

Chondroitin Sulfate Ameliorates Hypertension in Male Offspring Rat Born to Mothers Fed an Adenine Diet

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. H2S-Generating Enzymes
2.3. NO Parameters
2.4. Plasma Short Chain Fatty Acids (SCFAs)
2.5. 16S rRNA Gene Sequencing and Analysis of Gut Microbiota Composition
2.6. Statistics
3. Results
3.1. Offspring Outcomes
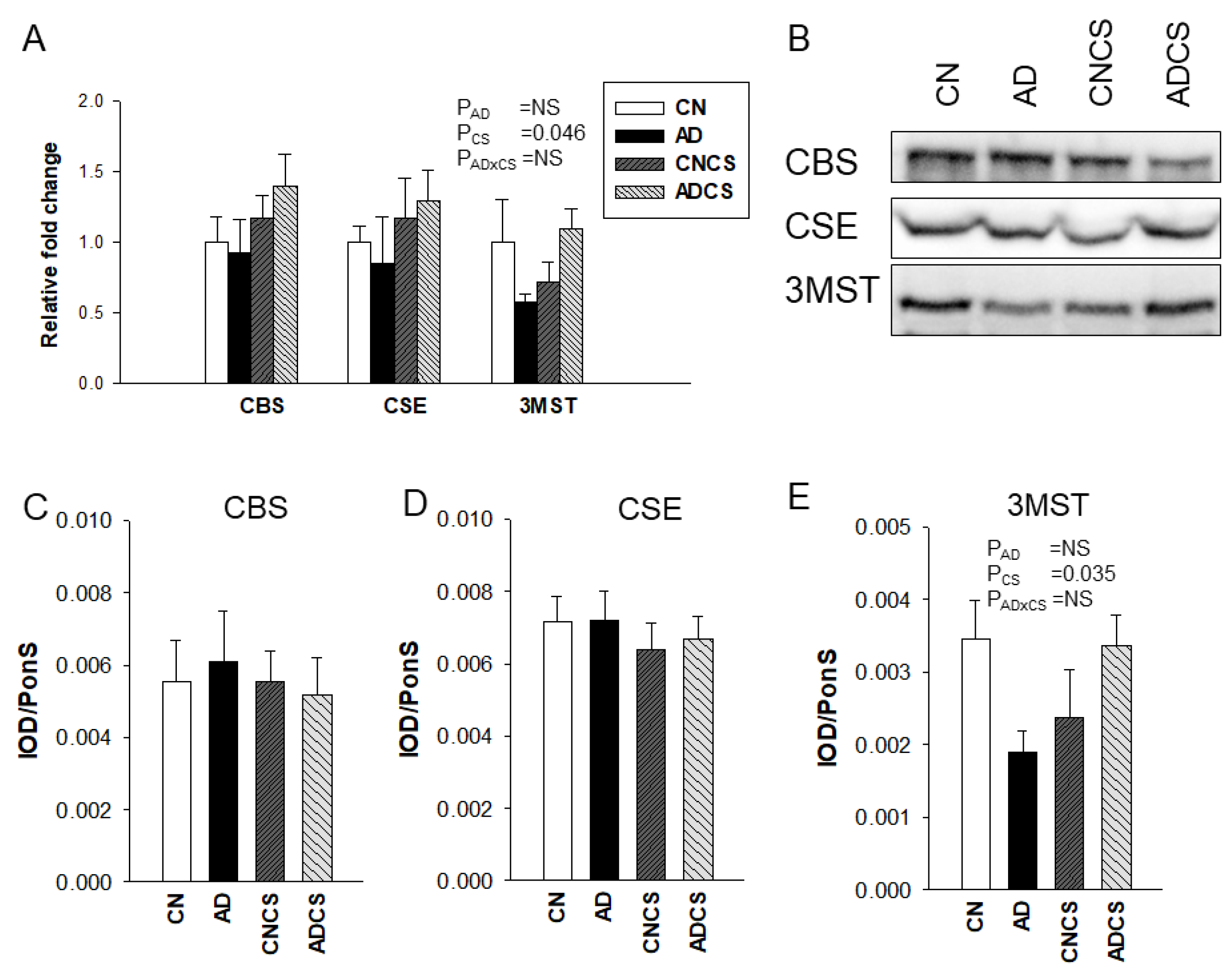
3.2. H2S Pathway
3.3. NO Pathway
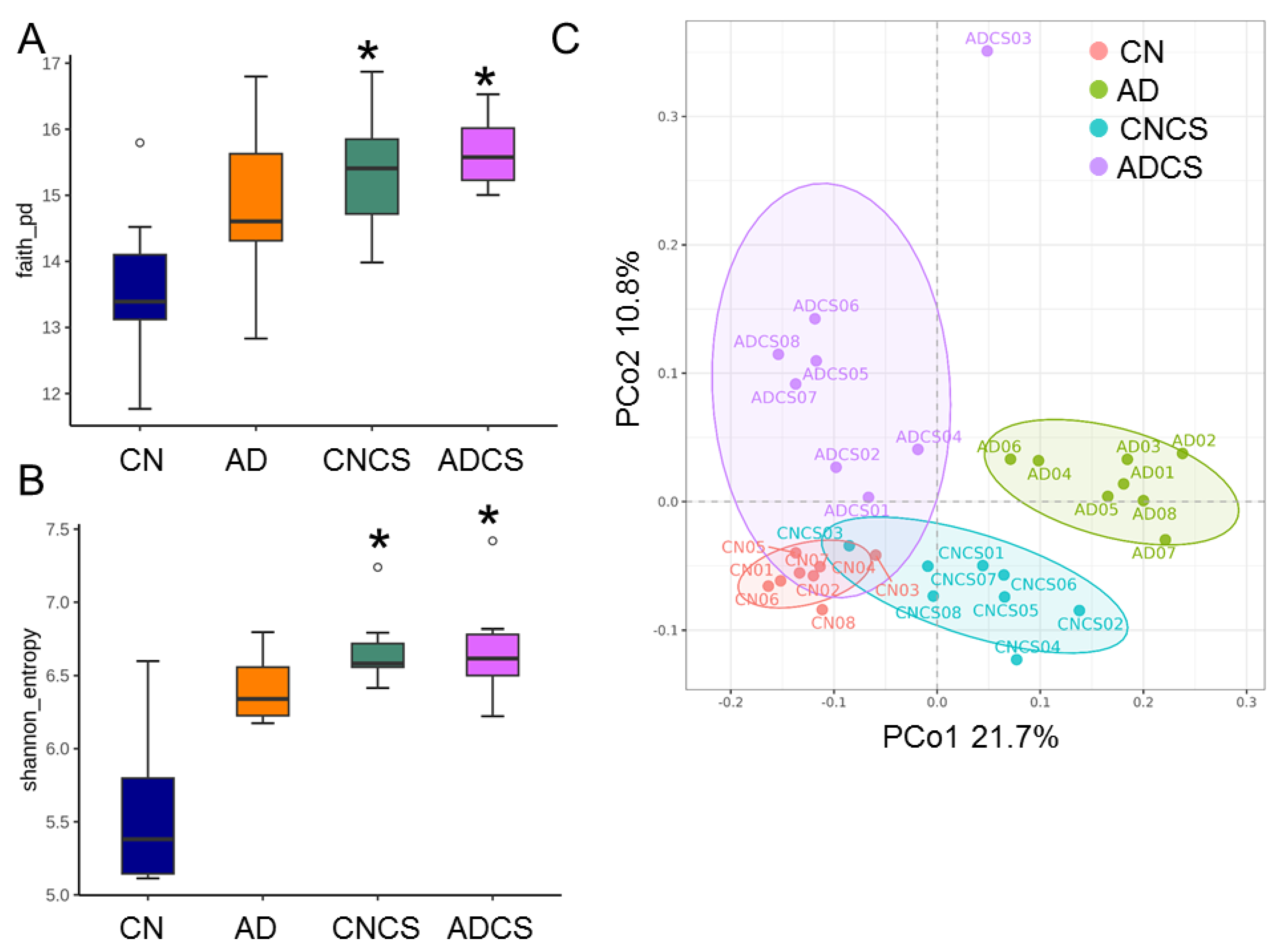
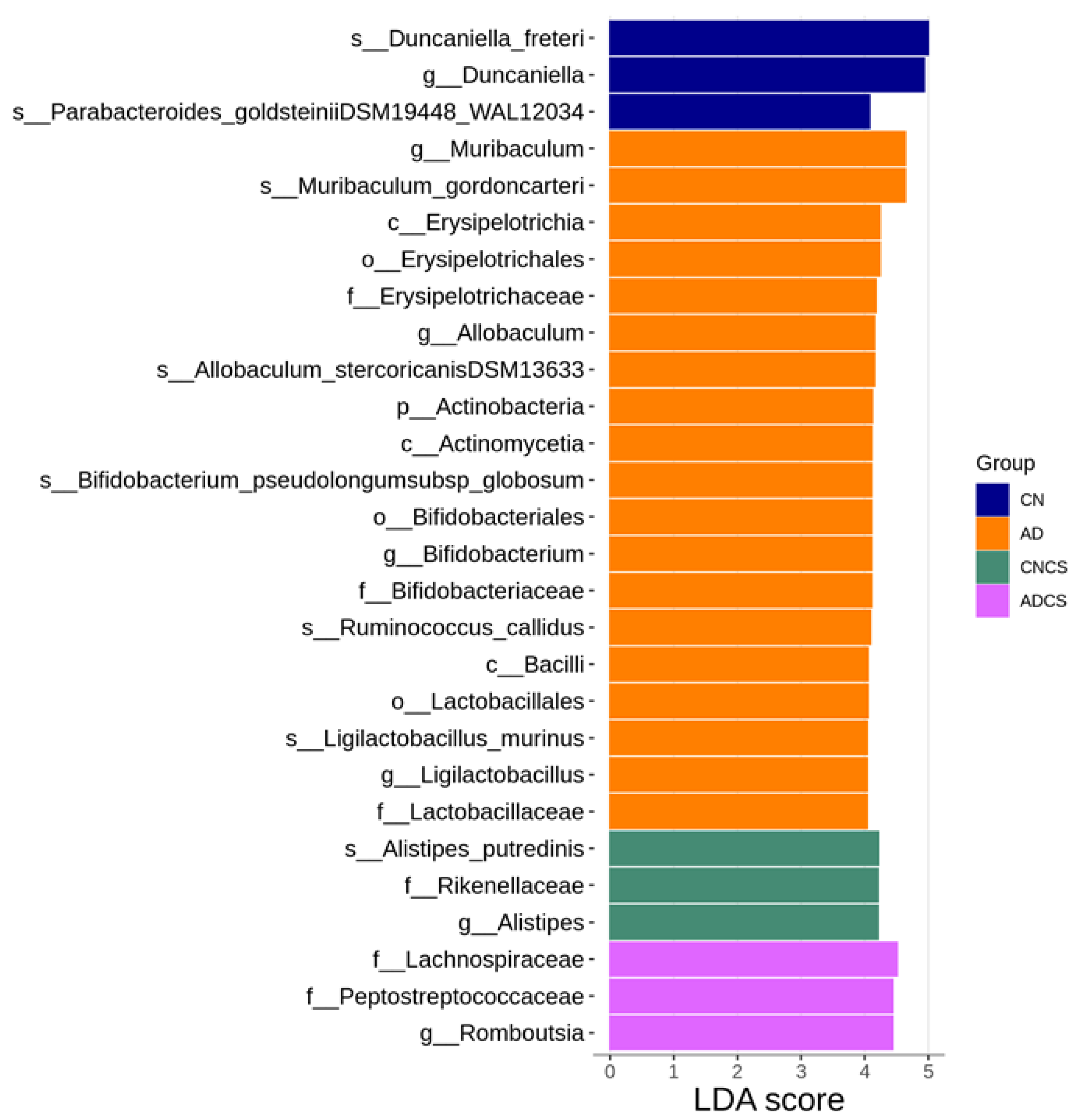
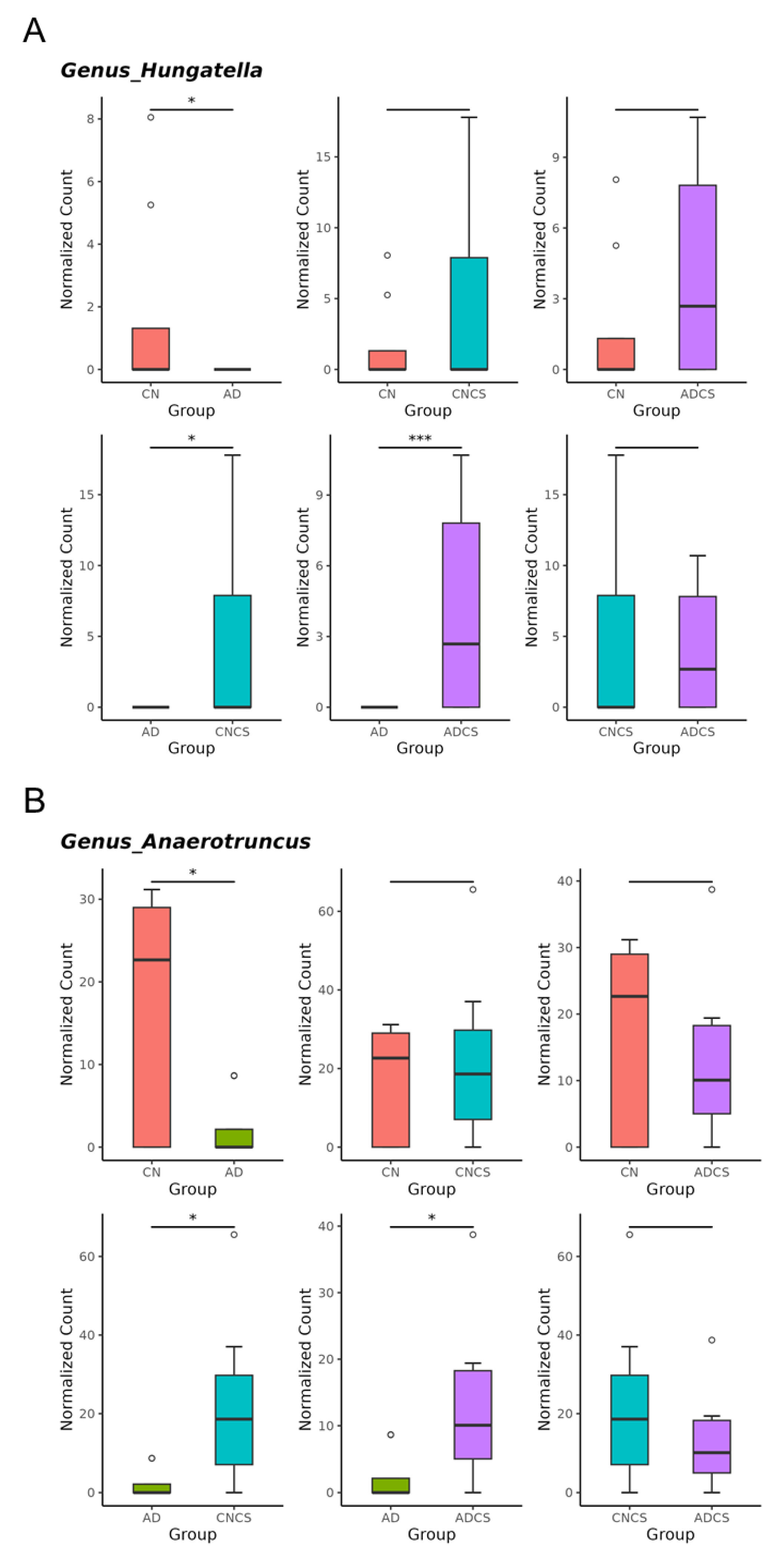
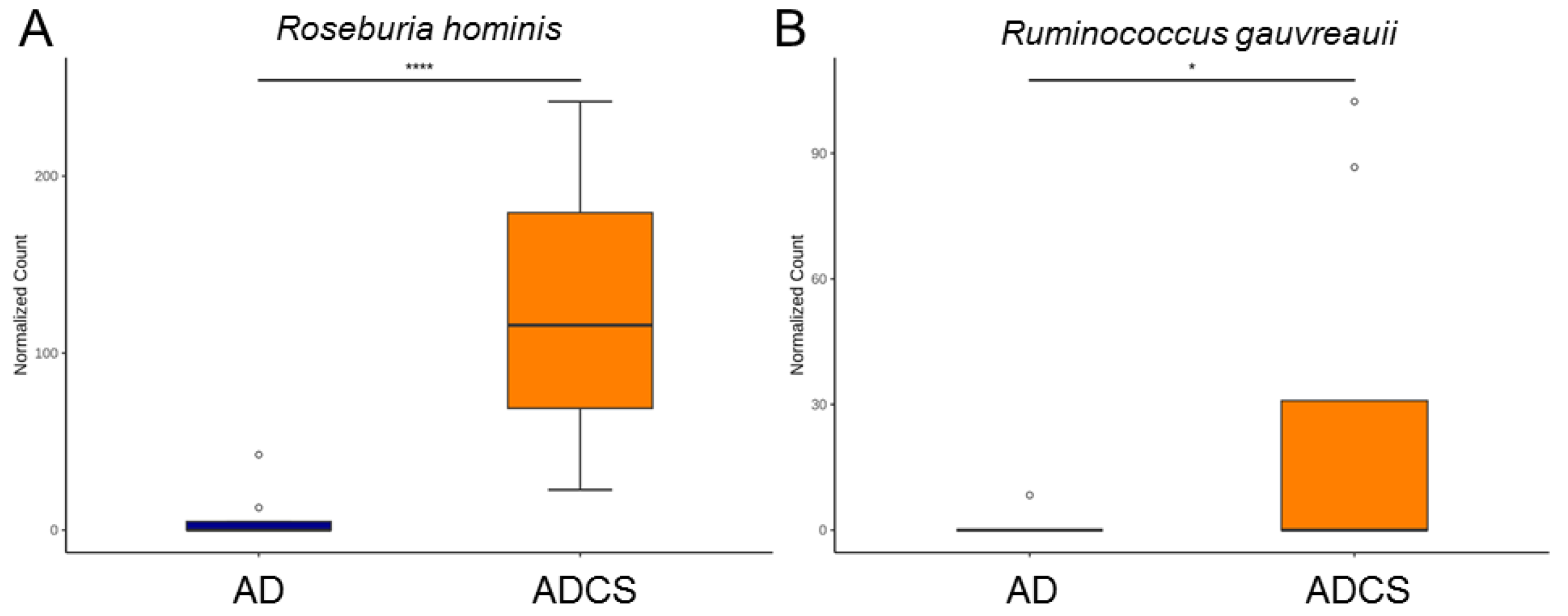
3.4. Differences in Microbiota Composition
3.5. SCFAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munkhaugen, J.; Lydersen, S.; Romundstad, P.R.; Widerøe, T.-E.; Vikse, B.E.; Hallan, S. Kidney function and future risk for adverse pregnancy outcomes: A population-based study from HUNT II, Norway. Nephrol. Dial. Transplant. 2009, 24, 3744–3750. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, G.B.; Alrukhaimi, M.; Liu, Z.H.; Zakharova, E.; Levin, A.; World Kidney Day Steering Committee. What we do and do not know about women and kidney diseases; Questions unanswered and answers unquestioned: Reflection on World Kidney Day and International Woman’s Day. Physiol. Int. 2018, 105, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.P.; Velazquez, M.A.; Eckert, J.J. Embryos, DOHaD and David Barker. J. Dev. Orig. Health Dis. 2015, 6, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Gluckman, P.D. Early developmental conditioning of later health and disease: Physiology or pathophysiology? Physiol. Rev. 2014, 94, 1027–1076. [Google Scholar] [CrossRef] [PubMed]
- Paauw, N.D.; van Rijn, B.B.; Lely, A.T.; Joles, J.A. Pregnancy as a critical window for blood pressure regulation in mother and child: Programming and reprogramming. Acta Physiol. 2017, 219, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Diwan, V.; Brown, L.; Gobe, G.C. Adenine-induced chronic kidney disease in rats. Nephrology 2018, 23, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Yang, H.W.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal Adenine-Induced Chronic Kidney Disease Programs Hypertension in Adult Male Rat Offspring: Implications of Nitric Oxide and Gut Microbiome Derived Metabolites. Int. J. Mol. Sci. 2020, 21, 7237. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Hsu, C.N. Protection by Means of Perinatal Oral Sodium Thiosulfate Administration against Offspring Hypertension in a Rat Model of Maternal Chronic Kidney Disease. Antioxidants 2023, 12, 1344. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Guo, Y.; Wang, K.; Zhang, C.; Ma, Y. A Review of Chondroitin Sulfate’s Preparation, Properties, Functions, and Applications. Molecules 2023, 28, 7093. [Google Scholar] [CrossRef]
- Meng, Z.; Liu, J.; Zhou, N. Efficacy and safety of the combination of glucosamine and chondroitin for knee osteoarthritis: A systematic review and meta-analysis. Arch. Orthop. Trauma Surg. 2023, 143, 409–421. [Google Scholar] [CrossRef]
- Stellavato, A.; Restaino, O.F.; Vassallo, V.; Finamore, R.; Ruosi, C.; Cassese, E.; De Rosa, M.; Schiraldi, C. Comparative Analyses of Pharmaceuticals or Food Supplements Containing Chondroitin Sulfate: Are Their Bioactivities Equivalent? Adv. Ther. 2019, 36, 3221–3237. [Google Scholar] [CrossRef] [PubMed]
- Barthe, L.; Woodley, J.; Lavit, M.; Przybylski, C.; Philibert, C.; Houin, G. In vitro intestinal degradation and absorption of chondroitin sulfate, a glycosaminoglycan drug. Arzneimittel-Forschung 2004, 54, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Shmagel, A.; Demmer, R.; Knights, D.; Butler, M.; Langsetmo, L.; Lane, N.E.; Ensrud, K. The Effects of Glucosamine and Chondroitin Sulfate on Gut Microbial Composition: A Systematic Review of Evidence from Animal and Human Studies. Nutrients 2019, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Pichette, J.; Fynn-Sackey, N.; Gagnon, J. Hydrogen Sulfide and Sulfate Prebiotic Stimulates the Secretion of GLP-1 and Improves Glycemia in Male Mice. Endocrinology 2017, 158, 3416–3425. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gao, Y.; Zhao, Y.; Yang, G.; Wang, C.; Zhao, Z.; Li, S. Chondroitin sulfate stimulates the secretion of H2S by Desulfovibrio to improve insulin sensitivity in NAFLD mice. Int. J. Biol. Macromol. 2022, 213, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Signaling molecules: Hydrogen sulfide and polysulfide. Antioxid. Redox Signal. 2015, 22, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Preventing Developmental Origins of Cardiovascular Disease: Hydrogen Sulfide as a Potential Target? Antioxidants 2021, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Vallières, M.; du Souich, P. Modulation of inflammation by chondroitin sulfate. Osteoarthr. Cartil. 2010, 18, S1–S6. [Google Scholar] [CrossRef] [PubMed]
- Reckelhoff, J.F. Gender differences in the regulation of blood pressure. Hypertension 2001, 37, 1199–1208. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Bode-Böger, S.M.; Scalera, F.; Ignarro, L.J. The L-arginine paradox: Importance of the L-arginine/asymmetrical dimethylarginine ratio. Pharmacol. Ther. 2007, 114, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L. Microbial short-chain fatty acids and blood pressure regulation. Curr. Hypertens. Rep. 2017, 19, 25. [Google Scholar] [CrossRef]
- Regueira-Iglesias, A.; Balsa-Castro, C.; Blanco-Pintos, T.; Tomás, I. Critical review of 16S rRNA gene sequencing workflow in microbiome studies: From primer selection to advanced data analysis. Mol. Oral Microbiol. 2023, 38, 347–399. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Rey, F.E.; Gonzalez, M.D.; Cheng, J.; Wu, M.; Ahern, P.P.; Gordon, J.I. Metabolic niche of a prominent sulfate-reducing human gut bacterium. Proc. Natl. Acad. Sci. USA 2013, 110, 13582–13587. [Google Scholar] [CrossRef]
- Wang, R. Roles of Hydrogen Sulfide in Hypertension Development and Its Complications: What, So What, Now What. Hypertension 2023, 80, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Targeting on Asymmetric Dimethylarginine-Related Nitric Oxide-Reactive Oxygen Species Imbalance to Reprogram the Development of Hypertension. Int. J. Mol. Sci. 2016, 17, 2020. [Google Scholar] [CrossRef]
- Jin, L.; Cui, X.; Cai, J. Study of gut microbiota in over-weighted and obese hypertensive patients. Infect. Dis. Informat. 2018, 31, 320–325. [Google Scholar]
- Guo, Y.; Li, X.; Wang, Z.; Yu, B. Gut Microbiota Dysbiosis in Human Hypertension: A Systematic Review of Observational Studies. Front. Cardiovasc. Med. 2021, 8, 650227. [Google Scholar] [CrossRef] [PubMed]
- Khalesi, S.; Sun, J.; Buys, N.; Jayasinghe, R. Effect of probiotics on blood pressure: A systematic review and meta-analysis of randomized, controlled trials. Hypertension 2014, 64, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liang, W.; Liang, J.; Dou, J.; Guo, F.; Zhang, D.; Xu, Z.; Wang, T. Probiotics: Functional food ingredients with the potential to reduce hypertension. Front. Cell Infect. Microbiol. 2023, 13, 1220877. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Lin, Y.J.; Hou, C.Y.; Tain, Y.L. Maternal Administration of Probiotic or Prebiotic Prevents Male Adult Rat Offspring against Developmental Programming of Hypertension Induced by High Fructose Consumption in Pregnancy and Lactation. Nutrients 2018, 10, 1229. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Li, Y.; Chen, M.; Xue, L.; Wang, J.; Ding, Y.; Gu, Q.; Zhang, J.; Yang, R.; Zhao, H.; et al. Effects of probiotics on hypertension. Appl. Microbiol. Biotechnol. 2023, 107, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.L.; Yen, G.C.; Huang, S.C.; Chen, S.C.; Hsu, C.L. The next generation beneficial actions of novel probiotics as potential therapeutic targets and prediction tool for metabolic diseases. J. Food Drug Anal. 2022, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Singh, P.; Nataraj, B.H.; Kokkiligadda, A.; Naithani, H.; Azmal Ali, S.; Behare, P.V.; Nagpal, R. Fostering next-generation probiotics in human gut by targeted dietary modulation: An emerging perspective. Food Res. Int. 2021, 150, 110716. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.Y.; Liao, N.B.; Zhang, Y.; Ye, X.Q.; Li, S.; Hu, Y.Q.; Liu, D.H.; Linhardt, R.J.; Wang, X.; Chen, S.G. In vitro fermentation behaviors of fucosylated chondroitin sulfate from Pearsonothuria graeffei by human gut microflora. Int. J. Biol. Macromol. 2017, 102, 1195–1201. [Google Scholar] [CrossRef]
- Tomasova, L.; Konopelski, P.; Ufnal, M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules 2016, 21, 1558. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | CN | AD | CNCS | ADCS | p Value | ||
|---|---|---|---|---|---|---|---|
| AD | CS | AD × CS | |||||
| Body weight (BW) (g) | 415 ± 18 | 415 ± 14 | 351 ± 7 *# | 426 ± 9 | 0.007 | 0.046 | 0.007 |
| Left kidney weight (LKW) (g) | 1.83 ± 0.08 | 1.94 ± 0.07 | 1.71 ± 0.06 # | 1.97 ± 0.08 | 0.014 | NS | NS |
| Combined kidney weight (CKW) (g) | 3.68 ± 0.16 | 3.94 ± 0.13 | 3.41 ± 0.1 # | 4.02 ± 0.13 | 0.003 | NS | NS |
| LKW-to-BW ratio (g/kg) | 0.44 ± 0.01 | 0.47 ± 0.01 | 0.49 ± 0.01 | 0.47 ± 0.03 | NS | NS | NS |
| CKW-to-BW ratio (g/kg) | 0.89 ± 0.01 | 0.95 ± 0.02 | 0.97 ± 0.02 | 0.95 ± 0.05 | NS | NS | NS |
| Systolic BP (mmHg) | 133 ± 1 | 149 ± 2 * | 135 ± 1 # | 137 ± 1 # | <0.001 | <0.001 | <0.001 |
| Diastolic BP (mmHg) | 88 ± 1 | 91 ± 3 | 88 ± 2 | 89 ± 2 | NS | NS | NS |
| Creatinine (μM) | 16.5 ± 0.5 | 18.8 ± 0.8 | 16.2 ± 0.8 | 18.8 ± 0.7 | 0.002 | NS | NS |
| Groups | CN | AD | CNCS | ADCS | p Value | ||
|---|---|---|---|---|---|---|---|
| AD | CS | AD × CS | |||||
| L-citrulline (μM) | 54.3 ± 3.3 | 43.0 ± 2.5 | 48.0 ± 1.7 | 46.1 ± 3.2 | 0.024 | NS | NS |
| L-arginine (μM) | 392.5 ± 21.6 | 286.2 ± 11.8 | 365.9 ± 18.2 | 302.1 ± 16.3 | <0.001 | NS | NS |
| ADMA (μM) | 2.06 ± 0.1 | 2.19 ± 0.1 | 1.88 ± 0.06 | 2.05 ± 0.17 | NS | NS | NS |
| SDMA (μM) | 2.07 ± 0.16 | 2.49 ± 0.20 | 2.28 ± 0.10 | 2.02 ± 0.08 | NS | NS | 0.035 |
| AAR (μM/μM) | 192.9 ± 12.2 | 132.5 ± 7.9 | 197.1 ± 13.0 | 152.7 ± 11.8 | <0.001 | 0.044 | NS |
| Groups | CN | AD | CNCS | ADCS | p Value | ||
|---|---|---|---|---|---|---|---|
| AD | CS | AD × CS | |||||
| Acetic acid (μM) | 943.9 ± 80.4 | 893.5 ± 42.3 | 1151 ± 49.4 *# | 785.8 ± 43.8 $ | 0.001 | NS | 0.01 |
| Propionic acid (μM) | 4.04 ± 0.78 | 3.8 ± 0.62 | 5.06 ± 0.27 | 4.59 ± 0.56 | NS | NS | NS |
| Isobutyric acid (μM) | 2.12 ± 0.12 | 1.75 ± 0.14 | 2.31 ± 0.09 | 1.9 ± 0.19 | 0.01 | NS | NS |
| Butyric acid (μM) | 10.98 ± 0.49 | 9.29 ± 0.4 | 10.45 ± 0.53 | 8.73 ± 0.31 | 0.001 | NS | NS |
| Isovaleric acid (μM) | 14.42 ± 2 | 7.62 ± 0.47 | 11.99 ± 0.67 | 6.67 ± 0.5 | <0.001 | NS | NS |
| Valeric acid (μM) | 3.88 ± 0.12 | 3.17 ± 0.19 | 4 ± 0.13 | 3.36 ± 0.14 | <0.001 | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tain, Y.-L.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.-F.; Hsu, C.-N. Chondroitin Sulfate Ameliorates Hypertension in Male Offspring Rat Born to Mothers Fed an Adenine Diet. Antioxidants 2024, 13, 944. https://doi.org/10.3390/antiox13080944
Tain Y-L, Hou C-Y, Chang-Chien G-P, Lin S-F, Hsu C-N. Chondroitin Sulfate Ameliorates Hypertension in Male Offspring Rat Born to Mothers Fed an Adenine Diet. Antioxidants. 2024; 13(8):944. https://doi.org/10.3390/antiox13080944
Chicago/Turabian StyleTain, You-Lin, Chih-Yao Hou, Guo-Ping Chang-Chien, Shu-Fen Lin, and Chien-Ning Hsu. 2024. "Chondroitin Sulfate Ameliorates Hypertension in Male Offspring Rat Born to Mothers Fed an Adenine Diet" Antioxidants 13, no. 8: 944. https://doi.org/10.3390/antiox13080944