


The Antioxidant and Anti-Inflammatory Properties of Bee Pollen from Acorn (Quercus acutissima Carr.) and Darae (Actinidia arguta)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Bee Pollen
2.3. Cell Culture and LPS Stimulation
2.4. Cell-Viability Assay
2.5. Intracellular Reactive Oxygen Species (ROS) Assay
2.6. Immunofluorescence (IF) Assay
2.7. Cell-Morphology Assay
2.8. Nitric Oxide (NO) Production Assay
2.9. Prostaglandin E2 (PGE2) Production Assay
2.10. Enzyme-Linked Immunosorbent (ELISA) Assay
2.11. Western Blot Analysis
2.12. Cell Cycle Assay
2.13. Statistical Analysis
3. Results
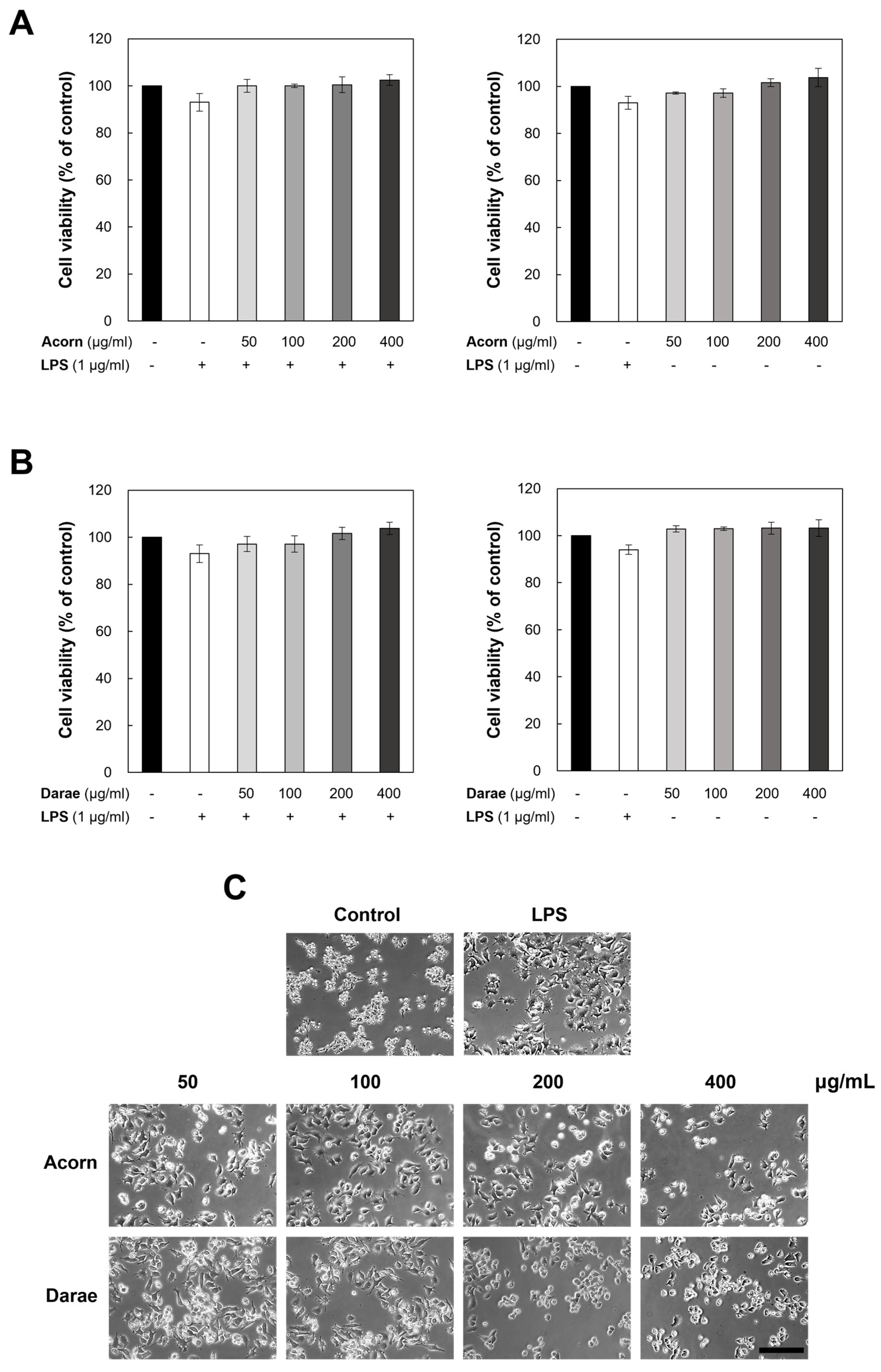
3.1. Effect of EEBP from Acorn and Darae on Cell Viability and Cytotoxicity in RAW 264.7 Macrophages
3.2. Antioxidant Effect of EEBP from Acorn and Darae on ROS Production in RAW 264.7 Macrophages
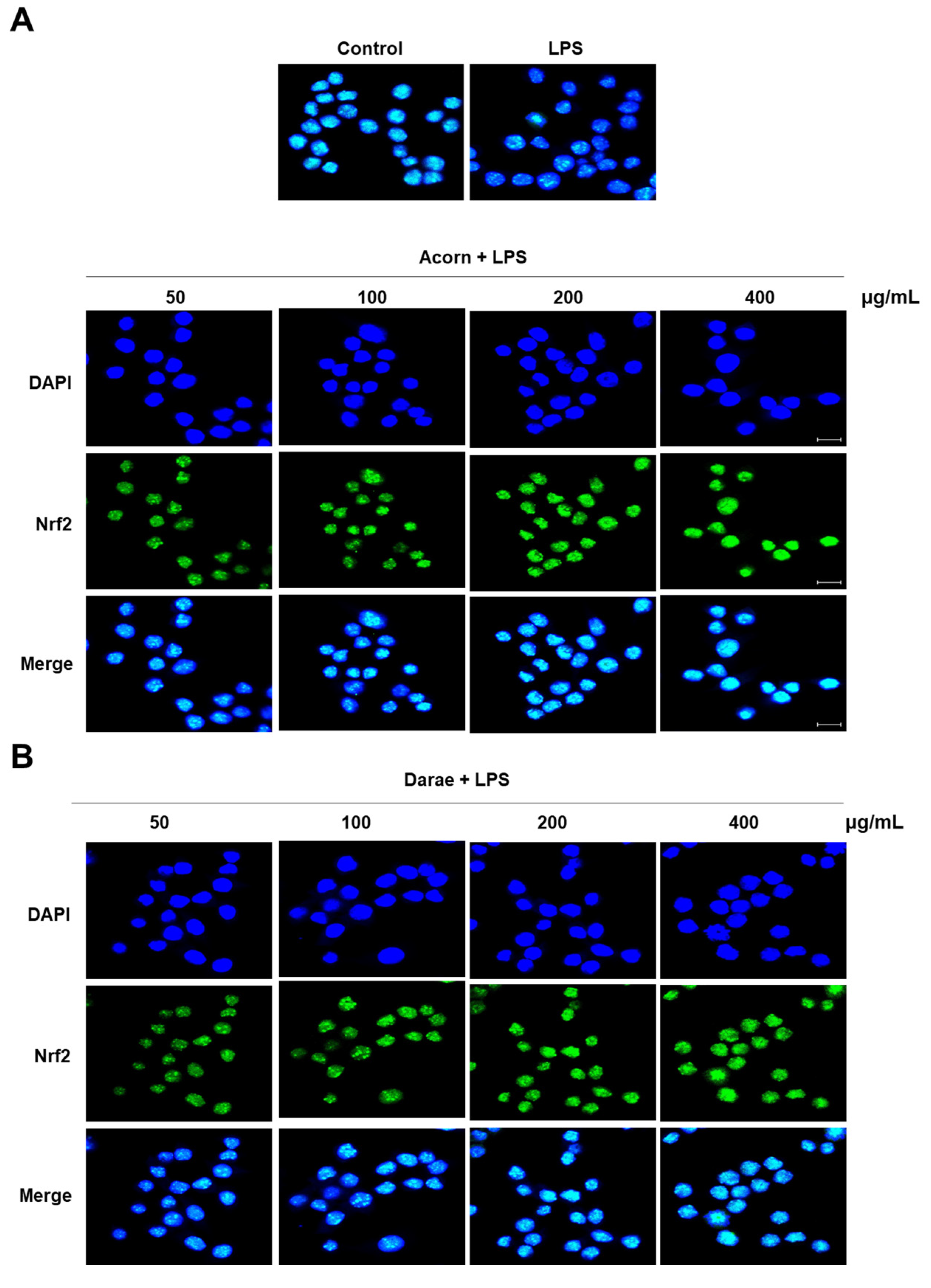
3.3. Effect of EEBP from Acorn and Darae on the Nrf2 Pathway in RAW 264.7 Macrophages
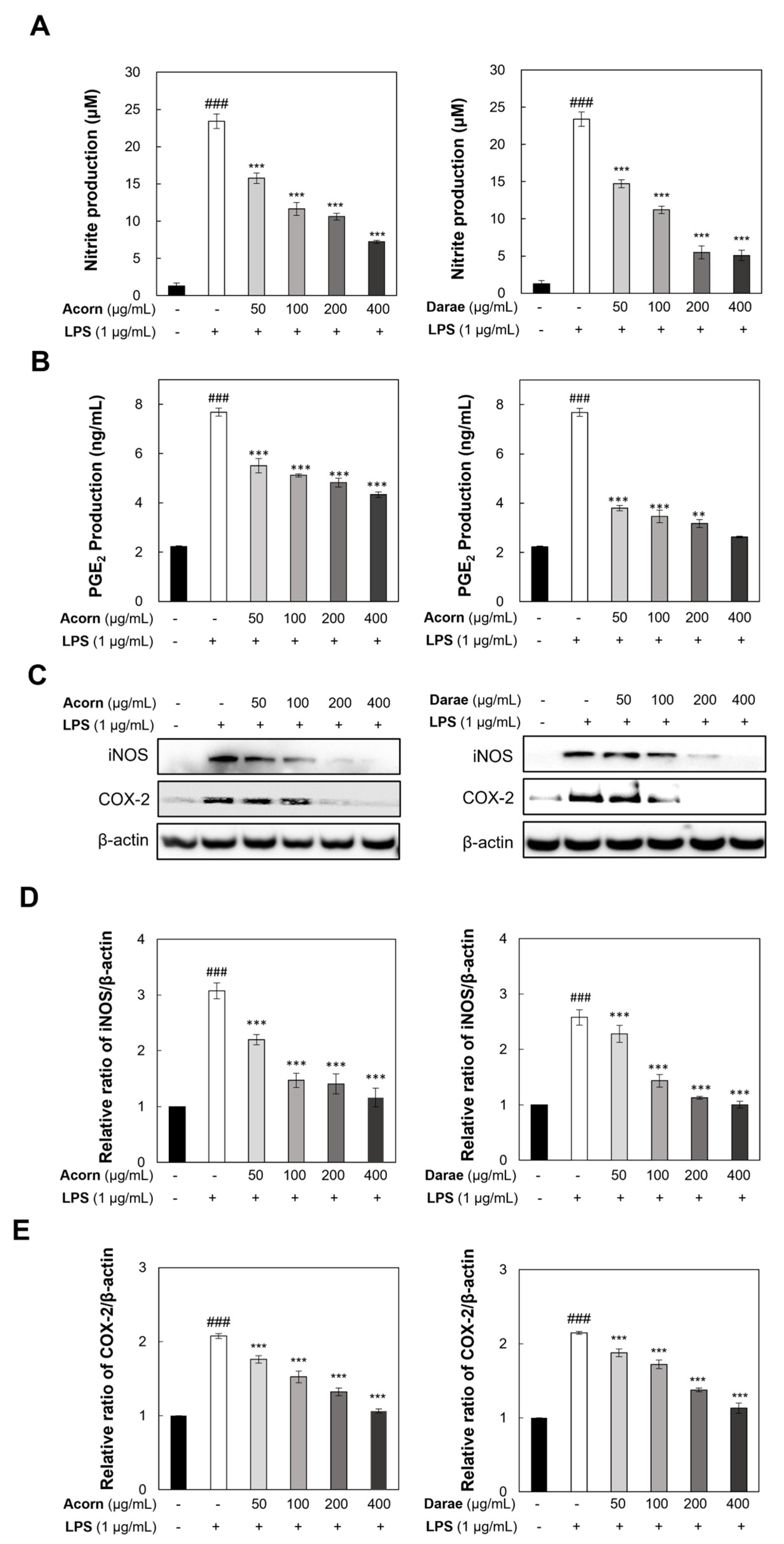
3.4. Effect of EEBP from Acorn and Darae on Pro-Inflammatory Mediators in RAW 264.7 Macrophages
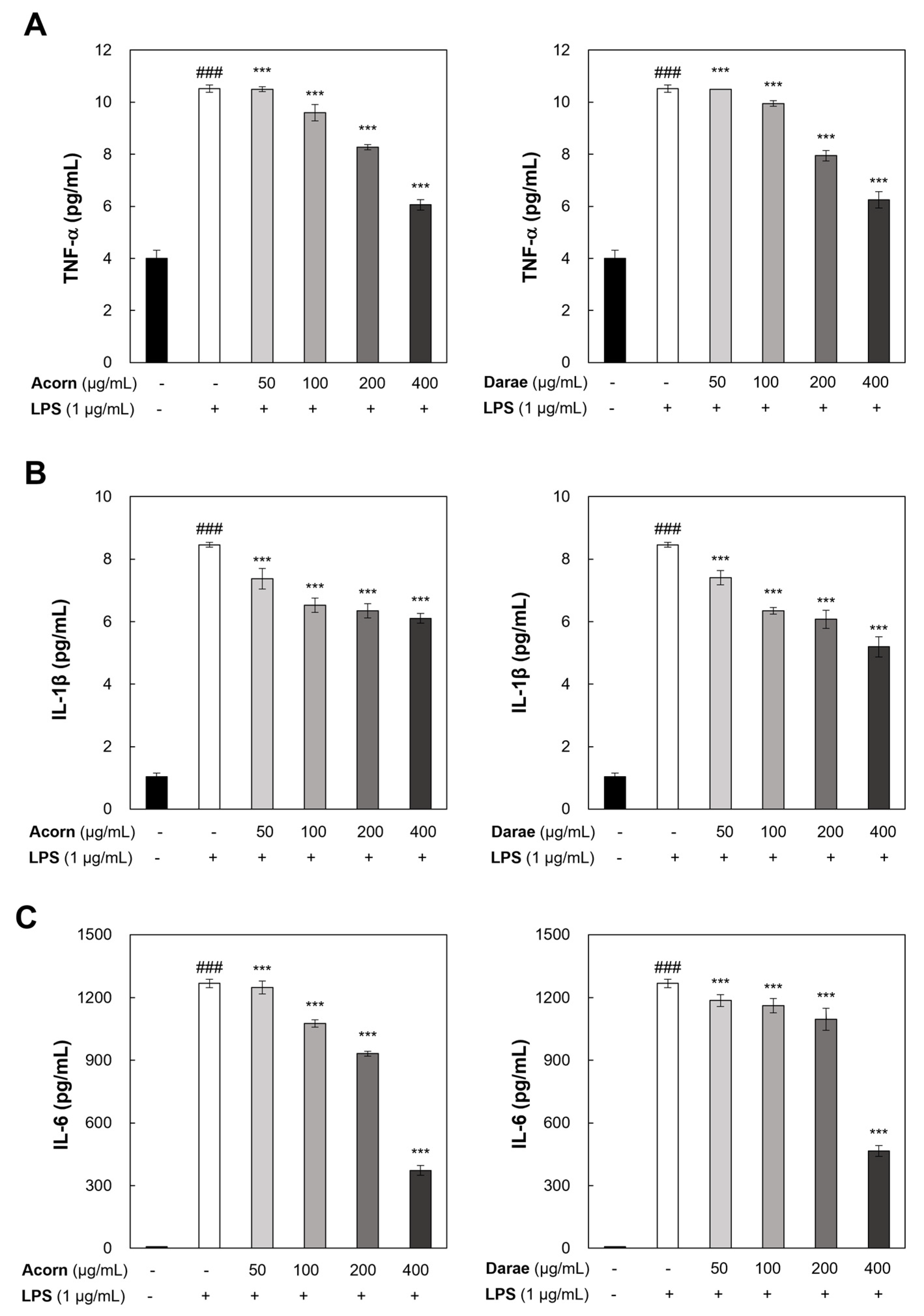
3.5. Effect of EEBP from Acorn and Darae on Pro-Inflammatory Cytokines in RAW 264.7 Macrophages
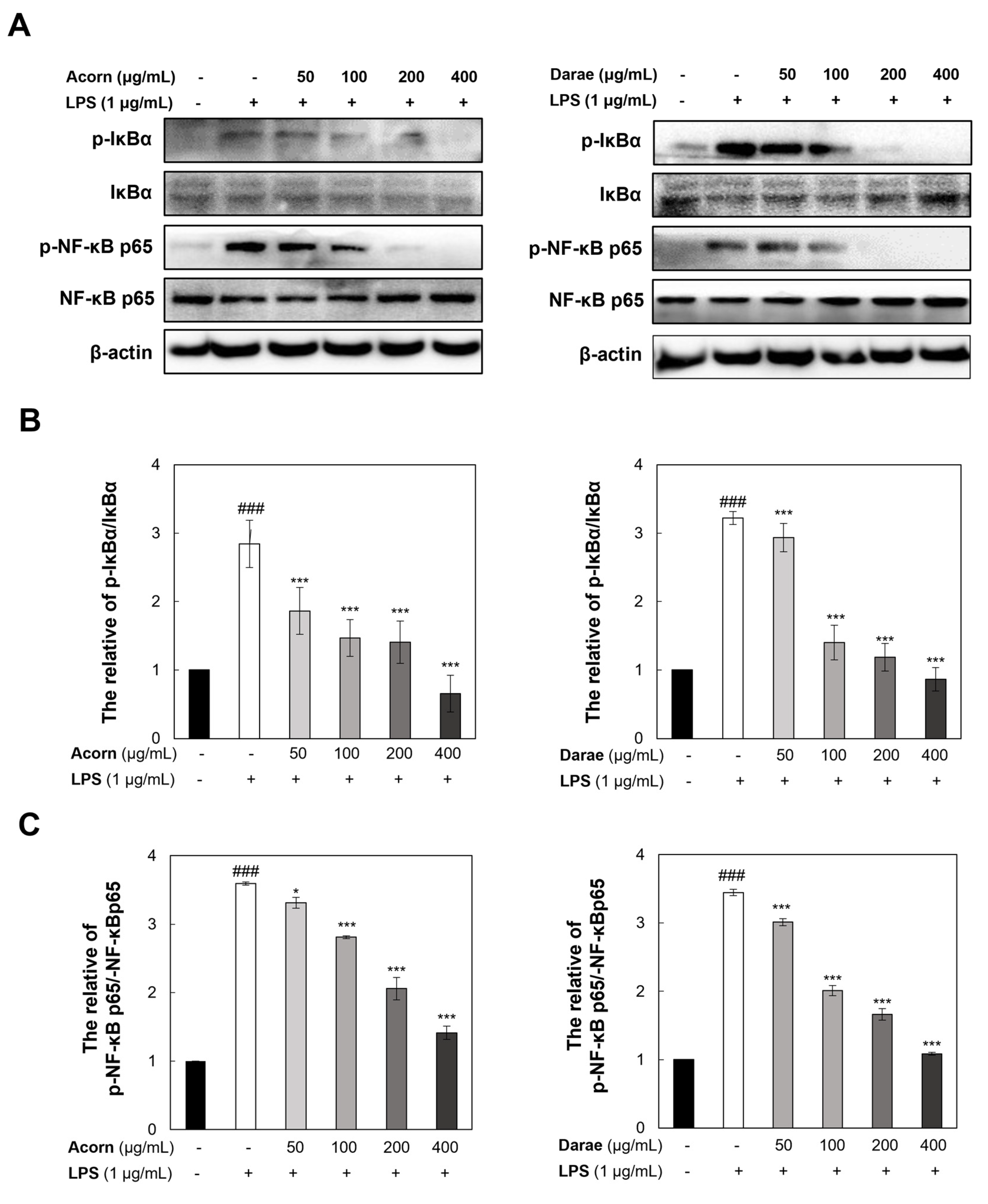
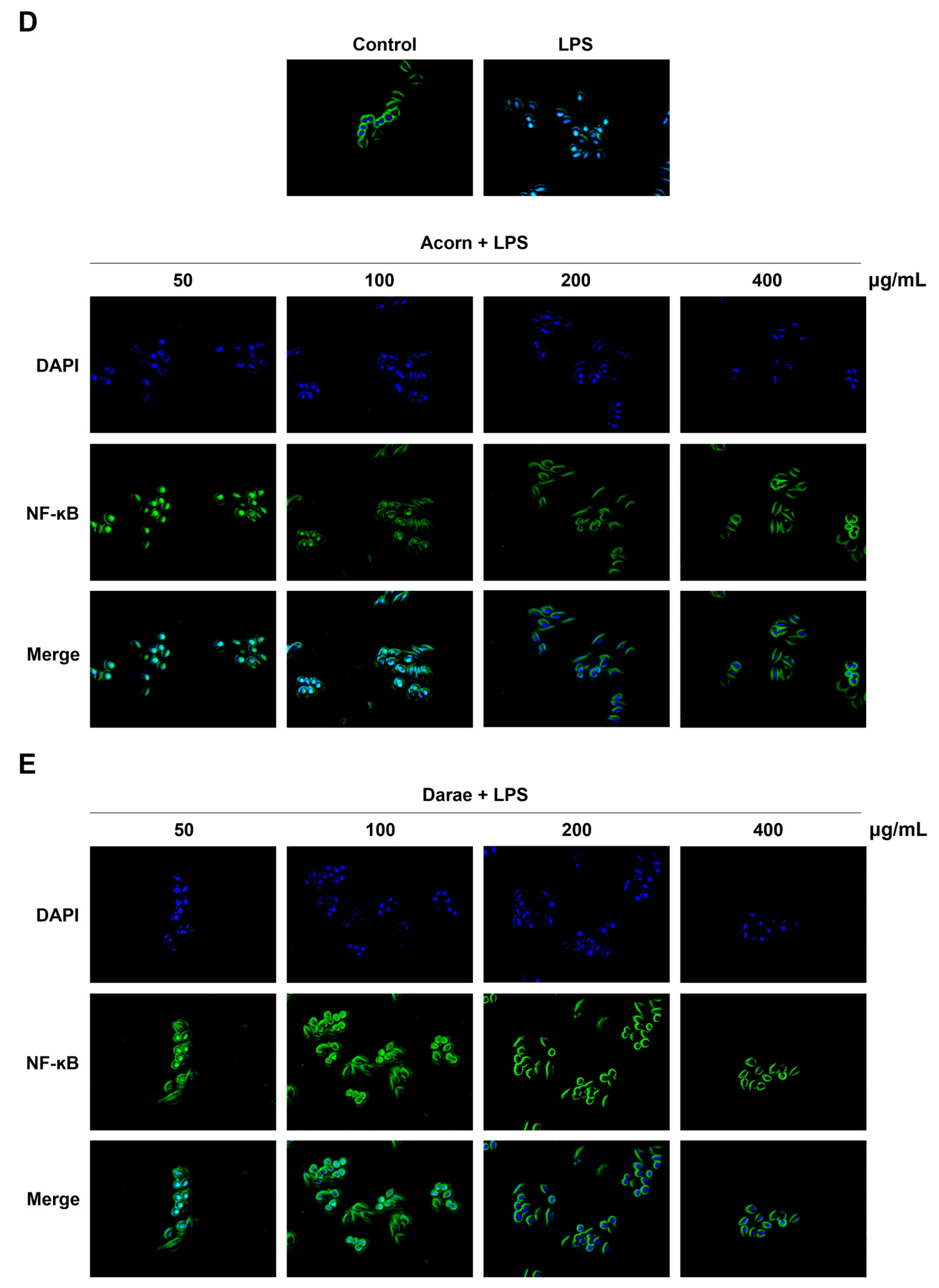
3.6. Effect of EEBP from Acorn and Darae on NF-κB in RAW 264.7 Macrophages
3.7. Effect of EEBP from Acorn and Darae on MAPKs in RAW 264.7 Macrophages
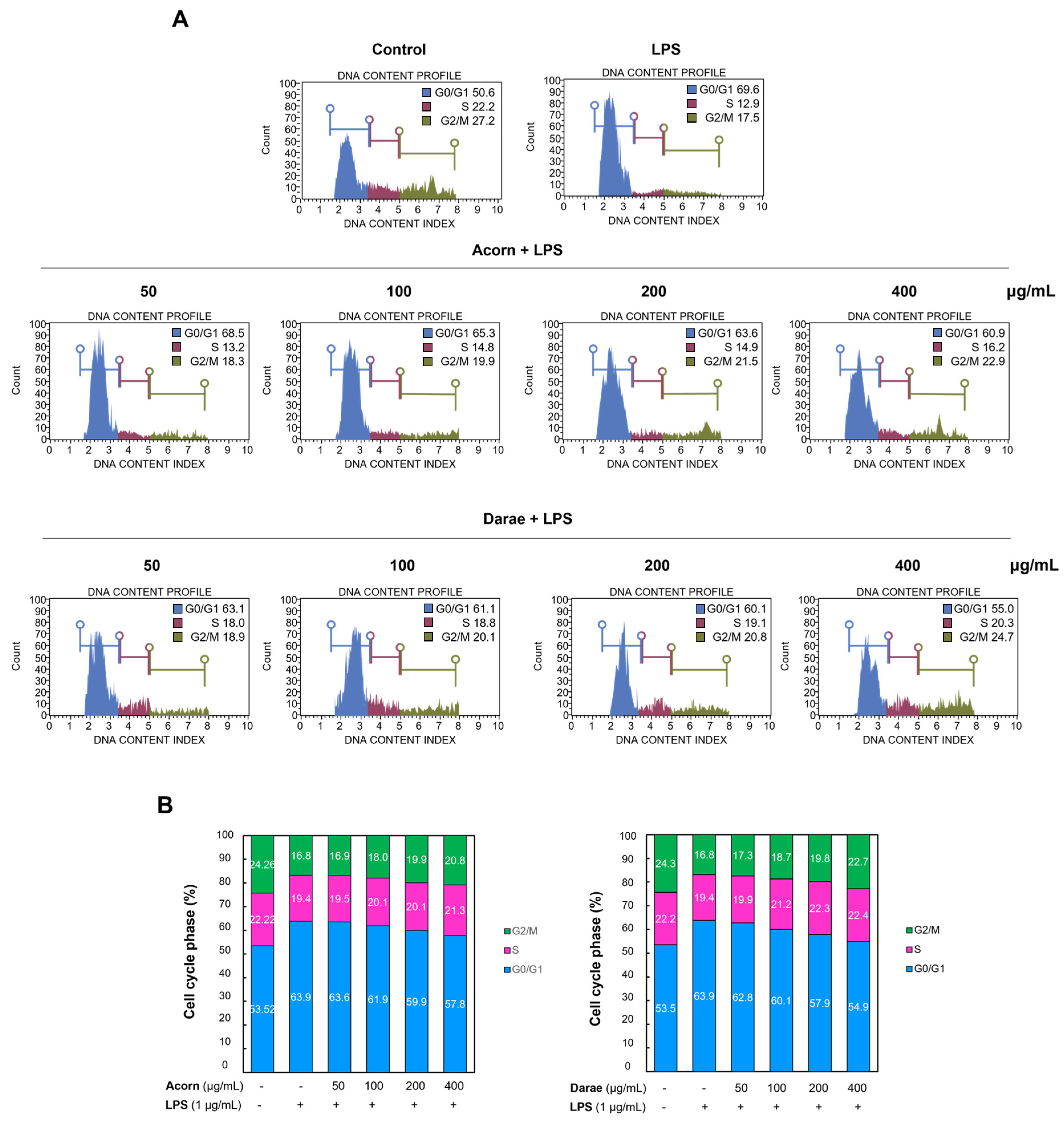
3.8. Effect of EEBP from Acorn and Darae on Cell Cycle in RAW 264.7 Macrophages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maldonado, E.; Morales-Pison, S.; Urina, F.; Solari, A. Aging hallmarks and the role of oxidative stress. Antioxidants 2023, 12, 651. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.Y.; Kim, D.H.; Lee, E.K.; Chung, K.W.; Chung, S.W.; Lee, B.G.; Seo, A.Y.; Chung, J.H.; Jung, Y.S.; Im, E.; et al. Redefining chronic inflammation in aging and age-related diseases: Proposal of the senoinflammation concept. Aging Dis. 2019, 10, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Martinello, M.; Mutinelli, F. Antioxidant activity in bee products: A review. Antioxidants 2021, 10, 71. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar]
- Lim, S.L.; Samaila, M.C.; Thuaibah, H.; Siti, K.A.; Mohamad, A.M.M.; Zauyah, Y.; Shamima, A.R. Antioxidant and anti-inflammatory properties of Erythroxylum cuneatum alkaloid leaf extract. Heliyon 2020, 6, e04141. [Google Scholar]
- Ali, A.M.; Kunugi, H. Apitherapy for Age-Related Skeletal Muscle Dysfunction (Sarcopenia): A Review on the Effects of Royal Jelly, Propolis, and Bee Pollen. Foods 2020, 9, 1362. [Google Scholar] [CrossRef] [PubMed]
- Algethami, J.S.; El-Wahed, A.A.A.; Elashal, M.H.; Ahmed, H.R.; Elshafiey, E.H.; Omar, E.M.; Naggar, Y.A.; Algethami, A.F.; Shou, Q.; Alsharif, S.M.; et al. Bee Pollen: Clinical Trials and Patent Applications. Nutrients 2022, 14, 2858. [Google Scholar] [CrossRef] [PubMed]
- Mauriello, G.; De Prisco, A.; Di Prisco, G.; La Storia, A.; Caprio, E. Microbial characterization of bee pollen from the vesuvius area collected by using three different traps. PLoS ONE 2017, 12, e0183208. [Google Scholar] [CrossRef]
- Li, Q.Q.; Wang, K.; Marcucci, M.C.; Sawaya, A.C.H.F.; Hu, L.; Xue, X.F.; Wu, L.M.; Hu, F.L. Nutrient-rich bee pollen: A treasure trove of active natural metabolites. J. Funct. Foods 2018, 49, 472–484. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elashal, M.H.; Yosri, N.; Du, M.; Musharraf, S.G.; Nahar, L.; Sarker, S.D.; Guo, Z.; Cao, W.; Zou, X.; et al. Bee pollen: Current status and therapeutic potential. Nutrients 2021, 13, 1876. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. Rep. 2021, 209, 112891. [Google Scholar]
- Priya, L.B.; Baskaran, R.; Huang, C.Y.; Padma, V.V. Neferine ameliorates cardiomyoblast apoptosis induced by doxorubicin: Possible role in modulating NADPH oxidase/ROS-mediated NFκB redox signaling cascade. Sci. Rep. 2017, 7, 12283. [Google Scholar] [CrossRef]
- Qi, S.; Feng, Z.; Li, Q.; Qi, Z.; Zhang, Y. Myricitrin modulates NADPH oxidase-dependent ROS production to inhibit endotoxin-mediated inflammation by blocking the JAK/STAT1 and NOX2/p47phox pathways. Oxidative Med. Cell. Longev. 2017, 2017, 9738745. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Gao, H.; Hou, Y.; Yu, J.; Sun, W.; Wang, Y.; Chen, X.; Feng, Y.; Xu, Q.M.; Chen, X. Dihydronortanshinone, a natural product, alleviates LPS-induced inflammatory response through NF-κB, mitochondrial ROS, and MAPK pathways. Toxicol. Appl. Pharmacol. 2018, 355, 1–8. [Google Scholar] [CrossRef]
- vGuha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.J.; DiPietro, L.A. Inflammation and wound healing: The role of the macrophage. Expert. Rev. Mol. Med. 2011, 13, E23. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Eteraf-Oskouei, T.; Shafiee-Khamneh, A.; Heshmati-Afshar, F.; Delazar, A. Anti-inflammatory and anti-angiogenesis effect of bee pollen methanolic extract using air pouch model of inflammation. Res. Pharm. Sci. 2020, 15, 66–75. [Google Scholar] [CrossRef]
- Kosedag, M.; Gulaboglu, M. Pollen and bee bread expressed highest anti-inflammatory activities among bee products in chronic inflammation: An experimental study with cotton pellet granuloma in rats. Inflammopharmacology 2023, 31, 1967–1975. [Google Scholar] [CrossRef]
- Lopes, A.J.O.; Vasconcelos, C.C.; Pereira, F.A.N.; Silva, R.H.M.; Queiroz, P.F.D.S.; Fernandes, C.V.; Garcia, J.B.S.; Ramos, R.M.; da Rocha, C.Q.; Lima, S.T.J.R.M.; et al. Anti-inflammatory and antinociceptive activity of pollen extract collected by stingless bee melipona fasciculata. Int. J. Mol. Sci. 2019, 20, 4512. [Google Scholar] [CrossRef]
- Capparelli, S.; Pieracci, Y.; Coppola, F.; Marchioni, I.; Sagona, S.; Felicioli, A.; Pistelli, L.; Pistelli, P. The colors of Tuscan bee pollen: Phytochemical profile and antioxidant activity. Nat. Prod. Res. 2024, 38, 2313–2319. [Google Scholar] [CrossRef] [PubMed]
- Miyata, R.; Hoshino, S.; Ahn, M.-R.; Kumazawa, S. Chemical Profiles of Korean Bee Pollens and Their Catechol- O-methyltransferase Inhibitory Activities. J. Agric. Food Chem. 2022, 70, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.R.; Chung, M.J.; Park, J.W.; Cho, S.M.; Choi, D.J.; Kim, S.M.; Chun, M.H.; Kim, I.B.; Kim, S.O.; Jang, S.J.; et al. Antiasthma effects through anti-inflammatory action of acorn (Quercus acutissima carr.) in vitro and in vivo. J. Food Biochem. 2012, 37, 108–118. [Google Scholar] [CrossRef]
- Lee, J.W.; Jo, S.E.; Han, S.I.; Kim, J.H. Ethanol-extracted acorn induces anti-inflammatory effects in human keratinocyte and production of hyaluronic acid in human fibroblasts. J. Med. Food 2023, 26, 595–604. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Hiraiwa, Y.; Hagiya, Y.; Nakajima, M.; Tanaka, T.; Ogura, S.I. 5-Aminolevulinic acid regulates the immune response in LPS-stimulated RAW 264.7 macrophages. BMC Immunol. 2018, 19, 41. [Google Scholar] [CrossRef] [PubMed]
- Merly, L.; Smith, S.L. Murine RAW 264.7 cell line as an immune target: Are we missing something? Immunopharmacol. Immunotoxicol. 2017, 39, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.P.; Abraham, A. Inhibition of LPS induced pro-inflammatory responses in RAW 264.7 macrophage cells by PVP-coated naringenin nanoparticle via down regulation of NF-κB/P38MAPK mediated stress signaling. Pharmacol. Rep. 2017, 69, 908–915. [Google Scholar] [CrossRef]
- Park, J.; Kwak, C.H.; Ha, S.H.; Kwon, K.M.; Abekura, F.; Cho, S.H.; Chang, Y.C.; Lee, Y.C.; Ha, K.T.; Chung, T.W.; et al. Ganglioside GM3 suppresses lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophage cells through NF-κB, AP-1, and MAPKs signaling. J. Cell. Biochem. 2018, 119, 1173–1182. [Google Scholar] [CrossRef]
- Sun, H.; Cai, W.; Wang, X.; Liu, Y.; Hou, B.; Zhu, X.; Qiu, L. Vaccaria hypaphorine alleviates lipopolysaccharide-induced inflammation via inactivation of NFκB and ERK pathways in Raw 264.7 cells. BMC Complement. Altern. Med. 2017, 17, 120. [Google Scholar] [CrossRef]
- Robbins, K.S. Effect of pecan phenolics on the release of nitric oxide from murine RAW 264.7 macrophage cells. Food Chem. 2016, 212, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, H.; Jantan, I.; Haque, M.A.; Kumolosasi, E. Anti-inflammatory effects of Phyllanthus amarus Schum. & Thonn. through inhibition of NF-κB, MAPK, and PI3K-Akt signaling pathways in LPS-induced human macrophages. BMC Complement. Altern. Med. 2018, 18, 224. [Google Scholar]
- Prescott, J.A.; Mitchell, J.P.; Cook, S.J. Inhibitory feedback control of NF-κB signalling in health and disease. Biochem. J. 2021, 478, 2619–2664. [Google Scholar] [PubMed]
- Lee, M.; Lee, N.Y.; Chung, K.S.; Cheon, S.Y.; Lee, K.T.; An, H.J. Roxatidine attenuates mast cell-mediated allergic inflammation via inhibition of NF-κB and p38 MAPK activation. Sci. Rep. 2017, 7, 41721. [Google Scholar] [CrossRef]
- Guo, W.; Sun, J.; Jiang, L.; Duan, L.; Huo, M.; Chen, N.; Zhong, W.; Wassy, L.; Yang, Z.; Feng, H. Imperatorin attenuates LPS-induced inflammation by suppressing NF-κB and MAPKs activation in RAW 264.7 macrophages. Inflammation 2012, 35, 1764–1772. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef]
- Pautz, A.; Art, J.; Hahn, S.; Nowag, S.; Voss, C.; Kleinert, H. Regulation of the expression of inducible nitric oxide synthase. Nitric Oxide 2010, 23, 75–93. [Google Scholar] [PubMed]
- Xie, Q.W.; Kashiwabara, Y.; Nathan, C. Role of transcription factor NF-kappa B/Rel in induction of nitric oxide synthase. J. Biol. Chem. 1994, 269, 4705–4708. [Google Scholar] [CrossRef]
- Kaur, B.; Singh, P. Inflammation: Biochemistry, cellular targets, anti-inflammatory agents and challenges with special emphasis on cyclooxygenase-2. Bioorg Chem. 2022, 121, 105663. [Google Scholar] [CrossRef]
- Chou, A.H.; Lee, H.C.; Liao, C.C.; Yu, H.P.; Liu, F.C. ERK/NF-kB/COX-2 Signaling Pathway Plays a Key Role in Curcumin Protection against Acetaminophen-Induced Liver Injury. Life 2023, 13, 2150. [Google Scholar] [CrossRef]
- Heo, Y.J.; Lee, N.; Choi, S.E.; Jeon, J.Y.; Han, S.J.; Kim, D.J.; Kang, Y.; Lee, K.W.; Kim, H.J. Amphiregulin Induces iNOS and COX-2 Expression through NF-κB and MAPK Signaling in Hepatic Inflammation. Mediat. Inflamm. 2023, 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Lee, E.K.; Gong, S.Y.; Sohng, J.K.; Kang, Y.J. Sparassis crispa exerts anti-inflammatory activity via suppression of TLR-mediated NF-κB and MAPK signaling pathways in LPS-induced RAW 264.7 macrophage cells. J. Ethanopharmacol. 2019, 231, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Li, L.X.; Wang, Y.; Zhai, J.C.; Chen, G.T.; Hu, K. Gambogic acid induces heme oxygenase-1 through Nrf2 signaling pathway and inhibits NF-κB and MAPK activation to reduce inflammation in LPS-activated RAW 264.7 cells. Biomed. Pharmacother. 2019, 109, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Jang, E.; Kang, S.Y.; Shin, J.S.; Han, H.S.; Kim, T.W.; Lee, D.H.; Lee, J.H.; Jang, D.S.; Lee, K.T. Anti-inflammatory potential of patrineolignan B isolated from Patrinia scabra in LPS-stimulated macrophages via inhibition of NF-κB, AP-1, and JAK/STAT pathways. Int. Immunopharmacol. 2020, 86, 106726. [Google Scholar] [CrossRef]
- Ren, J.; Su, D.; Li, L.; Cai, H.; Zhang, M.; Zhai, J.; Li, M.; Wu, X.; Hu, K. Anti-inflammatory effects of Aureusidin in LPS-stimulated RAW264.7 macrophages via suppressing NF-κB and activating ROS- and MAPKs-dependent Nrf2/HO-1 signaling pathways. Toxicol. Appl. Pharmacol. 2020, 387, 114846. [Google Scholar]
- Alam, M.B.; Chowdhury, N.S.; Sohrab, M.H.; Rana, M.S.; Hasan, C.M.; Lee, S.H. Cerevisterol Alleviates Inflammation via Suppression of MAPK/NF-κB/AP-1 and Activation of the Nrf2/HO-1 Signaling Cascade. Biomolecules 2020, 10, 199. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, J.-E.; Lee, J.-Y.; Baek, J.-Y.; Kim, S.W.; Ahn, M.-R. The Antioxidant and Anti-Inflammatory Properties of Bee Pollen from Acorn (Quercus acutissima Carr.) and Darae (Actinidia arguta). Antioxidants 2024, 13, 981. https://doi.org/10.3390/antiox13080981
Kwak J-E, Lee J-Y, Baek J-Y, Kim SW, Ahn M-R. The Antioxidant and Anti-Inflammatory Properties of Bee Pollen from Acorn (Quercus acutissima Carr.) and Darae (Actinidia arguta). Antioxidants. 2024; 13(8):981. https://doi.org/10.3390/antiox13080981
Chicago/Turabian StyleKwak, Jeong-Eun, Joo-Yeon Lee, Ji-Yoon Baek, Sun Wook Kim, and Mok-Ryeon Ahn. 2024. "The Antioxidant and Anti-Inflammatory Properties of Bee Pollen from Acorn (Quercus acutissima Carr.) and Darae (Actinidia arguta)" Antioxidants 13, no. 8: 981. https://doi.org/10.3390/antiox13080981
APA StyleKwak, J.-E., Lee, J.-Y., Baek, J.-Y., Kim, S. W., & Ahn, M.-R. (2024). The Antioxidant and Anti-Inflammatory Properties of Bee Pollen from Acorn (Quercus acutissima Carr.) and Darae (Actinidia arguta). Antioxidants, 13(8), 981. https://doi.org/10.3390/antiox13080981