

Therapeutic Effects of Hinokitiol through Regulating the SIRT1/NOX4 against Ligature-Induced Experimental Periodontitis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
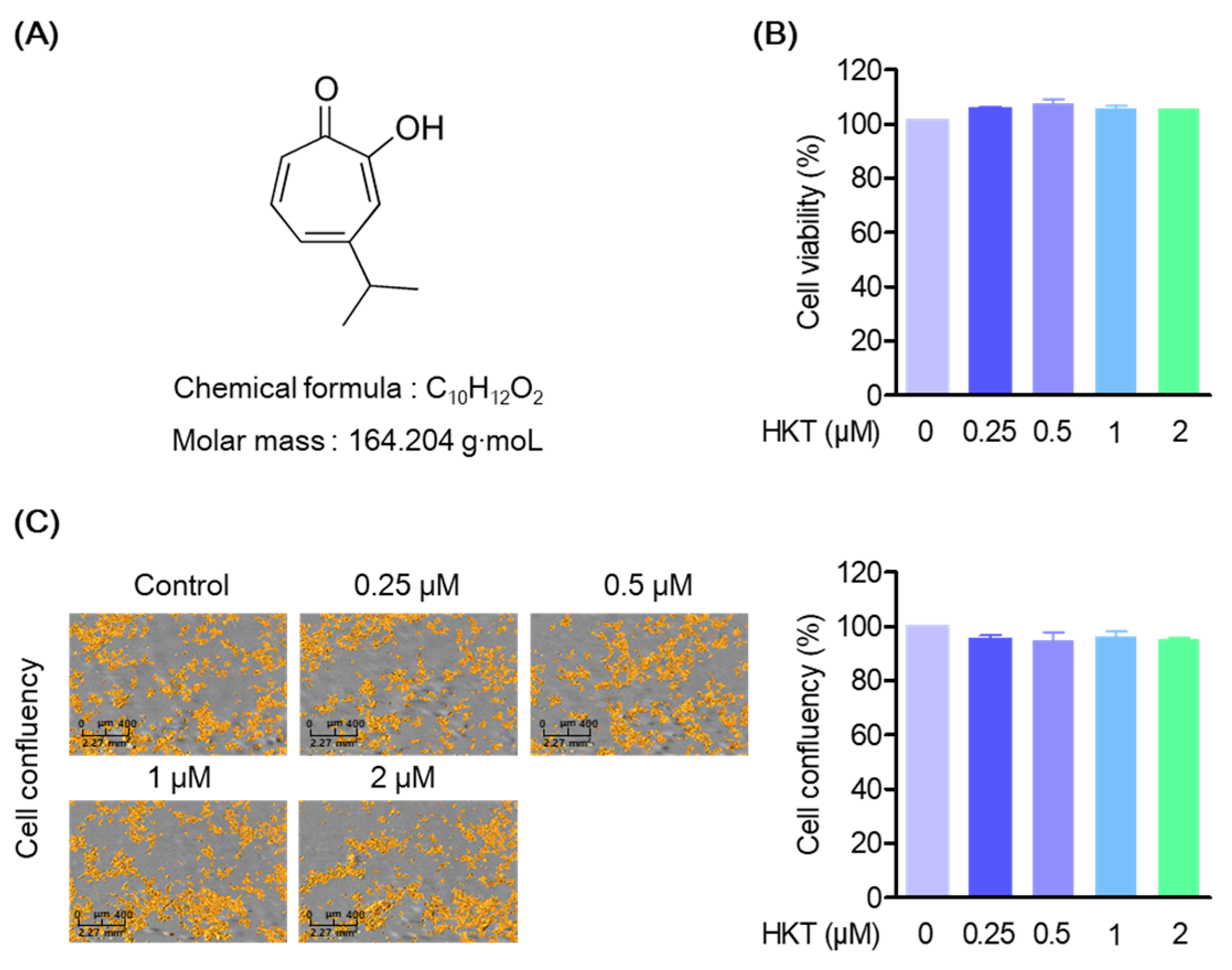
2.3. Cell Viability of HKT
2.4. Evaluation of Inhibition of ROS Generation by HKT
2.5. RT-qPCR
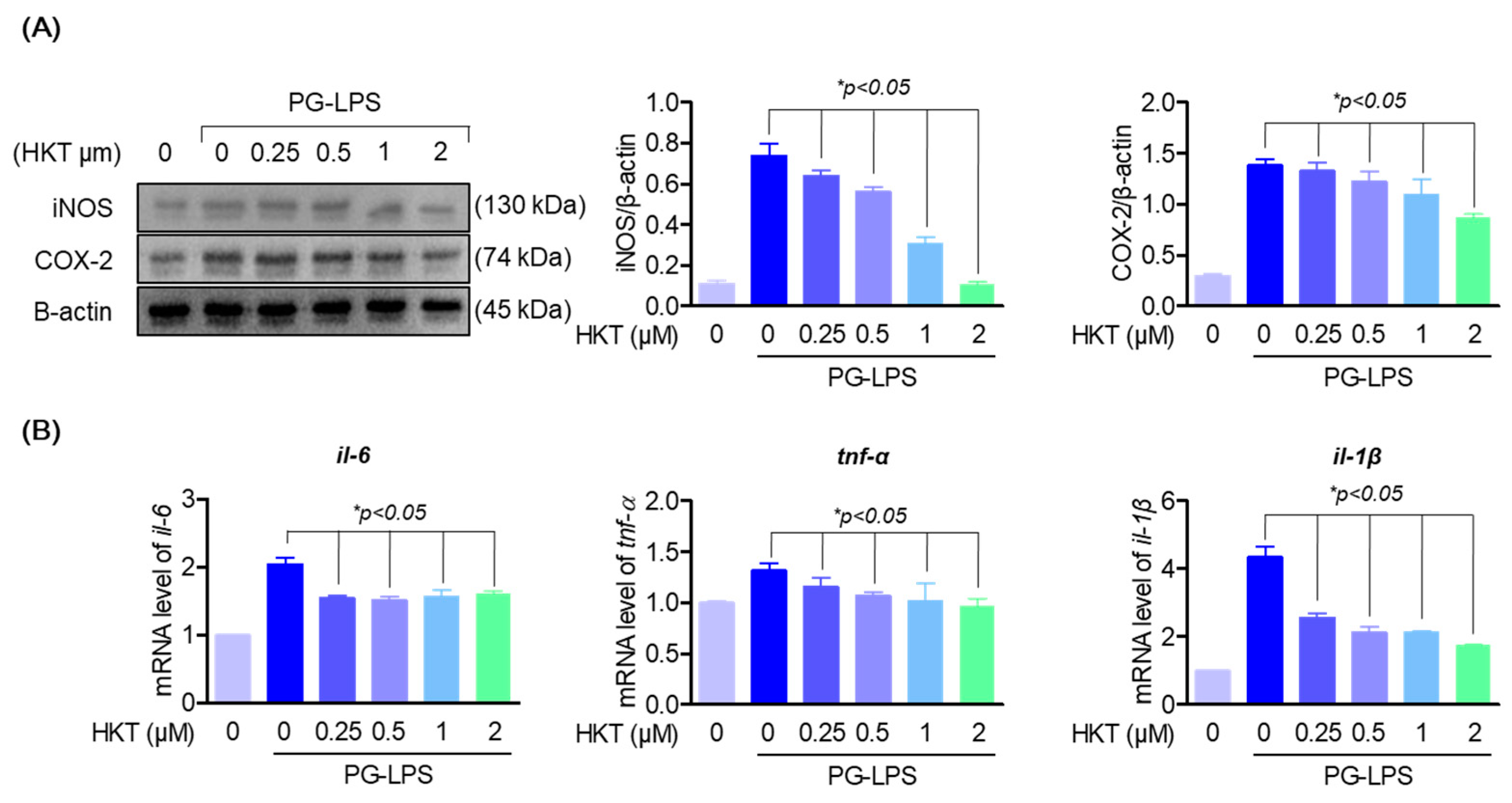
2.6. Western Blotting Analysis
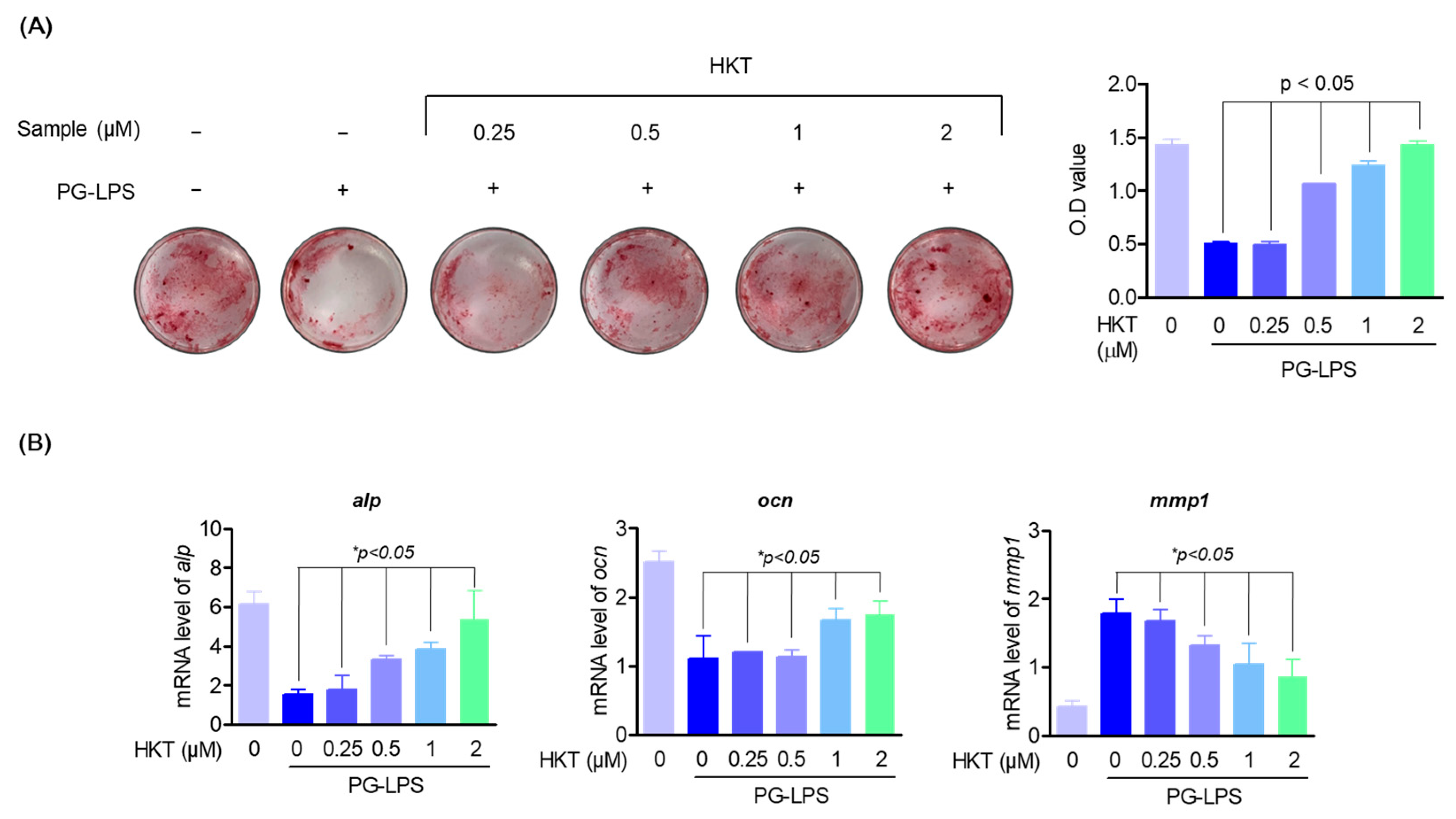
2.7. Mineralization Assay
2.8. Animals
2.9. Ligature-Induced Periodontal Inflammation Model
2.10. Micro-Computed Tomography (Micro-CT) Imaging and Analysis
2.11. Histological Staining
2.12. Statistical Analysis
3. Results
3.1. HKT Is Not Cytotoxic in HPDL Cell
3.2. HKT Inhibits Oxidative Stress Generated by PG-LPS
3.3. HKT Restores the Induction of Osteoblast Differentiation Suppressed by PG-LPS.
3.4. HKT Downregulate PG-LPS-Induced Expression of Pro-Inflammatory Cytokines and Mediators in HPDL Cells
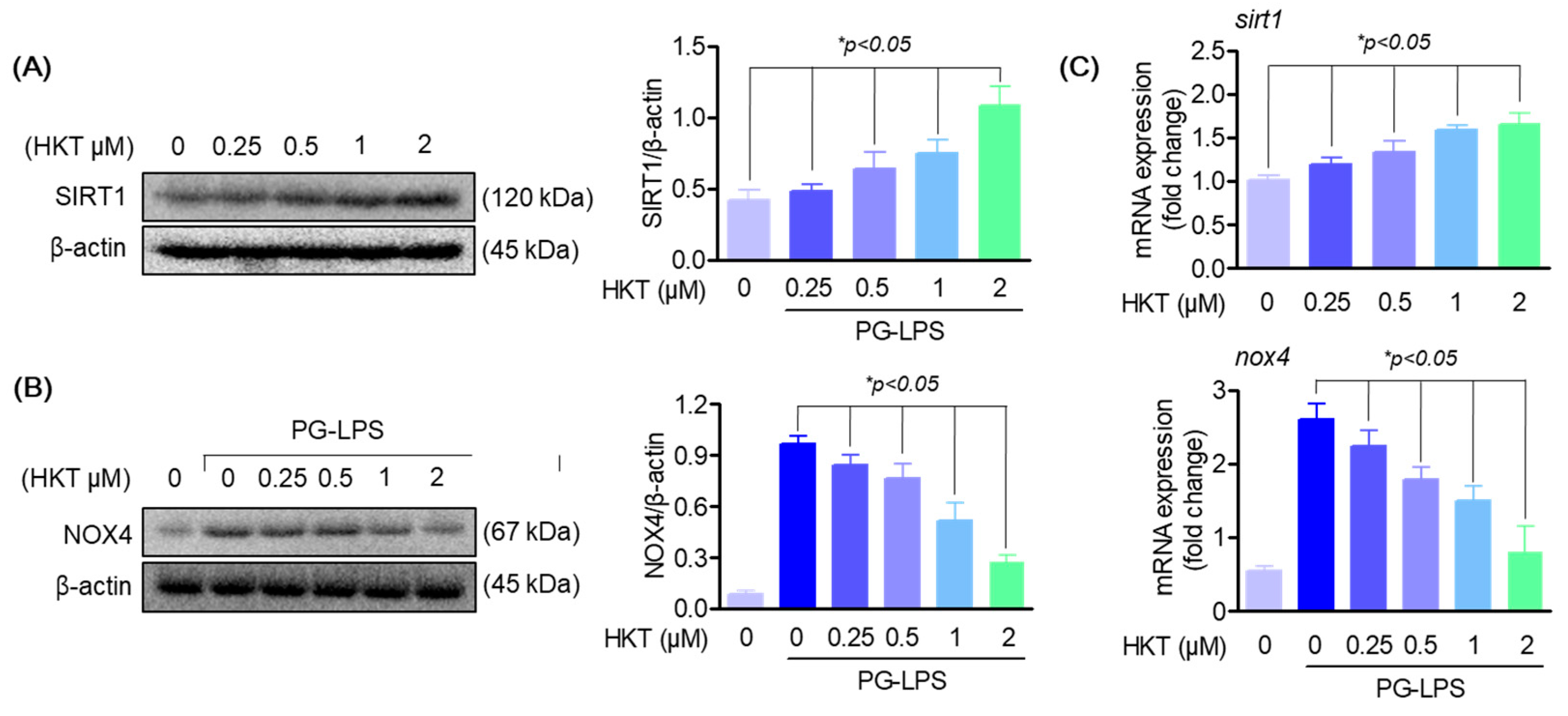
3.5. HKT Regulates SIRT1/NOX4 Protein Expression
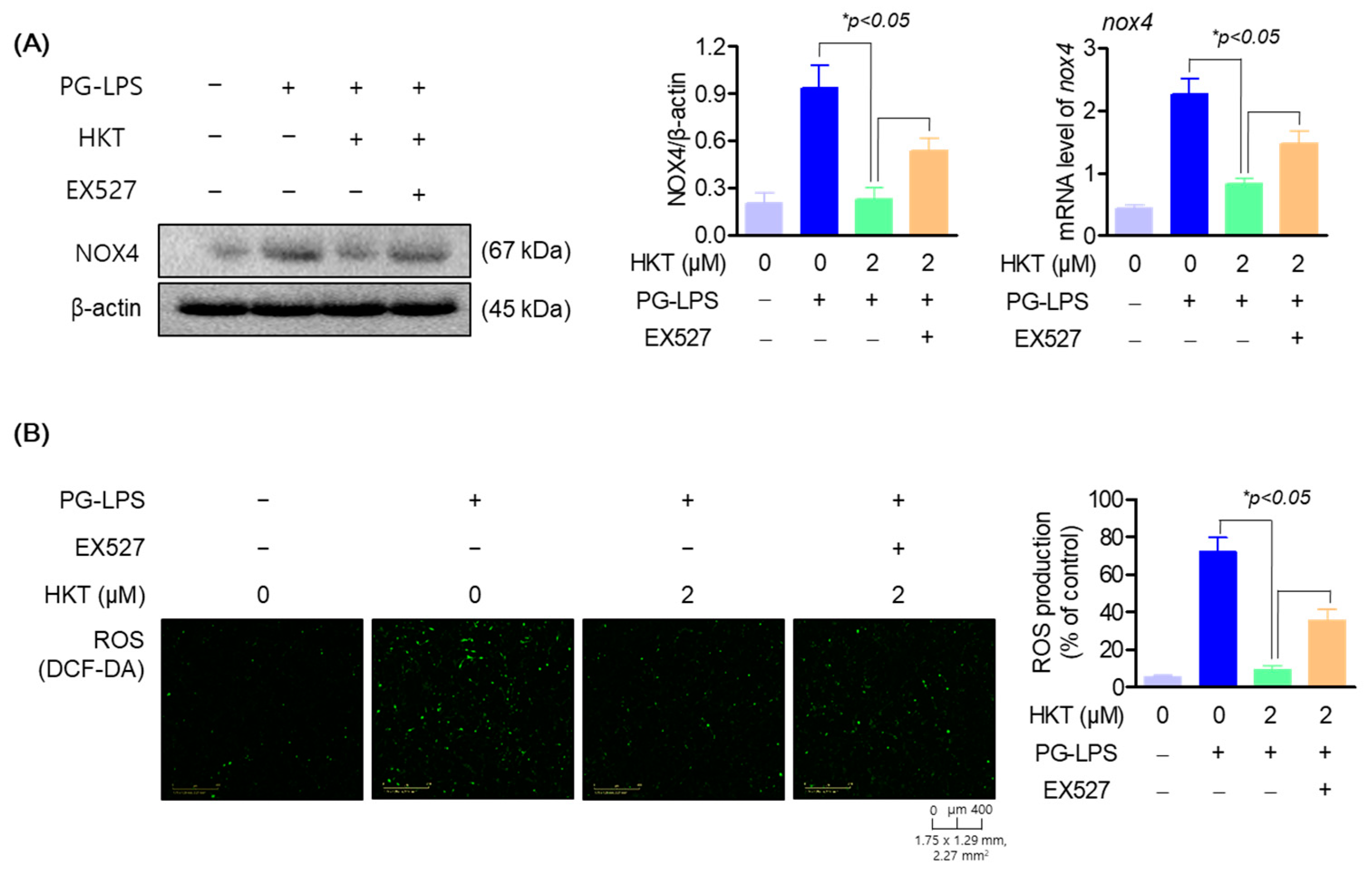
3.6. HKT Alleviates ROS Production by Regulating the SIRT1/NOX4 Pathway
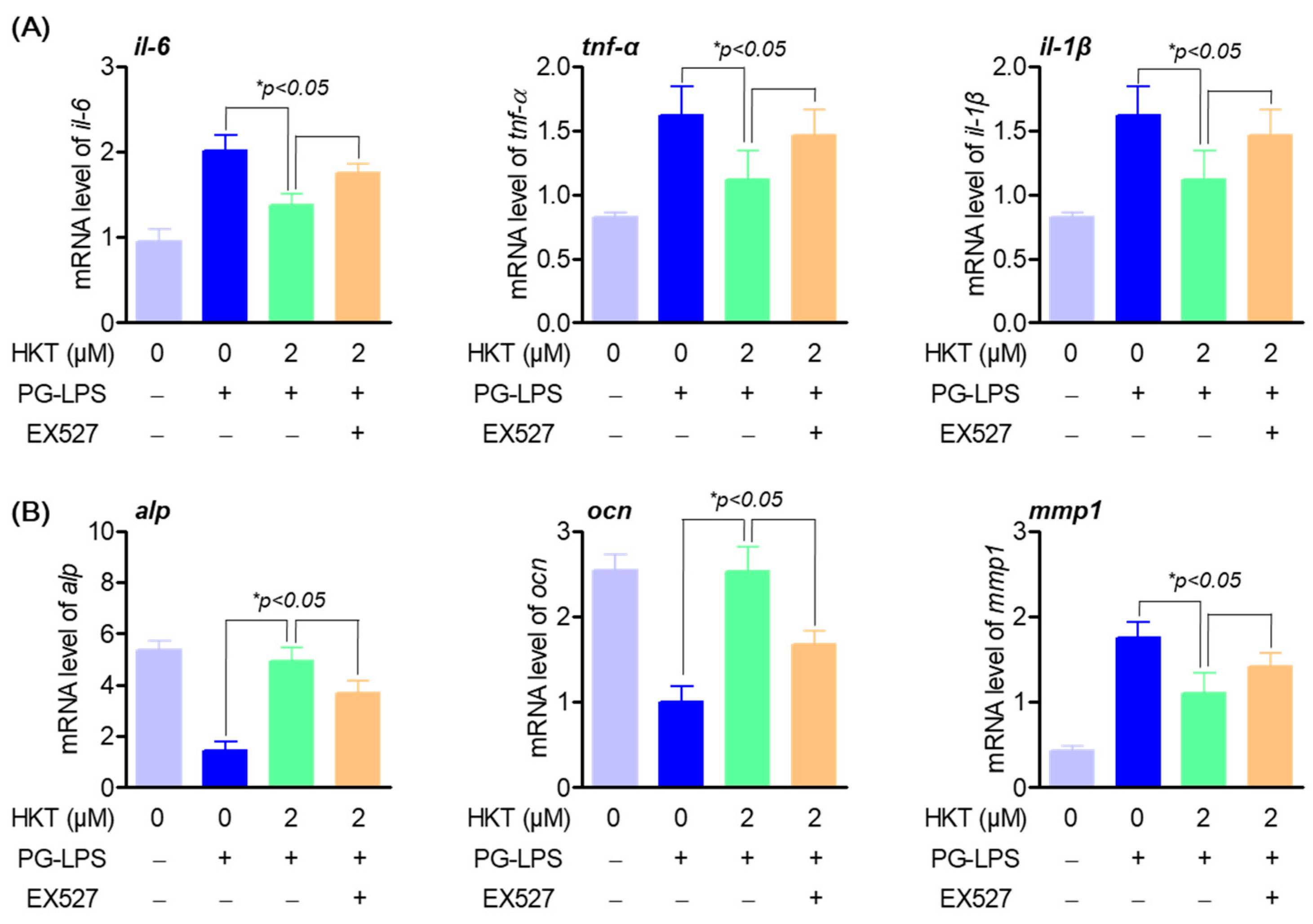
3.7. HKT Regulates Pro-Inflammatory Cytokines and Osteoblast Differentiation-Inducing Specific Genes through the SIRT1/NOX4 Pathway
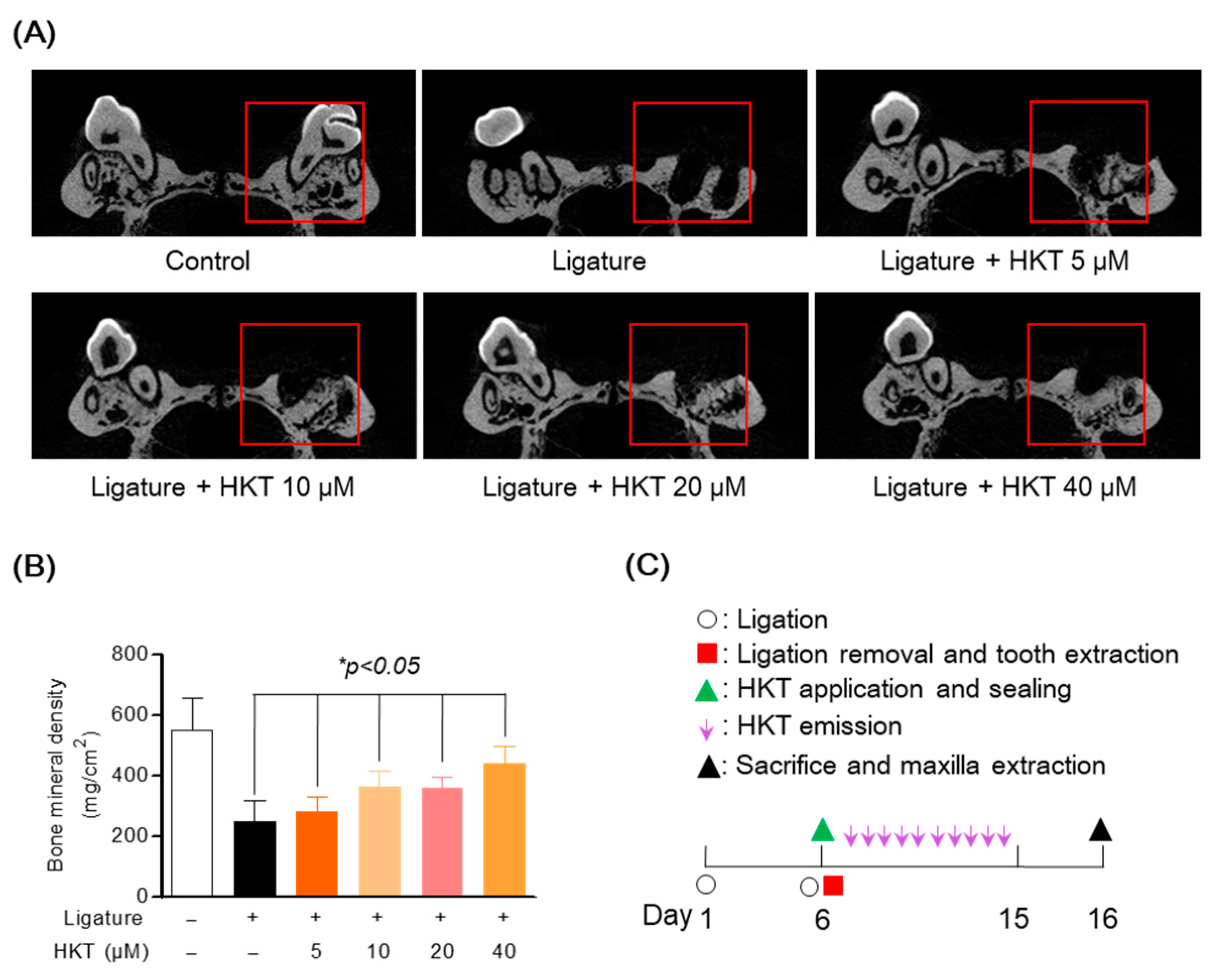
3.8. HKT Recovers Damaged Alveolar Bone in a Periodontal Inflammation Model
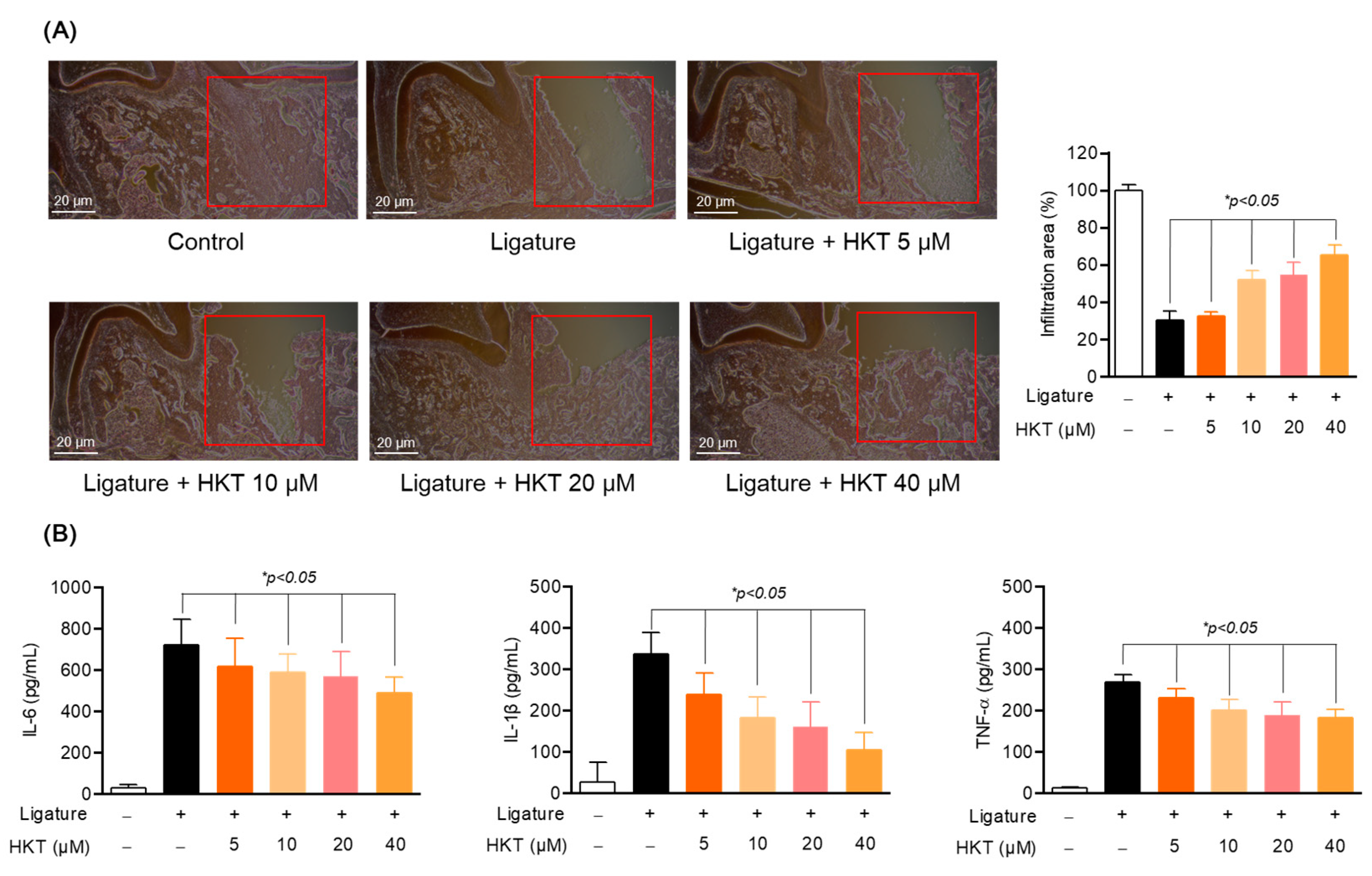
3.9. HKT Exhibits Periodontal Tissue Recovery and Anti-Inflammatory Effects in a Periodontal Inflammation Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haffajee, A.D.; Socransky, S.S. Microbial etiological agents of destructive periodontal diseases. Periodontology 2000 1994, 5, 78–111. [Google Scholar] [CrossRef] [PubMed]
- Baelum, V.; López, R. Periodontal disease epidemiology—Learned and unlearned? Periodontology 2000 2013, 62, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Nazir, M.A. Prevalence of periodontal disease, its association with systemic diseases and prevention. Int. J. Health Sci. 2017, 11, 72–80. [Google Scholar]
- Krutyhołowa, A.; Strzelec, K.; Dziedzic, A.; Bereta, G.P.; Łazarz-Bartyzel, K.; Potempa, J.; Gawron, K. Host and bacterial factors linking periodontitis and rheumatoid arthritis. Front. Immunol. 2022, 13, 980805. [Google Scholar] [CrossRef] [PubMed]
- Fatima, Z.; Shahzadi, C.; Nosheen, A.; Khan, M.; Rehman, H.U. Periodontitis is a risk factor for developing cardiovascular diseases. J. Pak. Med. Assoc. 2020, 70, 1941–1943. [Google Scholar] [PubMed]
- Salhi, L.; Reners, M. Update on the Bidirectional Link Between Diabetes and Periodontitis. Adv. Exp. Med. Biol. 2022, 1373, 231–240. [Google Scholar] [PubMed]
- Slots, J. Periodontitis: Facts, fallacies and the future. Periodontology 2000 2017, 75, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yoneda, M.; Hirofuji, T. Mixed red-complex bacterial infection in periodontitis. Int. J. Dent. 2013, 2013, 587279. [Google Scholar] [CrossRef] [PubMed]
- Palm, E.; Demirel, I.; Bengtsson, T.; Khalaf, H. The role of toll-like and protease-activated receptors and associated intracellular signaling in Porphyromonas gingivalis-infected gingival fibroblasts. APMIS 2017, 125, 157–169. [Google Scholar] [CrossRef]
- Oz, H.S.; Puleo, D.A. Animal models for periodontal disease. J. Biomed. Biotechnol. 2011, 2011, 754857. [Google Scholar] [CrossRef]
- Nakajima, K.; Hamada, N.; Takahashi, Y.; Sasaguri, K.; Tsukinoki, K.; Umemoto, T.; Sato, S. Restraint stress enhances alveolar bone loss in an experimental rat model. J. Periodontal Res. 2006, 41, 527–534. [Google Scholar] [CrossRef] [PubMed]
- De Molon, R.S.; Mascarenhas, V.I.; de Avila, E.D.; Finoti, L.S.; Toffoli, G.B.; Spolidorio, D.M.; Scarel-Caminaga, R.M.; Tetradis, S.; Cirelli, J.A. Long-term evaluation of oral gavage with periodontopathogens or ligature induction of experimental periodontal disease in mice. Clin. Oral Investig. 2016, 20, 1203–1216. [Google Scholar] [CrossRef] [PubMed]
- Kesavalu, L.; Bakthavatchalu, V.; Rahman, M.M.; Su, J.; Raghu, B.; Dawson, D.; Fernandes, G.; Ebersole, J.L. Omega-3 fatty acid regulates inflammatory cytokine/mediator messenger RNA expression in Porphyromonas gingivalis-induced experimental periodontal disease. Oral Microbiol. Immunol. 2007, 22, 232–239. [Google Scholar] [CrossRef]
- Yang, Y.J.; Song, J.H.; Yang, J.H.; Kim, M.J.; Kim, K.Y.; Kim, J.K.; Jin, Y.B.; Kim, W.H.; Kim, S.; Kim, K.R.; et al. Anti-Periodontitis Effects of Dendropanax morbiferus H. Lév Leaf Extract on Ligature-Induced Periodontitis in Rats. Molecules 2023, 28, 849. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Hajishengallis, G. Optimization of the ligature-induced periodontitis model in mice. J. Immunol. Methods 2013, 394, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.J.; Ruddy, M.J.; Wong, G.C.; Sfintescu, C.; Baker, P.J.; Smith, J.B.; Evans, R.T.; Gaffen, S.L. An essential role for IL-17 in preventing pathogen-initiated bone destruction: Recruitment of neutrophils to inflamed bone requires IL-17 receptor-dependent signals. Blood 2007, 109, 3794–3802. [Google Scholar] [CrossRef] [PubMed]
- Bast, A.; Kubis, H.; Holtfreter, B.; Ribback, S.; Martin, H.; Schreiner, H.C.; Dominik, M.J.; Breitbach, K.; Dombrowski, F.; Kocher, T.; et al. NADPH Oxidase Contributes to Resistance against Aggregatibacter actinomycetemcomitans-Induced Periodontitis in Mice. Infect. Immun. 2017, 85, e00849-16. [Google Scholar] [CrossRef]
- Nishida, E.; Hara, Y.; Kaneko, T.; Ikeda, Y.; Ukai, T.; Kato, I. Bone resorption and local interleukin-1alpha and interleukin-1beta synthesis induced by Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis lipopolysaccharide. J. Periodontal Res. 2001, 36, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Estarreja, J.; Pimenta, A.C.; Botelho, J.; Arminda, M.V.; José, J.M.; João, R.; Rui, P.; Vanessa, M.; Vanessa, M. Blood count, endocrine, immunologic, renal, and hepatic markers in a case-control animal study of induced periodontitis in female rodents. Front. Physiol. 2024, 15, 1327399. [Google Scholar] [CrossRef]
- Peiya, L.; Hiromi, N.; Ohsugi, Y.; Tsuchiya, Y.; Shimohira, T.; Komatsu, K.; Liu, A.; Shiba, T.; Aoki, A.; Iwata, T.; et al. Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease. Int. J. Mol. Sci. 2021, 22, 8900. [Google Scholar] [CrossRef]
- Chipashvili, O.; Bor, B. Ligature-induced periodontitis mouse model protocol for studying Saccharibacteria. STAR Protoc. 2023, 3, 101167. [Google Scholar] [CrossRef]
- Duncan, M.T.; DeLuca, T.A.; Kuo, H.Y.; Yi, M.; Mrksich, M.; Miller, W.M. SIRT1 is a critical regulator of K562 cell growth, survival, and differentiation. Exp. Cell Res. 2016, 344, 40–52. [Google Scholar] [CrossRef]
- Imai, S.; Guarente, L. Ten years of NAD-dependent SIR2 family deacetylases: Implications for metabolic diseases. Trends Pharmacol. Sci. 2010, 31, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.R.; Wright, J.; Bauter, M.; Seweryniak, K.; Kode, A.; Rahman, I. Sirtuin regulates cigarette smoke-induced proinflammatory mediator release via RelA/p65 NF-kappaB in macrophages in vitro and in rat lungs in vivo: Implications for chronic inflammation and aging. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L567–L576. [Google Scholar] [CrossRef]
- Li, K.; Lv, G.; Pan, L. Sirt1 alleviates LPS induced inflammation of periodontal ligament fibroblasts via downregulation of TLR4. Int. J. Biol. Macromol. 2018, 119, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Xu, Y.; Yang, W.; Chu, K.; Li, S.; Guo, L. N-acetylcysteine protects human periodontal ligament fibroblasts from pyroptosis and osteogenic differentiation dysfunction through the SIRT1/NF-κB/Caspase-1 signaling pathway. Arch. Oral Biol. 2023, 148, 105642. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Shin, S.I.; Shin, K.S.; Lee, Y.R.; Park, B.H.; Kim, E.C. The role of sirtuin 1 in osteoblastic differentiation in human periodontal ligament cells. J. Periodontal Res. 2011, 46, 712–721. [Google Scholar] [CrossRef]
- Chen, F.; Haigh, S.; Barman, S.; Fulton, D.J. From form to function: The role of Nox4 in the cardiovascular system. Front. Physiol. 2012, 3, 412. [Google Scholar] [CrossRef]
- Hoff, P.; Buttgereit, F. NADPH oxidase 4 represents a potential target for the treatment of osteoporosis. Cell. Mol. Immunol. 2014, 11, 317–319. [Google Scholar] [CrossRef]
- Zhai, L.; Pei, H.; Yang, Y.; Zhu, Y.; Ruan, S. NOX4 promotes Kupffer cell inflammatory response via ROS-NLRP3 to aggravate liver inflammatory injury in acute liver injury. Aging 2022, 14, 6905–6916. [Google Scholar] [CrossRef]
- Gölz, L.; Memmert, S.; Rath-Deschner, B.; Jäger, A.; Appel, T.; Baumgarten, G.; Götz, W.; Frede, S. LPS from P. gingivalis and hypoxia increases oxidative stress in periodontal ligament fibroblasts and contributes to periodontitis. Mediators Inflamm. 2014, 2014, 986264. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zheng, L.; Zhang, B.; Deng, Z.; Li, H. Synergistic protection of quercetin and lycopene against oxidative stress via SIRT1-Nox4-ROS axis in HUVEC cells. Curr. Res. Food Sci. 2022, 5, 1985–1993. [Google Scholar] [CrossRef]
- Hacioglu, C. Capsaicin inhibits cell proliferation by enhancing oxidative stress and apoptosis through SIRT1/NOX4 signaling pathways in HepG2 and HL-7702 cells. J. Biochem. Mol. Toxicol. 2022, 36, e22974. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Chen, Z.; Xia, Y.; Lu, D.; Yang, X.; Sun, A.; Zou, Y.; Qian, J.; Ge, J. Liraglutide attenuates NLRP3 inflammasome-dependent pyroptosis via regulating SIRT1/NOX4/ROS pathway in H9c2 cells. Biochem. Biophys. Res. Commun. 2018, 499, 267–272. [Google Scholar] [CrossRef]
- Shih, Y.H.; Chang, K.W.; Hsia, S.M.; Yu, C.C.; Fuh, L.J.; Chi, T.Y.; Shieh, T.M. In vitro antimicrobial and anticancer potential of hinokitiol against oral pathogens and oral cancer cell lines. Microbiol. Res. 2013, 168, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Trust, T.J. Antibacterial activity of tropolone. Antimicrob. Agents Chemother. 1975, 7, 500–506. [Google Scholar] [CrossRef]
- Jayakumar, T.; Liu, C.H.; Wu, G.Y.; Lee, T.Y.; Manubolu, M.; Hsieh, C.Y.; Yang, C.H.; Sheu, J.R. Hinokitiol Inhibits Migration of A549 Lung Cancer Cells via Suppression of MMPs and Induction of Antioxidant Enzymes and Apoptosis. Int. J. Mol. Sci. 2018, 19, 939. [Google Scholar] [CrossRef]
- Chen, H.Y.; Cheng, W.P.; Chiang, Y.F.; Hong, Y.H.; Ali, M.; Huang, T.C.; Wang, K.L.; Shieh, T.M.; Chang, H.Y.; Hsia, S.M. Hinokitiol Exhibits Antitumor Properties through Induction of ROS-Mediated Apoptosis and p53-Driven Cell-Cycle Arrest in Endometrial Cancer Cell Lines (Ishikawa, HEC-1A, KLE). Int. J. Mol. Sci. 2021, 22, 8268. [Google Scholar] [CrossRef]
- Yang, S.C.; Chen, H.Y.; Chuang, W.L.; Wang, H.C.; Hsieh, C.P.; Huang, Y.F. Different Cell Responses to Hinokitiol Treatment Result in Senescence or Apoptosis in Human Osteosarcoma Cell Lines. Int. J. Mol. Sci. 2022, 23, 1632. [Google Scholar] [CrossRef]
- Ye, J.; Xu, Y.F.; Lou, L.X.; Jin, K.; Miao, Q.; Ye, X.; Xi, Y. Anti-inflammatory effects of hinokitiol on human corneal epithelial cells: An in vitro study. Eye 2015, 29, 964–971. [Google Scholar] [CrossRef]
- Lee, J.H.; Moon, J.H.; Lee, Y.J.; Park, S.Y. SIRT1, a Class III Histone Deacetylase, Regulates LPS-Induced Inflammation in Human Keratinocytes and Mediates the Anti-Inflammatory Effects of Hinokitiol. J. Investig. Dermatol. 2017, 137, 1257–1266. [Google Scholar] [CrossRef] [PubMed]
- Byeon, S.E.; Lee, Y.G.; Kim, J.C.; Han, J.G.; Lee, H.Y.; Cho, J.Y. Hinokitiol, a natural tropolone derivative, inhibits TNF-alpha production in LPS-activated macrophages via suppression of NF-kappaB. Planta Med. 2008, 74, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, T.; Hsu, W.H.; Yen, T.L.; Luo, J.Y.; Kuo, Y.C.; Fong, T.H.; Sheu, J.R. Hinokitiol, a natural tropolone derivative, offers neuroprotection from thromboembolic stroke in vivo. Evid.-Based Complement. Altern. Med. 2013, 2013, 840487. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Zhang, Z.; Wang, Z.; Wu, Q.; He, Y.; Xu, Y.; Ding, Z.; Zhao, H.; Da, H.; Zhang, F.; et al. Hinokitiol functions as a ferroptosis inhibitor to confer neuroprotection. Free Radic. Biol. Med. 2022, 190, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Q.; Fu, Z.; Sun, P.; Zhang, T.; Wang, K.; Li, X.; Qian, Y. Hinokitiol inhibits RANKL-induced osteoclastogenesis in vitro and prevents ovariectomy-induced bone loss in vivo. Int. Immunopharmacol. 2021, 96, 107619. [Google Scholar] [CrossRef]
- Tang, H.; He, K.; Zhao, K.; Zheng, C.; Wu, W.; Jin, W.; Yang, L.; Xie, B. Protective Effects of Hinokitiol on Neuronal Ferroptosis by Activating the Keap1/Nrf2/HO-1 Pathway in Traumatic Brain Injury. J. Neurotrauma 2024, 41, 734–750. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Carl, A.G.; Gunn, W.G.; Alexandra, P.; Darwin, J.P. An Alizarin red-based assay of mineralization by adherent cells in culture: Comparison with cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar]
- Miyauchi, S.; Kitagaki, J.; Masumoto, R.; Imai, A.; Kobayashi, K.; Nakaya, A.; Kawai, S.; Fujihara, C.; Asano, Y.; Yamashita, M.; et al. Sphingomyelin Phosphodiesterase 3 Enhances Cytodifferentiation of Periodontal Ligament Cells. J. Dent. Res. 2017, 96, 339–346. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Tanaka, N.; Morita, T.; Nezu, A.; Tanimura, A.; Mizoguchi, I.; Tojyo, Y. Signaling mechanisms involved in protease-activated receptor-1-mediated interleukin-6 production by human gingival fibroblasts. J. Pharmacol. Exp. Ther. 2004, 311, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Hiyoshi, T.; Domon, H.; Maekawa, T.; Yonezawa, D.; Kunitomo, E.; Tabeta, K.; Terao, Y. Protective effect of hinokitiol against periodontal bone loss in ligature-induced experimental periodontitis in mice. Arch. Oral Biol. 2020, 112, 104679. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Dong, H.; Huang, N.; Fang, J. Oxidative stress and inflammation regulation of sirtuins: New insights into common oral diseases. Front. Physiol. 2022, 13, 953078. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.C.; Kesavalu, L.; Walker, S.; Genco, C.A. Virulence factors of Porphyromonas gingivalis. Periodontology 2000, 20, 168–238. [Google Scholar] [CrossRef]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas Gingivalis: An Overview of Periodontopathic Pathogen below the Gum Line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef]
- Ema, M.; Harazonoa, A.; Fujiib, S.; Kawashima, K. Evaluation of developmental toxicity of b-thujaplicin (hinokitiol) following oral administration during organogenesis in rats. Food Chem. Toxicol. 2004, 42, 465–470. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence (5′→3′) | Accession Number | |
|---|---|---|---|
| il-6 | Forward | AGTGAGGAACAAGCCAGAGC | NM_000600.4 |
| Reverse | GTCAGGGGTGGTTATTGCAT | ||
| il-1β | Forward | AACCTCTTCGAGGCACAAGG | NM_000576.2 |
| Reverse | GTCCTGGAAGGAGCACTTCAT | ||
| tnf-α | Forward | GCCTCTTCTCCTTCCTGATCGT | NM_000594.2 |
| Reverse | TGAGGGTTTGCTACAACATGGG | ||
| alp | Forward | TGCAGTACGAGCTGAACAGG | NM_000478 |
| Reverse | GTCAATTCTGCCTCCTTCCA | ||
| ocn | Forward | CGCTACCTGTATCAATGGCTGG | NM_199173 |
| Reverse | CTCCTGAAAGCCGATGTGGTCA | ||
| mmp-1 | Forward | ATGAAGCAGCCCAGATGTGGAG | NM_002421 |
| Reverse | TGGTCCACATCTGCTCTTGGCA | ||
| sirt1 | Forward | TAGACACGCTGGAACAGGTTGC | NM_012238.5 |
| Reverse | CTCCTCGTACAGCTTCACAGTC | ||
| nox4 | Forward | GCCAGAGTATCACTACCTCCAC | NM_016931 |
| Reverse | CTCGGAGGTAAGCCAAGAGTGT | ||
| β-actin | Forward | AGAGCTACGAGCTGCCTGAC | NM_001101 |
| Reverse | AGCACTGTGTTGGCGTACAG | ||
| gapdh | Forward | TGTTCGTCATGGGTGTGAAC | NM_002046 |
| Reverse | GTCTTCTGGGTGGCAGTGAT | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-Y.; Kim, E.-N.; Jeong, G.-S. Therapeutic Effects of Hinokitiol through Regulating the SIRT1/NOX4 against Ligature-Induced Experimental Periodontitis. Antioxidants 2024, 13, 550. https://doi.org/10.3390/antiox13050550
Kim T-Y, Kim E-N, Jeong G-S. Therapeutic Effects of Hinokitiol through Regulating the SIRT1/NOX4 against Ligature-Induced Experimental Periodontitis. Antioxidants. 2024; 13(5):550. https://doi.org/10.3390/antiox13050550
Chicago/Turabian StyleKim, Tae-Yeon, Eun-Nam Kim, and Gil-Saeng Jeong. 2024. "Therapeutic Effects of Hinokitiol through Regulating the SIRT1/NOX4 against Ligature-Induced Experimental Periodontitis" Antioxidants 13, no. 5: 550. https://doi.org/10.3390/antiox13050550
APA StyleKim, T.-Y., Kim, E.-N., & Jeong, G.-S. (2024). Therapeutic Effects of Hinokitiol through Regulating the SIRT1/NOX4 against Ligature-Induced Experimental Periodontitis. Antioxidants, 13(5), 550. https://doi.org/10.3390/antiox13050550