
Vitamin E Attenuates Red-Light-Mediated Vasodilation: The Benefits of a Mild Oxidative Stress
 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Pressure Myography
2.3. Exosome Tracking
2.4. Cell Culture
2.5. Immunofluorescence
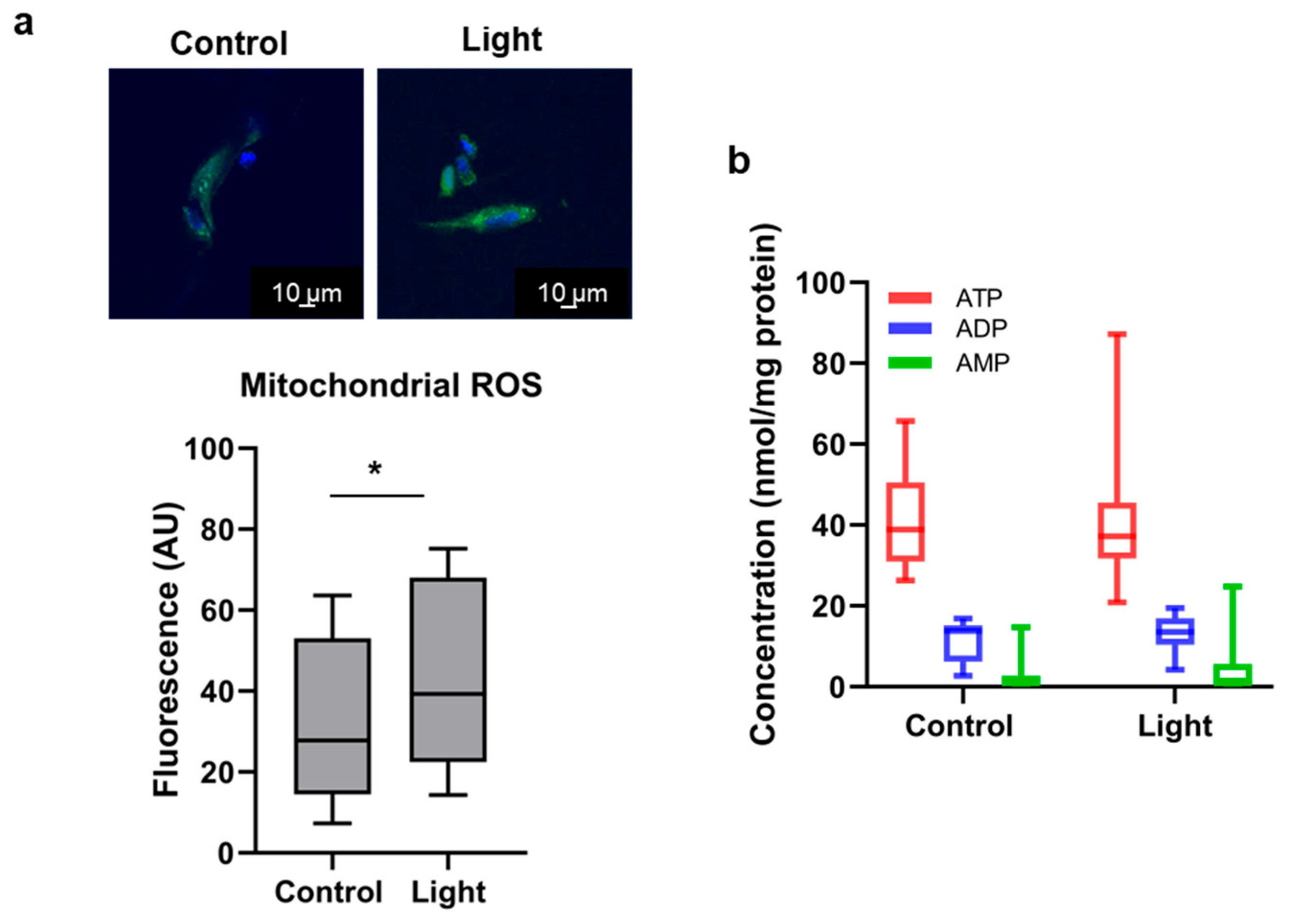
2.6. Adenine Nucleotide Analysis
2.7. Statistics
3. Results
3.1. Effect of Trolox and Catalase on Red-Light-Mediated Dilation of Ex Vivo Blood Vessels
3.1.1. Trolox
3.1.2. Catalase
3.2. Effect of 670 nm Light on Reactive Oxygen Species (ROS) Levels
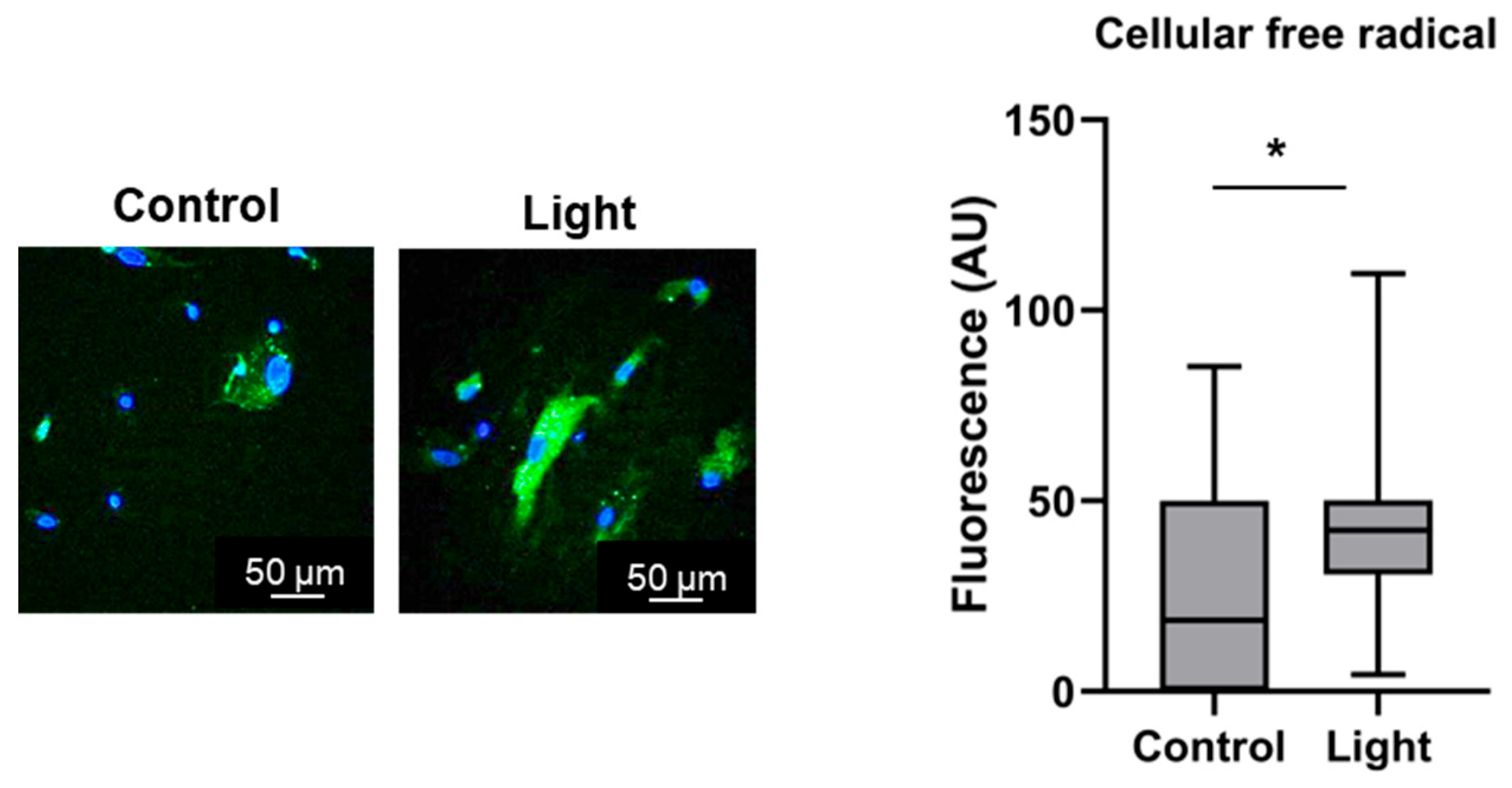
3.2.1. Microsomal and Cellular Free Radicals
3.2.2. Mitochondrial ROS
3.3. Effect of Red-Light-Induced Oxidative Stress on Exosome Formation and Release
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mester, E.; Ludany, G.; Selyei, M.; Szende, B.; Total, G.J. The stimulating effect of low power laser rays on biological systems. Laser Rev. 1968, 1, 3. [Google Scholar]
- Mester, E.; Szende, B.; Gartner, P. The effect of laser beams on the growth of hair in mice. Radiobiol. Radiother. 1968, 9, 621–626. [Google Scholar]
- Hamblin, M.R. Photobiomodulation or low-level laser therapy. J. Biophotonics 2016, 9, 1122–1124. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Dai, T.; Sharma, S.K.; Huang, Y.Y.; Carroll, J.D.; Hamblin, M.R. The nuts and bolts of low-level laser (light) therapy. Ann. Biomed. Eng. 2012, 40, 516–533. [Google Scholar] [CrossRef] [PubMed]
- Colombo, E.; Signore, A.; Aicardi, S.; Zekiy, A.; Utyuzh, A.; Benedicenti, S.; Amaroli, A. Experimemntal and clinical applications of red and near-infrared photobiomodulation and endothelial dysfunction: A review. Biomedicines 2021, 9, 274. [Google Scholar] [CrossRef] [PubMed]
- Karu, T. Primary and secondary mechanisms of action of visible to near-IR radiation on cells. J. Photochem. Photobiol. 1999, 49, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Keszler, A.; Lindemer, B.; Hogg, N.; Weihrauch, D.; Lohr, N.L. Wavelength-dependence of vasodilation and NO release from S-nitrosothiols and dinitrosys iron complexes by far red/near infrared light. Arch. Biochem. Biophys. 2018, 649, 47–52. [Google Scholar] [CrossRef]
- Wajih, N.; Alipour, E.; Rigal, F.; Zhu, J.; Perlegas, A.; Caudell, D.L.; Kim-Shapiro, D. Effects of nitrite and far-red light on coagulation. Nitric Oxide 2021, 107, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, S.; Roessing, M.; Morita, A.; Kopp, T.; Ogawa, E.; Katagiri, W.; Feil, S.; Huang, P.L.; Atochin, D.N.; Kashiwagi, S. Near-infrared II photobiomodulation augments nitric oxide bioavailability via phosphorylation of endothelial nitric oxide synthase. FASEB J. 2022, 36, e22490. [Google Scholar] [CrossRef]
- George, S.; Hamblin, M.R.; Abrahamse, H. Effect of red light and near infrared laser on the generation of reactive oxygen species in primary dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2018, 188, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Kim, H.E.; Cho, H.; Shi, S.; Kim, B.; Kim, O. Red light-emitting diode irradiation regulates oxidative stress and inflammation through SPHK1/NF-κB activation in human keratinocytes. J. Photochem. Photobiol. B Biol. 2018, 186, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Lim, S.T.S. LED light-induced ROS differentially regulates focal adhesion of kinase activity in HaCaT cell viability. Curr. Issues Mol. Biol. 2022, 44, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Mostafavinia, A.; Ahmadi, H.; Amini, A.; Roudafshani, Z.; Hamblin, M.R.; Chien, S.; Bayat, M. The effect of photobiomodulation therapy on antioxidants and oxidative stress profiles of adipose derived mesenchymal stem cells in diabetic rats. Spectrochim. Acta Part A Mol. Biomol. Spect. 2021, 262, 120157. [Google Scholar] [CrossRef] [PubMed]
- De Marchi, T.; Ferlito, J.V.; Ferlito, M.V.; Salvador, M.; Leal-Junior, E.C.P. Can photobiomodulation therapy (PBMT) minimize exercise-induced oxidative stress? A systematic review and meta-analysis. Antioxidants 2022, 11, 1671. [Google Scholar] [CrossRef] [PubMed]
- Keszler, A.; Lindemer, B.; Broeckel, G.; Weihrauch, D.; Gao, Y.; Lohr, N.L. In vivo characterization of a red light-activated vasodilation: A photobiomodulation study. Front. Physiol. 2022, 13, 880158. [Google Scholar] [CrossRef] [PubMed]
- Keszler, A.; Lindemer, B.; Weihrauch, D.; Jones, D.; Hogg, N.; Lohr, N.L. Red/near infrared light stimulates release of an endothelium dependent vasodilator and rescues vascular disfunction in a diabetes model. Free Radic. Biol. Med. 2017, 113, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Weihrauch, D.; Keszler, A.; Lindemer, B.; Krolikowski, J.; Lohr, N.L. Red light stimulates vasodilation through extracellular vesicle trafficking. J. Photochem. Photobiol. B Biol. 2021, 220, 112212. [Google Scholar] [CrossRef]
- Perez, J.; Hill, B.G.; Benavides, G.A.; Dranka, B.P.; Darley-Usmar, V.M. Role of cellular bioenergetics in smooth muscle cell proliferation induced by platelet-derived growth factor. Biochem. J. 2010, 428, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Broniowska, K.A.; Diers, A.R.; Corbett, J.A.; Hogg, N. Nitric oxide effect on naphtoquinone toxicity in endothelial cells: Role of bioenergetic dysfunction and PARP activation. Biochemistry 2013, 52, 4364–4372. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Levine, M. Vitamin C physiology: The known and unknown and Goldilocks. Oral Dis. 2016, 22, 463–493. [Google Scholar] [CrossRef] [PubMed]
- Kashiba-Iwatsuki, M.; Yamaguchi, M.; Inoue, M. Role of ascorbic acid in the metabolism of S-nitroso-glutathione. FEBS Lett. 1996, 389, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Keszler, A.; Lindemer, B.; Hogg, N.; Lohr, N.L. Ascorbate attenuates red light mediated vasodilation: Potential role of S-nitrosothiols. Redox Biol. 2019, 20, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Elia, P.; Azoulay, A.; Zeiri, Y. On the efficiency of water soluble antioxidants. Ultrason. Sonochem. 2012, 19, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Muller, K.; Korth, H.G.; de Groot, H.; Kirsch, M. Reaction of Vitamin E compounds with N-nitrosated tryptophan derivatives and its analytical use. Chem. Eur. J. 2007, 13, 7532–7542. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Kehm, R.; Baldensperger, T.; Raupbach, J.; Hohn, A. Protein oxidation—Formation mechanisms, detection and relevance as biomarkers in human diseases. Redox Biol. 2021, 42, 101901. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.P. Imaging free radicals in organelles, cells, tissues, and in vivo with immune-spin trapping. Redox Biol. 2016, 8, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Balaban, R.S.; Nemoto, S.; Fionkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Munro, D.; Treberg, J.R. A radical shift in perspective: Mitochondria as regulators of reactive oxygen species. J. Exp. Biol. 2017, 220, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, R.; Chen, Y.; Wang, M.; Du, J. Crosstalk between oxidative stress and exosomes. Ox. Med. Cell. Longev. 2022, 2022, 3553617. [Google Scholar] [CrossRef]
- Chiaradia, E.; Tancini, B.; Emiliani, C.; Delo, F.; Pellegrino, R.M.; Tognoloni, A.; Urbanelli, L.; Buratta, S. Extracellular vesicles under oxidative stress condition: Biological properties and physiological roles. Cells 2021, 10, 1763. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Rice, G.E.; Scholz-Romero, K.; Sweeney, E.; Peiris, H.; Kobayashi, M.; Duncombe, G.; Mitchell, M.D.; Salomon, C. The effect of glucose on the release and bioactivity of exosomes from first trimester trophoblast cells. J. Clin. Endocrin. Metabol. 2015, 100, E1280–E1288. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, N.; Chisci, E.; Giovannoni, R. The role of hydrogen peroxide in redox-dependent signaling: Homeostatic and pathological responses in mammalian cells. Cells 2018, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Kubo, K.; Waguri, S.; Yabashi, A.; Shin, H.W.; Katoh, Y.; Nakayama, K. Rab11 regulates exocytosis of recycling vesicles at the plasma membrane. J. Cell Sci. 2012, 125, 4049–4057. [Google Scholar] [CrossRef] [PubMed]
- Neuman, S.D.; Lee, A.R.; Selegue, J.E.; Cavanagh, A.T.; Bashirullah, A. A novel function for Rab1 and Rab11 during secretory granule maturation. J. Cell Sci. 2021, 134, 259037. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Lakkaraju, A. Early endosome morphology in health and disease. Adv. Exp. Med. Biol. 2018, 1074, 335–343. [Google Scholar] [PubMed]
- Weihrauch, D.; Keszler, A.; Broeckel, G.; Aranda, E.; Lindemer, B.; Lohr, N.L. Red light mediates the exocytosis of vasodilatory vesicles from cultured endothelial cells: A cellular, and ex vivo murine model. Photochem. Photobiol. Sci. 2024, in press. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nature Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Gruenberg, J. Life in the lumen: The multivesicular endosome. Traffic 2020, 21, 76–93. [Google Scholar] [CrossRef]
- Atienzar-Aroca, S.; Flores-Bellver, M.; Serrano-Heras, G.; Martinez-Gil, N.; Barcia, J.M.; Aparicio, S.; Perez-Cremades, D.; Garcia-Verdugo, J.M.; Diaz-llopis, M.; Romero, F.J.; et al. Oxidative stress in retinal pigment epithelium cells increases exosome secretion and ptomotes angiogenesis in endothelial cells. J. Cell. Mol. Med. 2016, 20, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.C.H.; Huang, Y.Y.; Arany, P.R.; Hamblin, M.R. Role of reactive oxygen species in low level light therapy. In Proceedings of the SPIE BIOS, San Jose, CA, USA, 24–29 January 2009; Volume 7165. [Google Scholar]
- Day, R.M.; Suzuki, Y.J.; Fanburg, B.L. Regulation of glutathione by oxidative stress in bovine pulmonary artery endothelial cells. Antiox. Redox. Signal. 2004, 5, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.; Ridgeway, T.D.; Higbee, R.G.; Howard, E.W.; Lucroy, M.D. Effect of wavelength on low-intensity laser irradiation-stimulated cell proliferation in vitro. Lasers Surg. Med. 2005, 36, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Ash, C.; Dubec, M.; Donne, K.; Bashford, T. Effect of the wavelength and beam width on penetration in light-tissue interaction using computational methods. Lasers Med. Sci. 2017, 32, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Nacharaju, P.; Friedman, A.; Friedman, J.M. Nitric oxide generating/releasing materials. Future Sci. OA 2015, 1, FSO54. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keszler, A.; Weihrauch, D.; Lindemer, B.; Broeckel, G.; Lohr, N.L. Vitamin E Attenuates Red-Light-Mediated Vasodilation: The Benefits of a Mild Oxidative Stress. Antioxidants 2024, 13, 668. https://doi.org/10.3390/antiox13060668
Keszler A, Weihrauch D, Lindemer B, Broeckel G, Lohr NL. Vitamin E Attenuates Red-Light-Mediated Vasodilation: The Benefits of a Mild Oxidative Stress. Antioxidants. 2024; 13(6):668. https://doi.org/10.3390/antiox13060668
Chicago/Turabian StyleKeszler, Agnes, Dorothee Weihrauch, Brian Lindemer, Grant Broeckel, and Nicole L. Lohr. 2024. "Vitamin E Attenuates Red-Light-Mediated Vasodilation: The Benefits of a Mild Oxidative Stress" Antioxidants 13, no. 6: 668. https://doi.org/10.3390/antiox13060668