl. Acad. Sci. USA 2004, 101, 2046–2051. [Google Scholar] [CrossRef]
- Canning, P.; Sorrell, F.J.; Bullock, A.N. Structural basis of Keap1 interactions with Nrf2. Free Radic. Biol. Med. 2015, 88, 101–107. [Google Scholar] [CrossRef]
- Furukawa, M.; Xiong, Y. BTB protein Keap1 targets antioxidant transcription factor Nrf2 for ubiquitination by the Cullin 3-Roc1 ligase. Mol. Cell. Biol. 2005, 25, 162–171. [Google Scholar] [CrossRef]
- Velichkova, M.; Hasson, T. Keap1 regulates the oxidation-sensitive shuttling of Nrf2 into and out of the nucleus via a Crm1-dependent nuclear export mechanism. Mol. Cell. Biol. 2005, 25, 4501–4513. [Google Scholar] [CrossRef]
- Levonen, A.L.; Landar, A.; Ramachandran, A.; Ceaser, E.K.; Dickinson, D.A.; Zanoni, G.; Morrow, J.D.; Darley-Usmar, V.M. Cellular mechanisms of redox cell signalling: Role of cysteine modification in controlling antioxidant defences in response to electrophilic lipid oxidation products. Biochem. J. 2004, 378, 373–382. [Google Scholar] [CrossRef]
- Zhang, D.D.; Hannink, M. Distinct cysteine residues in Keap1 are required for Keap1-dependent ubiquitination of Nrf2 and for stabilization of Nrf2 by chemopreventive agents and oxidative stress. Mol. Cell. Biol. 2003, 23, 8137–8151. [Google Scholar] [CrossRef]
- Fuse, Y.; Kobayashi, M. Conservation of the Keap1-Nrf2 system: An evolutionary journey through stressful space and time. Molecules 2017, 22, 436. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, Y. Molecular basis of the Keap1–Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, H.; Xu, M.; Wang, X.; Wang, C.; Lian, Y.; Mehmood, A.; Dai, H. Stevia residue extract ameliorates oxidative stress in d-galactose-induced aging mice via Akt/Nrf2/HO-1 pathway. J. Funct. Foods. 2019, 52, 587–595. [Google Scholar] [CrossRef]
- Ghareghomi, S.; Rahban, M.; Moosavi-Movahedi, Z.; Habibi-Rezaei, M.; Saso, L.; Moosavi-Movahedi, A.A. The potential role of curcumin in modulating the master antioxidant pathway in diabetic hypoxia-induced complications. Molecules 2021, 26, 7658. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, B.N.; Lawson, G.; Chan, J.Y.; Banuelos, J.; Cortés, M.M.; Hoang, Y.D.; Ortiz, L.; Rau, B.A.; Luderer, U. Knockout of the transcription factor NRF2 disrupts spermatogenesis in an age-dependent manner. Free Radic. Biol. Med. 2010, 49, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Rotimi, D.E.; Ojo, O.A.; Olaolu, T.D.; Adeyemi, O.S. Exploring Nrf2 as a therapeutic target in testicular dysfunction. Cell Tissue Res. 2022, 390, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Mai, Z.; Zhou, Y.; Gao, X.; Yu, B. Low NRF2 mRNA expression in spermatozoa from men with low sperm motility. Tohoku J. Exp. Med. 2012, 228, 259–266. [Google Scholar] [CrossRef]
- Wajda, A.; Łapczuk, J.; Grabowska, M.; Słojewski, M.; Laszczyńska, M.; Urasińska, E.; Droździk, M. Nuclear factor E2-related factor-2 (Nrf2) expression and regulation in male reproductive tract. Pharmacol. Rep. 2016, 68, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lin, H.; Yang, L.; Chen, K.; Luo, H.; Liu, J.; Gao, X.; Xia, X.; Huang, Z. Genetic variation in the Nrf2 promoter associates with defective spermatogenesis in humans. J. Mol. Med. 2012, 90, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Fernandez, J.; Alférez, M.J.M.; López-Aliaga, I.; Diaz-Castro, J. Protective effects of fermented goat milk on genomic stability, oxidative stress and inflammatory signalling in testis during anaemia recovery. Sci. Rep. 2019, 9, 2232124. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Bai, Y.; Zhang, Z.; Xin, Y.; Cai, L. Protection by sulforaphane from type 1 diabetes-induced testicular apoptosis is associated with the up-regulation of Nrf2 expression and function. Toxicol. Appl. Pharmacol. 2014, 279, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Aydos, O.S.; Yukselten, Y.; Aydos, D.; Sunguroglu, A.; Aydos, K. Relationship between functional Nrf2 gene promoter polymorphism and sperm DNA damage in male infertility. Syst. Biol. Reprod. Med. 2021, 67, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Song, W.; Xu, D.; Chen, X.; Li, X.; Zhao, Y. Autophagy induced by ROS aggravates testis oxidative damage in diabetes via breaking the feedforward loop linking p62 and Nrf2. Oxid. Med. Cell. Longev. 2020, 2020, 7156579. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhao, C.; Guo, H.; Liu, T.; Li, Y.; Qi, Y.; Deussing, J.M.; Zhang, Y.; Tan, J.; Han, H.; et al. Obesity induces male mice infertility via oxidative stress, apoptosis, and glycolysis. Reproduction 2023, 166, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Lu, S.; Ou, B.; Feng, J.; Wang, Z.; Li, H.; Lu, X.; Ma, Y. PACAP ameliorates the fertility of obese mice through PAC1/PKA/ERK/Nrf2 signal axis. J. Endocrinol. 2021, 248, 337–354. [Google Scholar] [CrossRef]
- Agarwal, A.; Maldonado Rosas, I.; Anagnostopoulou, C.; Cannarella, R.; Boitrelle, F.; Munoz, L.V.; Finelli, R.; Durairajanayagam, D.; Henkel, R.; Saleh, R. Oxidative stress and assisted reproduction: A comprehensive review of its pathophysiological role and strategies for optimizing embryo culture environment. Antioxidants 2022, 11, 477. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, Z. Obesity, a serious etiologic factor for male subfertility in modern society. Reproduction 2017, 154, R123–R131. [Google Scholar] [CrossRef]
- Gan, M.; Jing, Y.; Xie, Z.; Ma, J.; Chen, L.; Zhang, S.; Zhao, Y.; Niu, L.; Wang, Y.; Li, X.; et al. Potential Function of Testicular MicroRNAs in Heat-Stress-Induced Spermatogenesis Disorders. Int. J. Mol. Sci. 2023, 24, 8809. [Google Scholar] [CrossRef] [PubMed]
- Aldahhan, R.A.; Stanton, P.G. Heat stress response of somatic cells in the testis. Mol. Cell. Endocrinol. 2021, 527, 111216. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cao, Y.; Wang, F.; Li, C. Scrotal heat induced the Nrf2-driven antioxidant response during oxidative stress and apoptosis in the mouse testis. Acta Histochem. 2014, 116, 883–890. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Sun, J.; Yang, D.; He, W.; Wang, J.; Qin, D.; Zhang, H.; Cai, H.; Liu, Y.; Li, N.; et al. Nrf2 activation mediates the protection of mouse Sertoli Cells damage under acute heat stress conditions. Theriogenology 2022, 177, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Sadeghiani, G.; Khanehzad, M.; Sadighi Gilani, M.A.; Amidi, F.; Malekzadeh, M.; Rastegar, T. Evaluation of Nrf2/ARE signaling pathway in the presence of pentoxifylline as a cryoprotectant in mouse spermatogonial stem cells. Biopreserv. Biobank. 2023, 21, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, N.; Nasr, N.H.; Abu Elnaga, N.A.; Abou-Hashim, F.; Kamel, S.; Warda, M.; Dessouki, S.M.; AbdRabou, M.A.; Mehaisen, G.M.K. Molecular and physiochemical evaluation of buck semen cryopreserved with antioxidants. Reprod. Domest. Anim. 2023, 58, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Jannatifar, R.; Asa, E.; Sahraei, S.S.; Verdi, A.; Piroozmanesh, H. N-acetyl-l-cysteine and alpha lipoic acid are protective supplement on human sperm parameters in cryopreservation of asthenoteratozoospermia patients. Andrologia 2022, 54, e14612. [Google Scholar] [CrossRef]
- Clark, I.; Brougham, M.F.H.; Spears, N.; Mitchell, R.T. The impact of vincristine on testicular development and function in childhood cancer. Hum. Reprod. Update 2023, 29, 233–245. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Abak, A.; Seify, M.; Mohaqiq, M.; Keshmir, F.; Taheri, M.; Ayatollahi, S.A. Effects of chemotherapeutic agents on male germ cells and possible ameliorating impact of antioxidants. Biomed. Pharmacother. 2021, 142, 112040. [Google Scholar] [CrossRef]
- Zakhem, G.A.; Motosko, C.C.; Mu, E.W.; Ho, R.S. Infertility and teratogenicity after paternal exposure to systemic dermatologic medications: A systematic review. J. Am. Acad. Dermatol. 2019, 80, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Jones, R.J.; Brodsky, R.A. Cyclophosphamide and cancer: Golden anniversary. Nat. Rev. Clin. Oncol. 2009, 6, 638–647. [Google Scholar] [CrossRef]
- Maremanda, K.P.; Khan, S.; Jena, G. Zinc protects cyclophosphamide-induced testicular damage in rat: Involvement of metallothionein, tesmin and Nrf2. Biochem. Biophys. Res. Commun. 2014, 445, 591–596. [Google Scholar] [CrossRef]
- Robles-Almazan, M.; Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Rodriguez-Garcia, C.; Quiles, J.L.; Ramirez-Tortosa, M. Hydroxytyrosol: Bioavailability, toxicity, and clinical applications. Food Res. Int. 2018, 105, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Peyrol, J.; Riva, C.; Amiot, M.J. Hydroxytyrosol in the prevention of the metabolic syndrome and related disorders. Nutrients 2017, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morató, J.; Xicota, L.; Fitó, M.; Farré, M.; Dierssen, M.; de la Torre, R. Potential role of olive oil phenolic compounds in the prevention of neurodegenerative diseases. Molecules 2015, 20, 4655–4680. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Trovato Salinaro, A.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Crea, R.; Cordaro, M.; Cuzzocrea, S.; et al. Hidrox® counteracts cyclophosphamide-induced male infertility through NRF2 pathways in a mouse model. Antioxidants 2021, 10, 778. [Google Scholar] [CrossRef] [PubMed]
- Bae, W.J.; Ha, U.S.; Choi, J.B.; Kim, K.S.; Kim, S.J.; Cho, H.J.; Hong, S.H.; Lee, J.Y.; Wang, Z.; Hwang, S.Y.; et al. Protective effect of decursin extracted from Angelica gigas in male infertility via Nrf2/HO-1 signaling pathway. Oxid. Med. Cell Longev. 2016, 2016, 5901098. [Google Scholar] [CrossRef]
- Arkali, G.; Aksakal, M.; Kaya, Ş.Ö. Protective effects of carvacrol against diabetes-induced reproductive damage in male rats: Modulation of Nrf2/HO-1 signalling pathway and inhibition of Nf-kB-mediated testicular apoptosis and inflammation. Andrologia 2021, 53, e13899. [Google Scholar] [CrossRef]
- Shi, X.; Fu, L. Piceatannol inhibits oxidative stress through modification of Nrf2-signaling pathway in testes and attenuates spermatogenesis and steroidogenesis in rats exposed to cadmium during adulthood. Drug Des. Dev. Ther. 2019, 13, 2811–2824. [Google Scholar] [CrossRef]
- Hu, W.H.; Dai, D.K.; Zheng, B.Z.; Duan, R.; Dong, T.T.; Qin, Q.W.; Tsim, K.W. Piceatannol, a natural analog of resveratrol, exerts anti-angiogenic efficiencies by blockage of vascular endothelial growth factor binding to its receptor. Molecules 2020, 25, 3769. [Google Scholar] [CrossRef]
- Rajput, S.A.; Liang, S.J.; Wang, X.Q.; Yan, H.C. Lycopene protects intestinal epithelium from deoxynivalenol-induced oxidative damage via regulating Keap1/Nrf2 Signaling. Antioxidants 2021, 10, 1493. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.Z.; Shen, Y.; Lin, J.; Wang, H.R.; Talukder, M.; Li, J.L. Lycopene prevents DEHP-induced Leydig cell damage with the Nrf2 antioxidant signaling pathway in mice. J. Agric. Food Chem. 2019, 68, 2031–2040. [Google Scholar] [CrossRef]
- Guo, Y.; Sun, J.; Li, T.; Zhang, Q.; Bu, S.; Wang, Q.; Lai, D. Melatonin ameliorates restraint stress-induced oxidative stress and apoptosis in testicular cells via NF-κB/iNOS and Nrf2/HO-1 signaling pathway. Sci. Rep. 2017, 7, 9599. [Google Scholar] [CrossRef]
- Dong, Y.; Zhao, J.; Zhu, Q.; Liu, H.; Wang, J.; Lu, W. Melatonin inhibits the apoptosis of rooster Leydig cells by suppressing oxidative stress via AKT-Nrf2 pathway activation. Free Radic. Biol. Med. 2020, 160, 1–12. [Google Scholar] [CrossRef]
- Bartolini, D.; Arato, I.; Mancuso, F.; Giustarini, D.; Bellucci, C.; Vacca, C.; Aglietti, M.C.; Stabile, A.M.; Rossi, R.; Cruciani, G.; et al. Melatonin modulates Nrf2 activity to protect porcine pre-pubertal Sertoli cells from the abnormal H2O2 generation and reductive stress effects of cadmium. J. Pineal Res. 2022, 73, e12806. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, J.; Zhu, Z.; Bao, X.; Zhang, M.; Ren, C.; Zhang, Q. Aucubin, a natural iridoid glucoside, attenuates oxidative stress-induced testis injury by inhibiting JNK and CHOP activation via Nrf2 up-regulation. Phytomedicine 2019, 64, 153057. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wu, X.; Liu, Y.; Wang, X.; Ling, X.; Ge, H.; Zhang, J. Curcumin improves asthenozoospermia by inhibiting reactive oxygen species reproduction through nuclear factor erythroid 2-related factor 2 activation. Andrologia 2020, 52, e13491. [Google Scholar] [CrossRef] [PubMed]
- Jannatifar, R.; Parivar, K.; Hayati Roodbari, N.; Nasr-Esfahani, M.H. The effect of N-acetyl-cysteine on NRF2 antioxidant gene expression in asthenoteratozoospermia men: A clinical trial study. Int. J. Fertil. Steril. 2020, 14, 171. [Google Scholar] [PubMed]
- Fallahi, S.; Rajaei, M.; Hesam, M.J.; Koolivand, M.; Malekzadeh, K. The effect of Phoenix dactylifera pollen on the expression of NRF2, SOD2, CAT, and GPX4 genes, and sperm parameters of fertile and infertile men: A controlled clinical trial. Int. J. Reprod. Biomed. 2021, 19, 545. [Google Scholar] [CrossRef] [PubMed]
- El-Kholy, W.M.; Soliman, T.N.; Darwish, A.M.G. Evaluation of date palm pollen (Phoenix dactylifera L.) encapsulation, impact on the nutritional and functional properties of fortified yoghurt. PLoS ONE 2019, 14, e0222789. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Signorini, C.; Saso, L.; Ghareghomi, S.; Telkoparan-Akillilar, P.; Collodel, G.; Moretti, E. Redox Homeostasis and Nrf2-Regulated Mechanisms Are Relevant to Male Infertility. Antioxidants 2024, 13, 193. https://doi.org/10.3390/antiox13020193
Signorini C, Saso L, Ghareghomi S, Telkoparan-Akillilar P, Collodel G, Moretti E. Redox Homeostasis and Nrf2-Regulated Mechanisms Are Relevant to Male Infertility. Antioxidants. 2024; 13(2):193. https://doi.org/10.3390/antiox13020193
Chicago/Turabian StyleSignorini, Cinzia, Luciano Saso, Somayyeh Ghareghomi, Pelin Telkoparan-Akillilar, Giulia Collodel, and Elena Moretti. 2024. "Redox Homeostasis and Nrf2-Regulated Mechanisms Are Relevant to Male Infertility" Antioxidants 13, no. 2: 193. https://doi.org/10.3390/antiox13020193
APA StyleSignorini, C., Saso, L., Ghareghomi, S., Telkoparan-Akillilar, P., Collodel, G., & Moretti, E. (2024). Redox Homeostasis and Nrf2-Regulated Mechanisms Are Relevant to Male Infertility. Antioxidants, 13(2), 193. https://doi.org/10.3390/antiox13020193