

The Mitochondrial-Derived Peptide MOTS-c Alleviates Radiation Pneumonitis via an Nrf2-Dependent Mechanism

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experimental Design
2.2. Immunohistochemistry Staining
2.3. Histological Analysis
2.4. Collection and Analysis of Bronchoalveolar Lavage Fluid
2.5. RNA-Sequence Data Acquirement
2.6. Cell Cultures and Reagents
2.7. Biochemical Indexes Analysis
2.8. Quantitative Real-Time PCR
2.9. Western Blot
2.10. Immunofluorescence Staining
2.11. TUNEL Staining
2.12. Flow Cytometry
2.13. Detection of Cytosolic ROS and Mitochondrial ROS in MLE-12 Cells
2.14. Mitochondrial Membrane Potential
2.15. Adenosine Triphosphate Content
2.16. Intracellular Calcium Concentration in MLE-12 Cells
2.17. Enzyme-Linked Immunosorbent Assay
2.18. Preparation of Primary AT II Cells
2.19. Statistical Analysis
3. Results
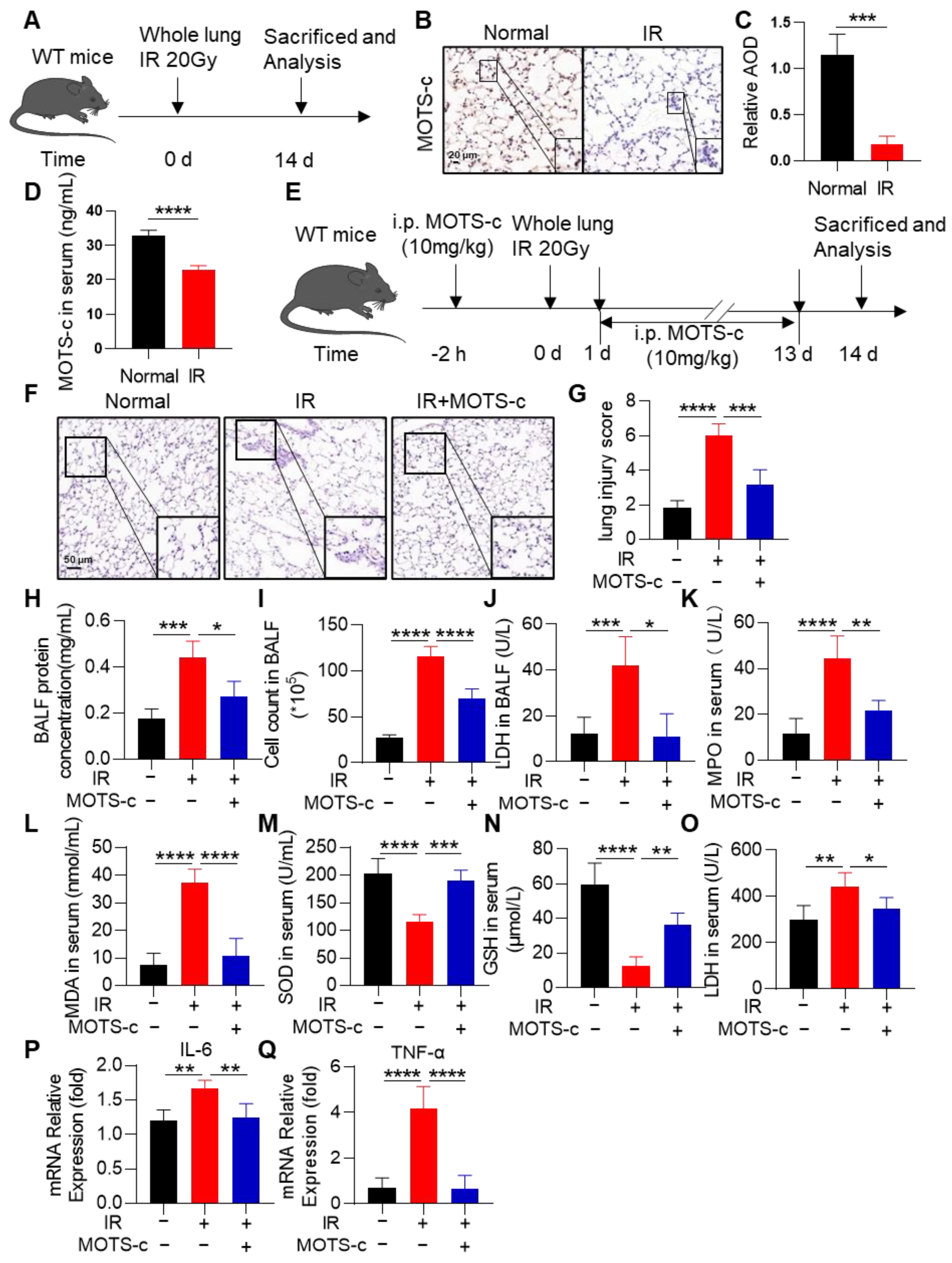
3.1. MOTS-c Alleviated the Oxidant Damage, Inflammation, and Lung Tissue Injury in Irradiated Mice
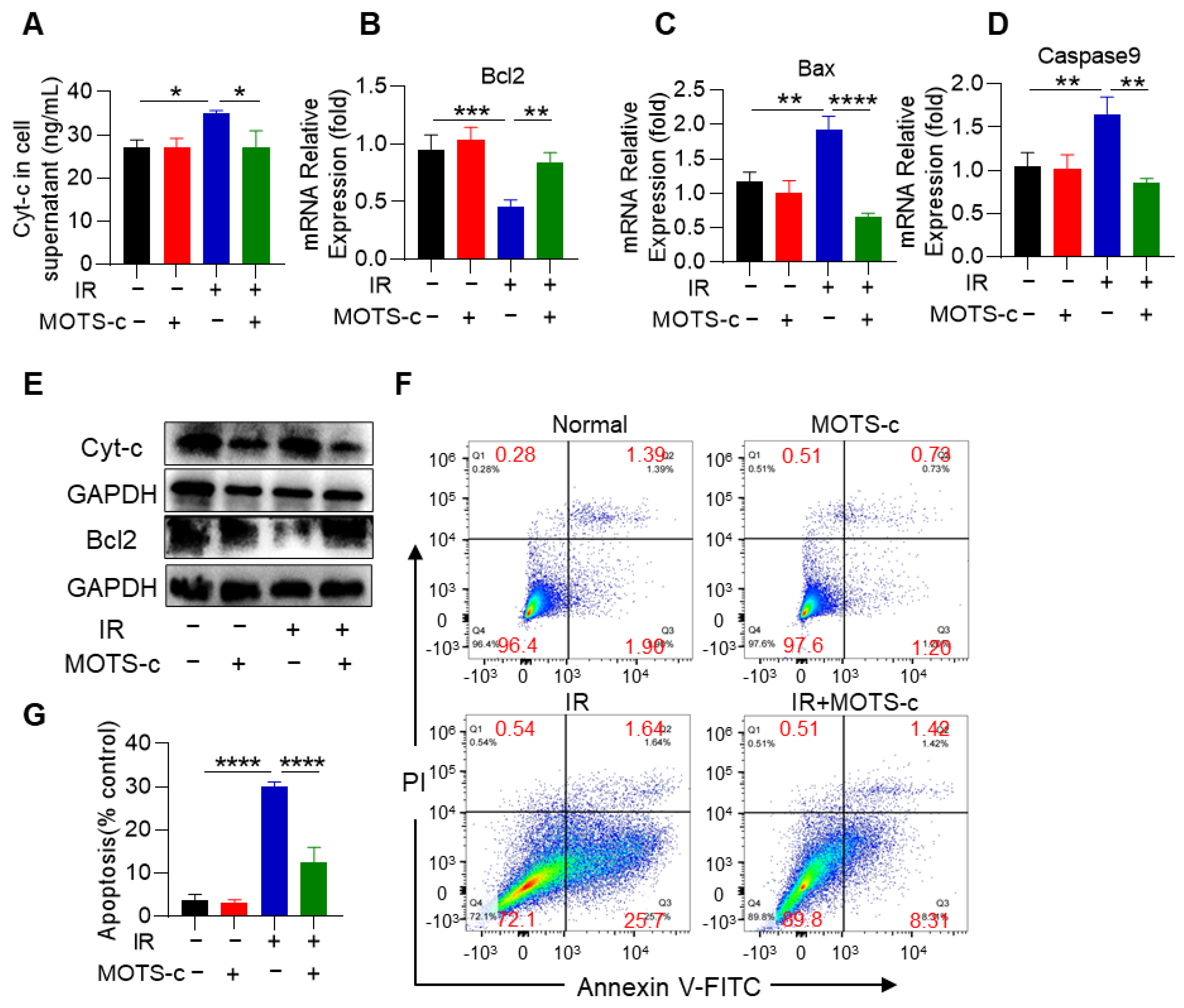
3.2. MOTS-c Prevented the Apoptosis of Alveolar Epithelial Cells in RP Mice
3.3. MOTS-c Alleviates Radiation-Induced Oxidative Stress and Inflammation in MLE-12 Cells
3.4. MOTS-c Reduced the Radiation-Induced Apoptosis of MLE-12 Cells
3.5. MOTS-c Relieved Radiation-Induced Mitochondrial Damage in MLE-12 Cells
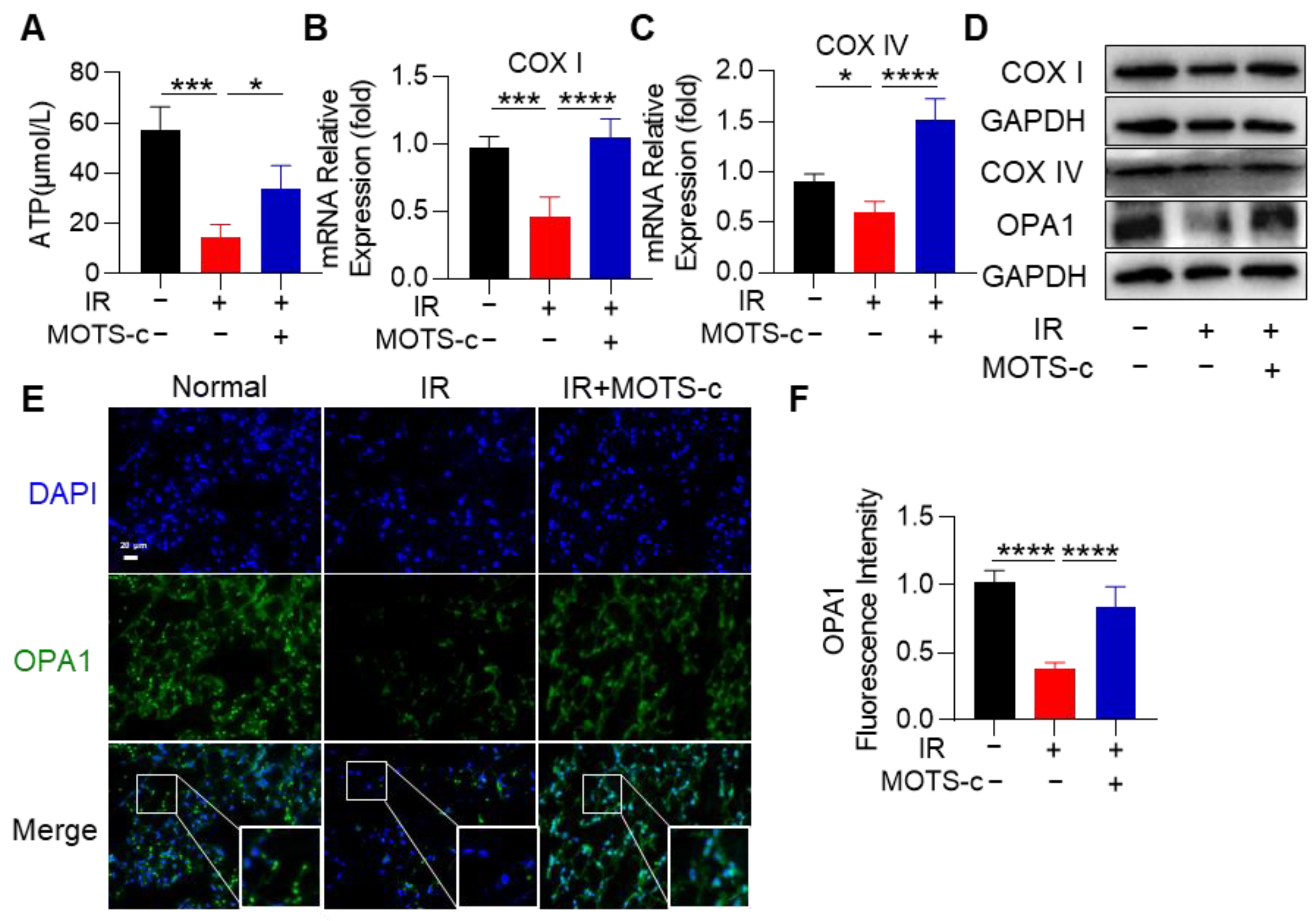
3.6. MOTS-c Mitigated Mitochondrial Injury in RP Mice
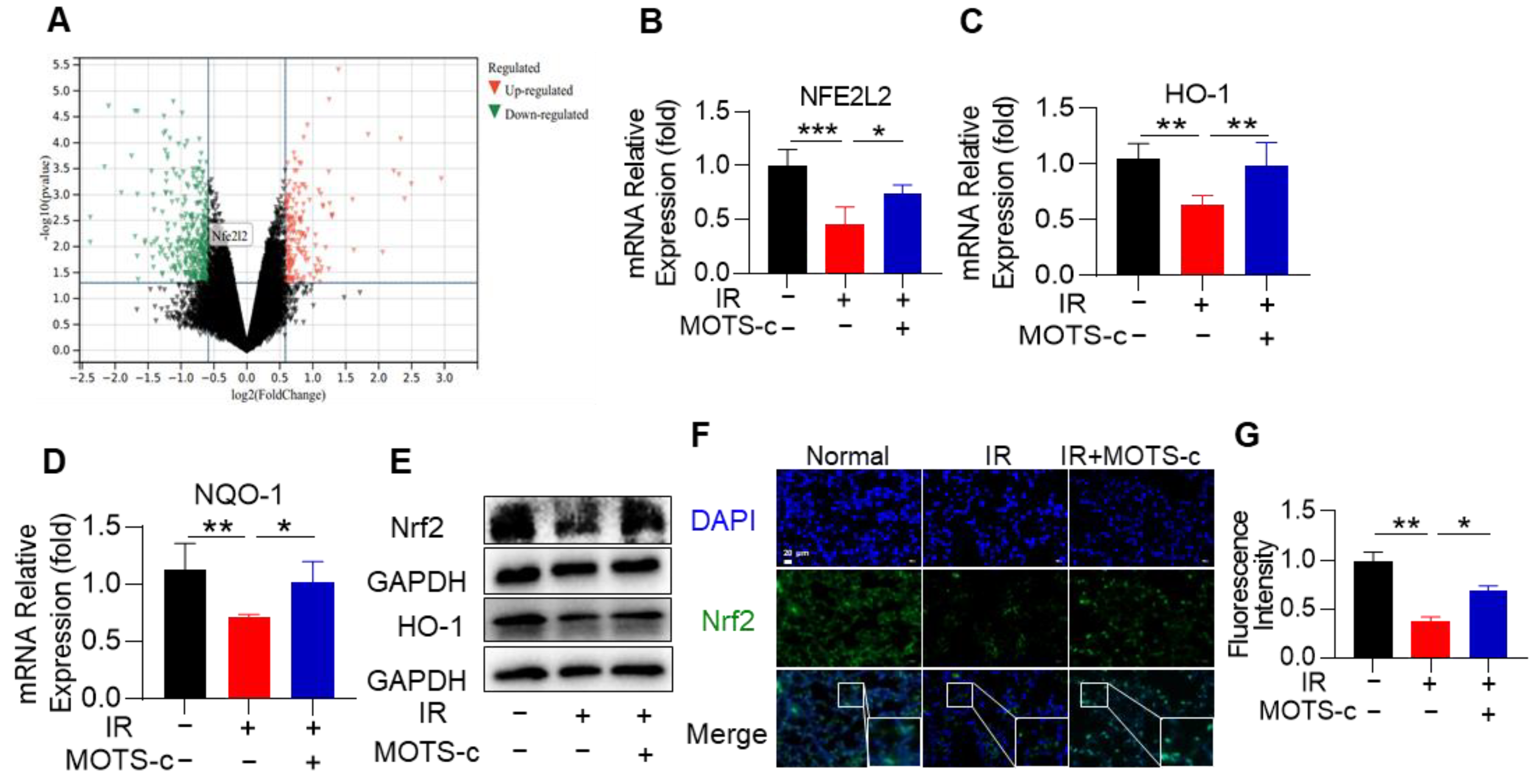
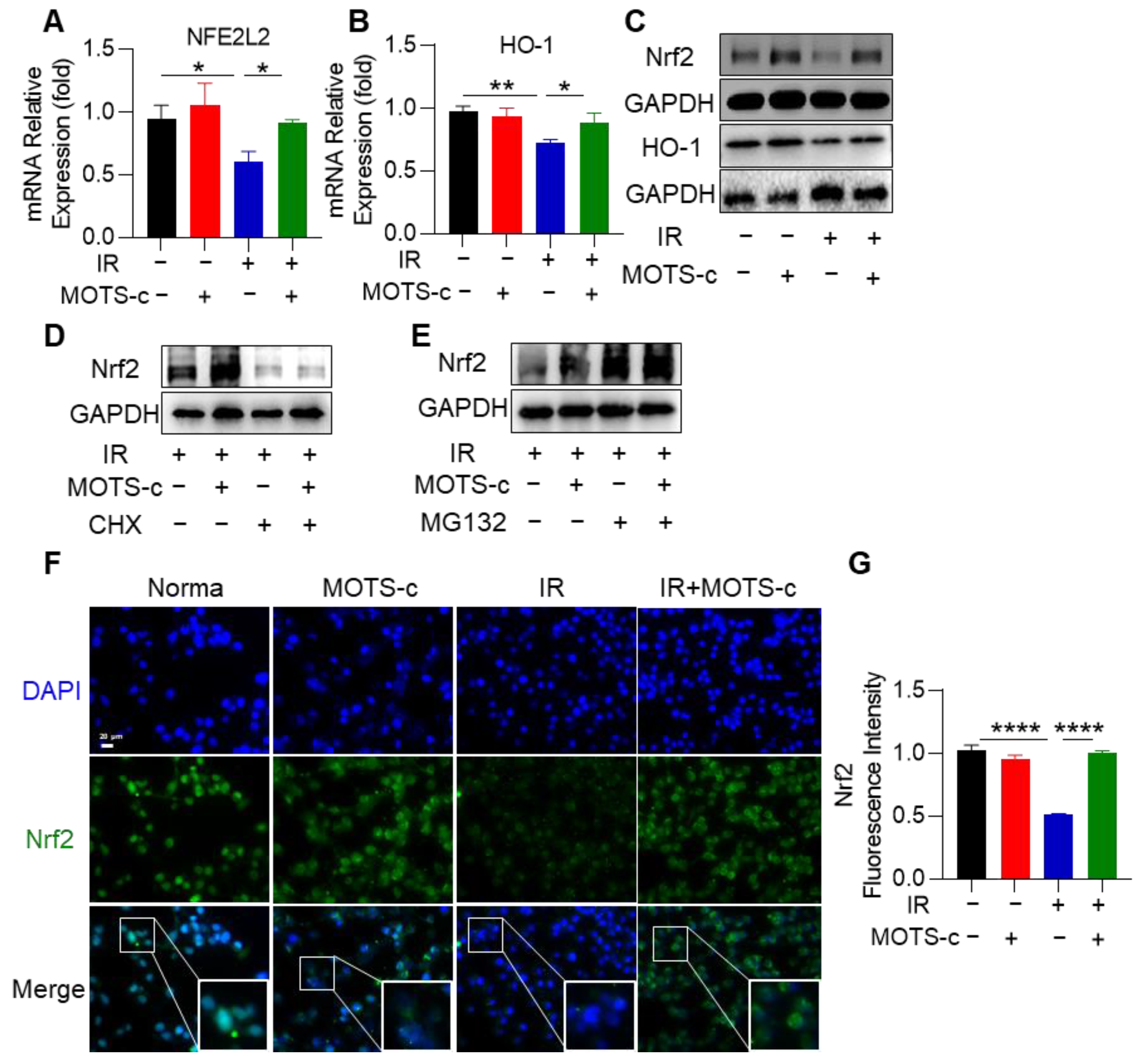
3.7. MOTS-c Increased Nrf2 Content and Induced Its Nucleus Translocation in RP Mice
3.8. MOTS-c Increased Nrf2 Content and Induced Its Nucleus Translocation in MLE-12 Cells
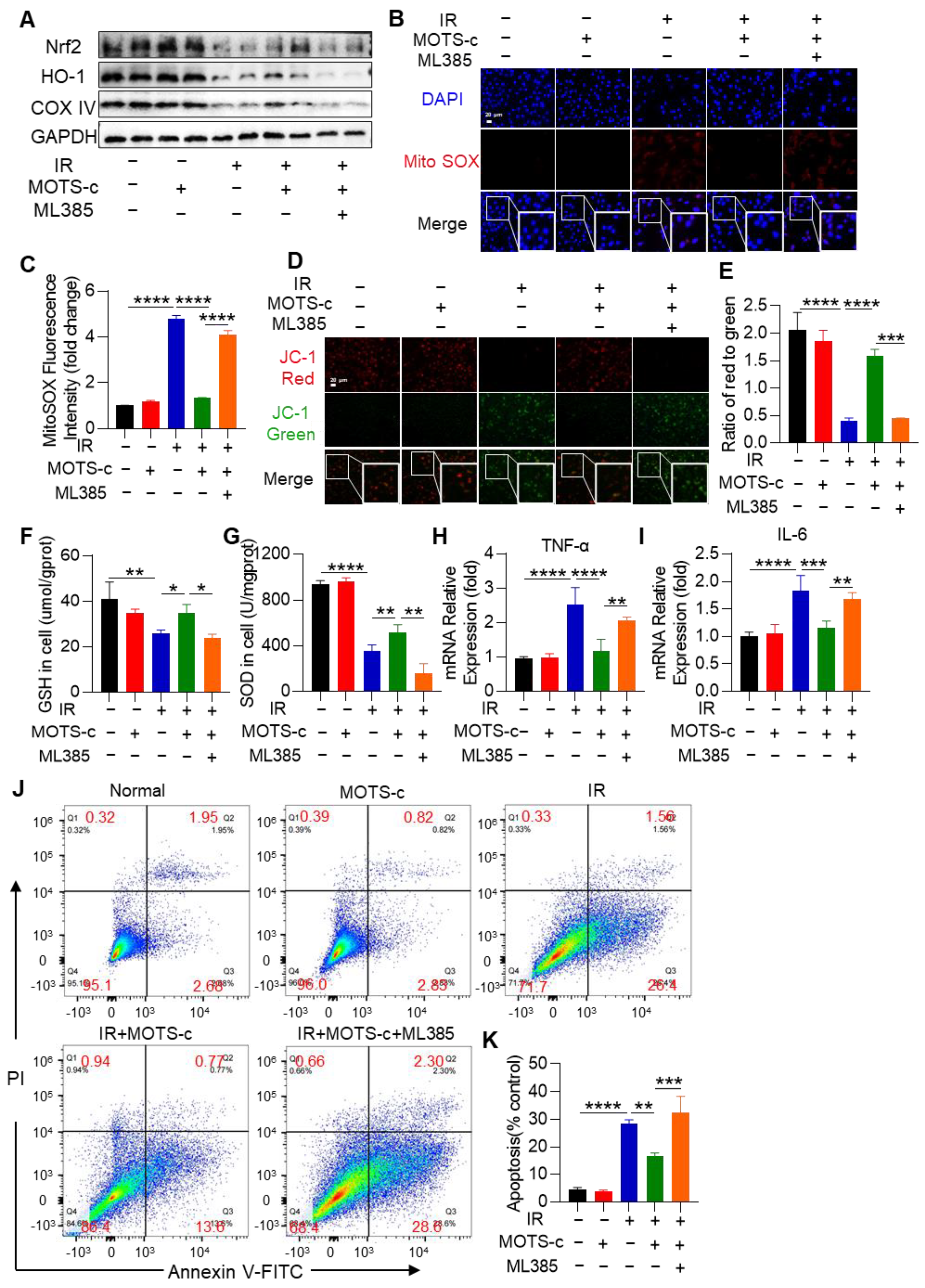
3.9. Inhibition of Nrf2 Abolished the Protective Function of MOTS-c in MLE-12 Cells
3.10. Nrf2 Deficiency Abolished the Protective Function of MOTS-c in RP Mice
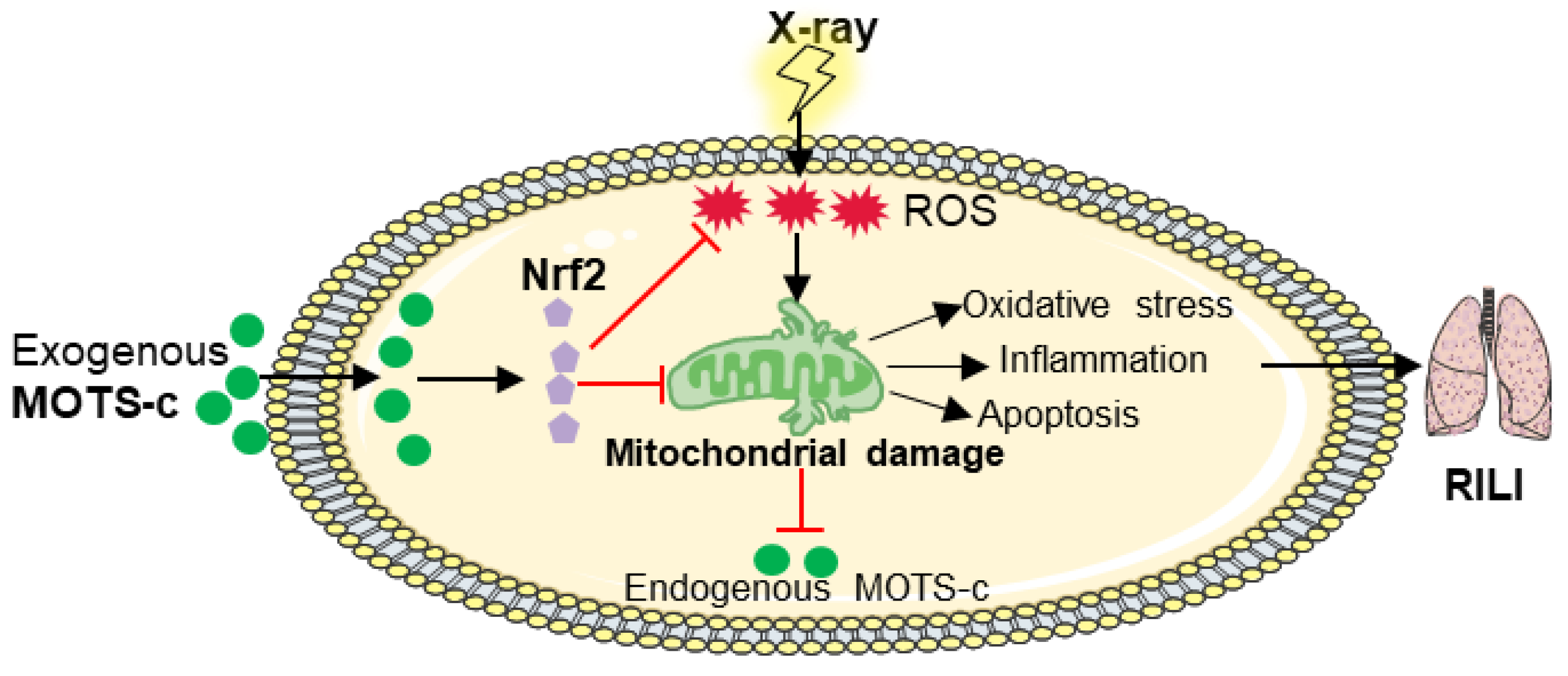
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanania, A.N.; Mainwaring, W.; Ghebre, Y.T.; Hanania, N.A.; Ludwig, M. Radiation-Induced Lung Injury: Assessment and Management. Chest 2019, 156, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shen, Z.; Jiang, X.; Wang, Y.; Yang, Z.; Mao, Y.; Wu, Z.; Li, G.; Chen, H. Mouse mesenchymal stem cell-derived exosomal miR-466f-3p reverses EMT process through inhibiting AKT/GSK3beta pathway via c-MET in radiation-induced lung injury. J. Exp. Clin. Cancer Res. 2022, 41, 128. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Du, L.; Ma, N.; Zhang, P.; Wang, Y.; Han, Y.; Huang, X.; Zhang, Q.; Tan, X.; Lei, X.; et al. Monophosphoryl lipid A ameliorates radiation-induced lung injury by promoting the polarization of macrophages to the M1 phenotype. J. Transl. Med. 2022, 20, 597. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Z.; Ren, Y.; Lei, Y.; Yang, S.; Shi, Y.; Peng, H.; Yang, W.; Guo, T.; Yu, Y.; et al. Mitochondrial-derived peptides in cardiovascular disease: Novel insights and therapeutic opportunities. J. Adv. Res. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qu, X.; Guan, C.; Luo, N.; Chen, H.; Li, A.; Zhuang, H.; Yang, J.; Diao, H.; Zeng, S.; et al. Mitochondrial micropeptide MOXI promotes fibrotic gene transcription by translocation to the nucleus and bridging N-acetyltransferase 14 with transcription factor c-Jun. Kidney Int. 2023, 103, 886–902. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Mehta, H.H.; Wan, J.; Kuehnemann, C.; Chen, J.; Hu, J.-F.; Hoffman, A.R.; Cohen, P. Mitochondrial peptides modulate mitochondrial function during cellular senescence. Aging 2018, 10, 1239–1256. [Google Scholar] [CrossRef]
- Zhong, P.; Peng, J.; Hu, Y.; Zhang, J.; Shen, C. Mitochondrial derived peptide MOTS-c prevents the development of heart failure under pressure overload conditions in mice. J. Cell Mol. Med. 2022, 26, 5369–5378. [Google Scholar] [CrossRef]
- Benayoun, B.A.; Lee, C. MOTS-c: A Mitochondrial-Encoded Regulator of the Nucleus. Bioessays 2019, 41, e1900046. [Google Scholar] [CrossRef]
- Yu, W.D.; Kim, Y.J.; Cho, M.J.; Seok, J.; Kim, G.J.; Lee, C.H.; Ko, J.J.; Kim, Y.S.; Lee, J.H. The mitochondrial-derived peptide MOTS-c promotes homeostasis in aged human placenta-derived mesenchymal stem cells in vitro. Mitochondrion 2021, 58, 135–146. [Google Scholar] [CrossRef]
- Wang, M.; Wang, G.; Pang, X.; Ma, J.; Yuan, J.; Pan, Y.; Fu, Y.; Laher, I.; Li, S. MOTS-c repairs myocardial damage by inhibiting the CCN1/ERK1/2/EGR1 pathway in diabetic rats. Front. Nutr. 2022, 9, 1060684. [Google Scholar] [CrossRef]
- Mohtashami, Z.; Singh, M.K.; Salimiaghdam, N.; Ozgul, M.; Kenney, M.C. MOTS-c, the Most Recent Mitochondrial Derived Peptide in Human Aging and Age-Related Diseases. Int. J. Mol. Sci. 2022, 23, 11991. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Miller, B.; Kumagai, H.; Silverstein, A.R.; Flores, M.; Yen, K. Mitochondrial-derived peptides in aging and age-related diseases. Geroscience 2021, 43, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, R.F.; Woodhead, J.S.T.; Hedges, C.P.; Zeng, N.; Wan, J.; Kumagai, H.; Lee, C.; Cohen, P.; Cameron-Smith, D.; Mitchell, C.J.; et al. Increased expression of the mitochondrial derived peptide, MOTS-c, in skeletal muscle of healthy aging men is associated with myofiber composition. Aging 2020, 12, 5244–5258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed]
- Zhai, D.; Ye, Z.; Jiang, Y.; Xu, C.; Ruan, B.; Yang, Y.; Lei, X.; Xiang, A.; Lu, H.; Zhu, Z.; et al. MOTS-c peptide increases survival and decreases bacterial load in mice infected with MRSA. Mol. Immunol. 2017, 92, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Moreira, P.I.; Carvalho, C.; Zhu, X.; Smith, M.A.; Perry, G. Mitochondrial dysfunction is a trigger of Alzheimer’s disease pathophysiology. Biochim. Biophys. Acta 2010, 1802, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Rustamov, N.; Roh, Y.-S. The Roles of NFR2-Regulated Oxidative Stress and Mitochondrial Quality Control in Chronic Liver Diseases. Antioxidants 2023, 12, 1928. [Google Scholar] [CrossRef] [PubMed]
- Esteras, N.; Abramov, A.Y. Nrf2 as a regulator of mitochondrial function: Energy metabolism and beyond. Free Radic. Biol. Med. 2022, 189, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.M. Nrf2 for protection against oxidant generation and mitochondrial damage in cardiac injury. Free Radic. Biol. Med. 2022, 179, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.M.; Narasimhan, M.; Shanmugam, G.; Milash, B.; Hoidal, J.R.; Rajasekaran, N.S. Differential regulation of miRNA and mRNA expression in the myocardium of Nrf2 knockout mice. BMC Genom. 2017, 18, 509. [Google Scholar] [CrossRef]
- Gao, L.; Kumar, V.; Vellichirammal, N.N.; Park, S.-Y.; Rudebush, T.L.; Yu, L.; Son, W.-M.; Pekas, E.J.; Wafi, A.M.; Hong, J.; et al. Functional, proteomic and bioinformatic analyses of Nrf2- and Keap1- null skeletal muscle. J. Physiol. 2020, 598, 5427–5451. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Q.; Wu, C.; Qiu, Y.B.; Wu, Y.X.; Chen, J.L.; Huang, J.F.; Chen, D.; Pang, Q.F. Heme oxygenase-1 attenuates seawater drowning-induced acute lung injury through a reduction in inflammation and oxidative stress. Int. Immunopharmacol. 2019, 74, 105634. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Ruiz, M.E.; Garasa, S.; Rodriguez, I.; Solorzano, J.L.; Barbes, B.; Yanguas, A.; Teijeira, A.; Etxeberria, I.; Aristu, J.J.; Halin, C.; et al. Intercellular Adhesion Molecule-1 and Vascular Cell Adhesion Molecule Are Induced by Ionizing Radiation on Lymphatic Endothelium. Int. J. Radiat. Oncol. Biol. Phys. 2017, 97, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Pan, Y.; He, J.; Zhong, H.; Wu, Y.; Ji, C.; Liu, L.; Cui, X. The mitochondrial-derived peptide MOTS-c relieves hyperglycemia and insulin resistance in gestational diabetes mellitus. Pharmacol. Res. 2022, 175, 105987. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Huang, K.; Xu, S.; Garcia, J.G.N.; Wang, C.; Cai, H. Targeting NOX4 alleviates sepsis-induced acute lung injury via attenuation of redox-sensitive activation of CaMKII/ERK1/2/MLCK and endothelial cell barrier dysfunction. Redox Biol. 2020, 36, 101638. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.C.; Lai, R.W.; Woodhead, J.S.T.; Joly, J.H.; Mitchell, C.J.; Cameron-Smith, D.; Lu, R.; Cohen, P.; Graham, N.A.; Benayoun, B.A.; et al. MOTS-c is an exercise-induced mitochondrial-encoded regulator of age-dependent physical decline and muscle homeostasis. Nat. Commun. 2021, 12, 470. [Google Scholar] [CrossRef]
- Qiu, Y.B.; Wan, B.B.; Liu, G.; Wu, Y.X.; Chen, D.; Lu, M.D.; Chen, J.L.; Yu, R.Q.; Chen, D.Z.; Pang, Q.F. Nrf2 protects against seawater drowning-induced acute lung injury via inhibiting ferroptosis. Respir. Res. 2020, 21, 232. [Google Scholar] [CrossRef]
- Ares, G.R. cGMP induces degradation of NKCC2 in the thick ascending limb via the ubiquitin-proteasomal system. Am. J. Physiol. Renal Physiol. 2019, 316, F838–F846. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.J.; Jenkinson, H.F.; Davis, S.A.; Mann, S.; O’Sullivan, D.J.; Barbour, M.E. Chlorhexidine hexametaphosphate nanoparticles as a novel antimicrobial coating for dental implants. J. Mater. Sci. Mater. Med. 2015, 26, 201. [Google Scholar] [CrossRef]
- Wu, Y.-X.; Zhang, Y.-R.; Jiang, F.-J.; He, S.; Zhang, Y.-L.; Chen, D.; Tong, Y.; Nie, Y.-J.; Pang, Q.-F. 4-OI ameliorates bleomycin-induced pulmonary fibrosis by activating Nrf2 and suppressing macrophage-mediated epithelial-mesenchymal transition. Inflamm. Res. 2023, 72, 1133–1145. [Google Scholar] [CrossRef]
- Zhang, M.; Qian, J.; Xing, X.; Kong, F.M.; Zhao, L.; Chen, M.; Lawrence, T.S. Inhibition of the tumor necrosis factor-alpha pathway is radioprotective for the lung. Clin. Cancer Res. 2008, 14, 1868–1876. [Google Scholar] [CrossRef]
- Giuranno, L.; Ient, J.; De Ruysscher, D.; Vooijs, M.A. Radiation-Induced Lung Injury (RILI). Front. Oncol. 2019, 9, 877. [Google Scholar] [CrossRef]
- Li, X.; Gong, Y.; Li, D.; Xiang, L.; Ou, Y.; Jiang, L.; Shu, P.; Liu, X.; Guo, F.; Qin, D.; et al. Low-Dose Radiation Therapy Promotes Radiation Pneumonitis by Activating NLRP3 Inflammasome. Int. J. Radiat. Oncol. Biol. Phys. 2020, 107, 804–814. [Google Scholar] [CrossRef]
- Flockerzi, E.; Schanz, S.; Rübe, C.E. Even low doses of radiation lead to DNA damage accumulation in lung tissue according to the genetically-defined DNA repair capacity. Radiother. Oncol. 2014, 111, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Drishya, S.; Dhanisha, S.S.; Raghukumar, P.; Guruvayoorappan, C. Amomum subulatum mitigates experimental thoracic radiation-induced lung injury by regulating antioxidant status and inflammatory responses. Food Funct. 2023, 14, 1545–1559. [Google Scholar] [CrossRef]
- Guo, H.; Chen, J.; Yu, H.; Dong, L.; Yu, R.; Li, Q.; Song, J.; Chen, H.; Zhang, H.; Pu, J.; et al. Activation of Nrf2/ARE pathway by Anisodamine (654-2) for Inhibition of cellular aging and alleviation of Radiation-Induced lung injury. Int. Immunopharmacol. 2023, 124, 110864. [Google Scholar] [CrossRef]
- Yin, Z.; Yang, G.; Deng, S.; Wang, Q. Oxidative stress levels and dynamic changes in mitochondrial gene expression in a radiation-induced lung injury model. J. Radiat. Res. 2019, 60, 204–214. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Rabbani, Z.N.; Jackson, I.L.; Vujaskovic, Z. Oxidative stress mediates radiation lung injury by inducing apoptosis. Int. J. Radiat. Oncol. Biol. Phys. 2012, 83, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Gollapudi, S. Susceptibility of naive and subsets of memory T cells to apoptosis via multiple signaling pathways. Autoimmun. Rev. 2007, 6, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, J.G. First steps in the acquisition of Spanish stress. J. Child. Lang. 1988, 15, 273–292. [Google Scholar] [CrossRef]
- Cen, M.; Ouyang, W.; Zhang, W.; Yang, L.; Lin, X.; Dai, M.; Hu, H.; Tang, H.; Liu, H.; Xia, J.; et al. MitoQ protects against hyperpermeability of endothelium barrier in acute lung injury via a Nrf2-dependent mechanism. Redox Biol. 2021, 41, 101936. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Du, L.; Yu, W.; Wang, Y.; Ma, N.; Qu, B. GSTP1 as a novel target in radiation induced lung injury. J. Transl. Med. 2021, 19, 297. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, M.; Cortesi, M.; Zamagni, A.; Tesei, A. The Role of Mesenchymal Stem Cells in Radiation-Induced Lung Fibrosis. Int. J. Mol. Sci. 2019, 20, 3876. [Google Scholar] [CrossRef] [PubMed]
- Cogno, N.; Bauer, R.; Durante, M. An Agent-Based Model of Radiation-Induced Lung Fibrosis. Int. J. Mol. Sci. 2022, 23, 13920. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Jing, Y.; Chen, Q.; Abbas, A.B.; Hu, J.; Xu, H. The intraperitoneal administration of MOTS-c produces antinociceptive and anti-inflammatory effects through the activation of AMPK pathway in the mouse formalin test. Eur. J. Pharmacol. 2020, 870, 172909. [Google Scholar] [CrossRef] [PubMed]
- Xinqiang, Y.; Quan, C.; Yuanyuan, J.; Hanmei, X. Protective effect of MOTS-c on acute lung injury induced by lipopolysaccharide in mice. Int. Immunopharmacol. 2020, 80, 106174. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gidlund, E.K.; Witasp, A.; Qureshi, A.R.; Soderberg, M.; Thorell, A.; Nader, G.A.; Barany, P.; Stenvinkel, P.; von Walden, F. Reduced skeletal muscle expression of mitochondrial-derived peptides humanin and MOTS-C and Nrf2 in chronic kidney disease. Am. J. Physiol. Renal Physiol. 2019, 317, F1122–F1131. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhu, S.; Wang, H.; Wang, L.; Du, T.; Ye, Z.; Zhai, D.; Zhu, Z.; Tian, X.; Lu, Z.; et al. MOTS-c inhibits Osteolysis in the Mouse Calvaria by affecting osteocyte-osteoclast crosstalk and inhibiting inflammation. Pharmacol. Res. 2019, 147, 104381. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Han, X.; Zhou, D.; Zhong, J.; Liu, L.; Wang, Y.; Ni, J.; Shen, X.; Liang, C.; Fang, H. BIG1 mediates sepsis-induced lung injury by modulating lipid raft-dependent macrophage inflammatory responses. Acta Biochim. Biophys. Sin. 2021, 53, 1088–1097. [Google Scholar] [CrossRef]
- Weng, F.B.; Zhu, L.F.; Zhou, J.X.; Shan, Y.; Tian, Z.G.; Yang, L.W. MOTS-c accelerates bone fracture healing by stimulating osteogenesis of bone marrow mesenchymal stem cells via positively regulating FOXF1 to activate the TGF-beta pathway. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2459. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Q.; Shan, Y.; Ye, F.; Zhang, X.; Cheng, J.; Wang, X.; Zhao, Y.; Dan, G.; Chen, M.; et al. The Mitochondrial-Derived Peptide (MOTS-c) Interacted with Nrf2 to Defend the Antioxidant System to Protect Dopaminergic Neurons Against Rotenone Exposure. Mol. Neurobiol. 2023, 60, 5915–5930. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ren, K.; Jiang, T.; Zhao, G.J. MOTS-c attenuates endothelial dysfunction via suppressing the MAPK/NF-kappaB pathway. Int. J. Cardiol. 2018, 268, 40. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, J.; Feng, M.; Peng, J.; Du, X.; Chu, H.; Chen, X. The Mitochondrial-Derived Peptide MOTS-c Attenuates Oxidative Stress Injury and the Inflammatory Response of H9c2 Cells Through the Nrf2/ARE and NF-kappaB Pathways. Cardiovasc. Eng. Technol. 2022, 13, 651–661. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Criteria | Score = 0 | Score = 1 | Score = 2 | Score = 3 | Score = 4 |
|---|---|---|---|---|---|
| Inflammation | Absent | Mild | Moderate | Severe | Very severe |
| Edema and hemorrhage | None | <10% | 10–30% | 30–50% | >50% |
| Alveolar septal thickening | <15 μm | 15–30 μm | 30–45 μm | 45–60 μm | >60 μm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Huang, J.; Zhang, Y.; Jiang, F.; Li, S.; He, S.; Sun, J.; Chen, D.; Tong, Y.; Pang, Q.; et al. The Mitochondrial-Derived Peptide MOTS-c Alleviates Radiation Pneumonitis via an Nrf2-Dependent Mechanism. Antioxidants 2024, 13, 613. https://doi.org/10.3390/antiox13050613
Zhang Y, Huang J, Zhang Y, Jiang F, Li S, He S, Sun J, Chen D, Tong Y, Pang Q, et al. The Mitochondrial-Derived Peptide MOTS-c Alleviates Radiation Pneumonitis via an Nrf2-Dependent Mechanism. Antioxidants. 2024; 13(5):613. https://doi.org/10.3390/antiox13050613
Chicago/Turabian StyleZhang, Yanli, Jianfeng Huang, Yaru Zhang, Fengjuan Jiang, Shengpeng Li, Shuai He, Jiaojiao Sun, Dan Chen, Ying Tong, Qingfeng Pang, and et al. 2024. "The Mitochondrial-Derived Peptide MOTS-c Alleviates Radiation Pneumonitis via an Nrf2-Dependent Mechanism" Antioxidants 13, no. 5: 613. https://doi.org/10.3390/antiox13050613
APA StyleZhang, Y., Huang, J., Zhang, Y., Jiang, F., Li, S., He, S., Sun, J., Chen, D., Tong, Y., Pang, Q., & Wu, Y. (2024). The Mitochondrial-Derived Peptide MOTS-c Alleviates Radiation Pneumonitis via an Nrf2-Dependent Mechanism. Antioxidants, 13(5), 613. https://doi.org/10.3390/antiox13050613