
In Vivo Antioxidant Activity of Common Dietary Flavonoids: Insights from the Yeast Model Saccharomyces cerevisiae




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Yeast Strains and Growth Conditions
2.3. Optimization of Bioactives Concentration
2.4. Assessment of Antioxidant Activity of Bioactive Chemicals
2.5. Measurement of Oxidative Damage
2.6. Determination of Yeast Antioxidant Capacity
2.7. Determination of Oxidized Glutathione Content
3. Results
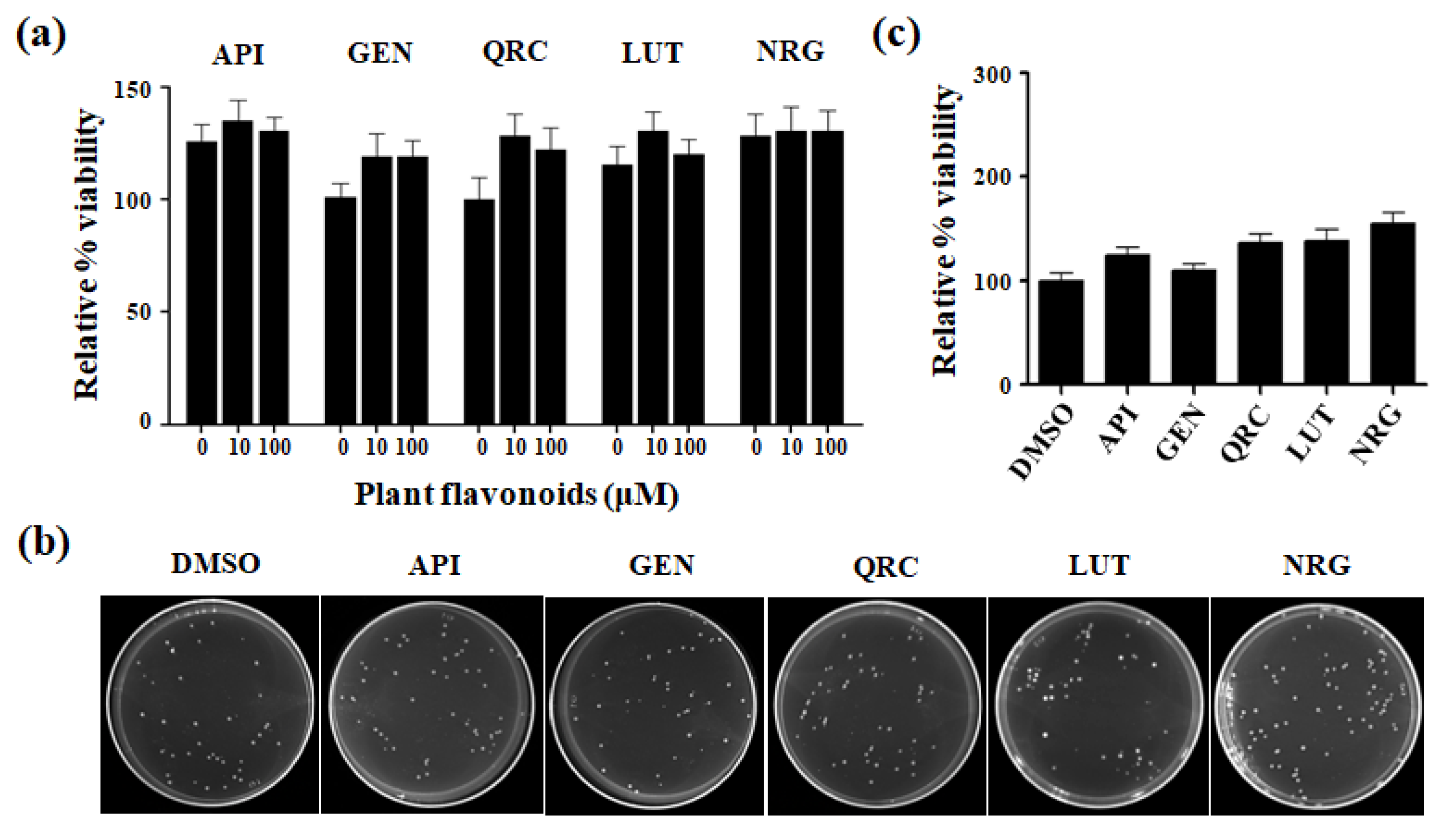
3.1. Effects of Selected Flavonoids on Cell Viability
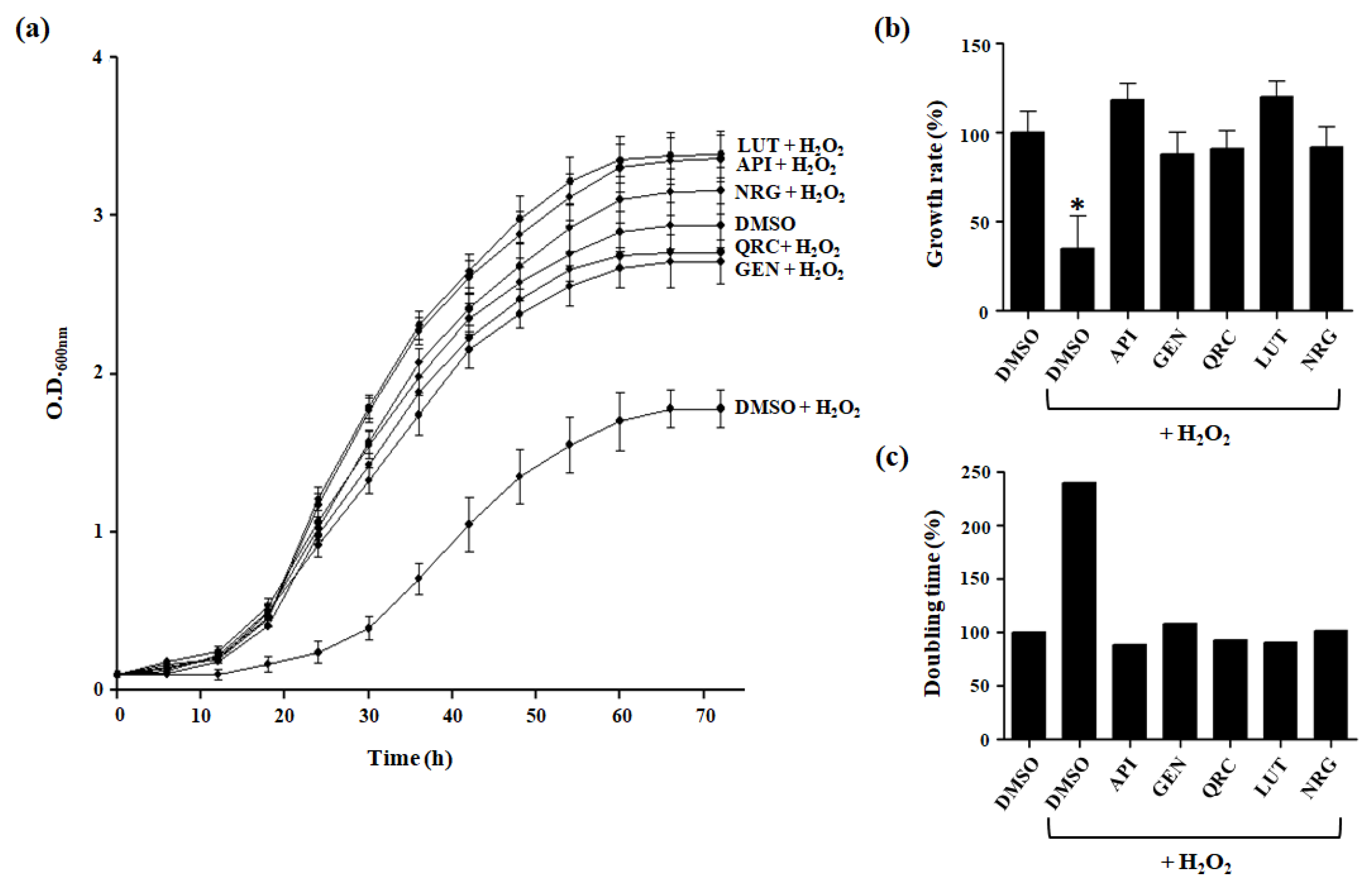
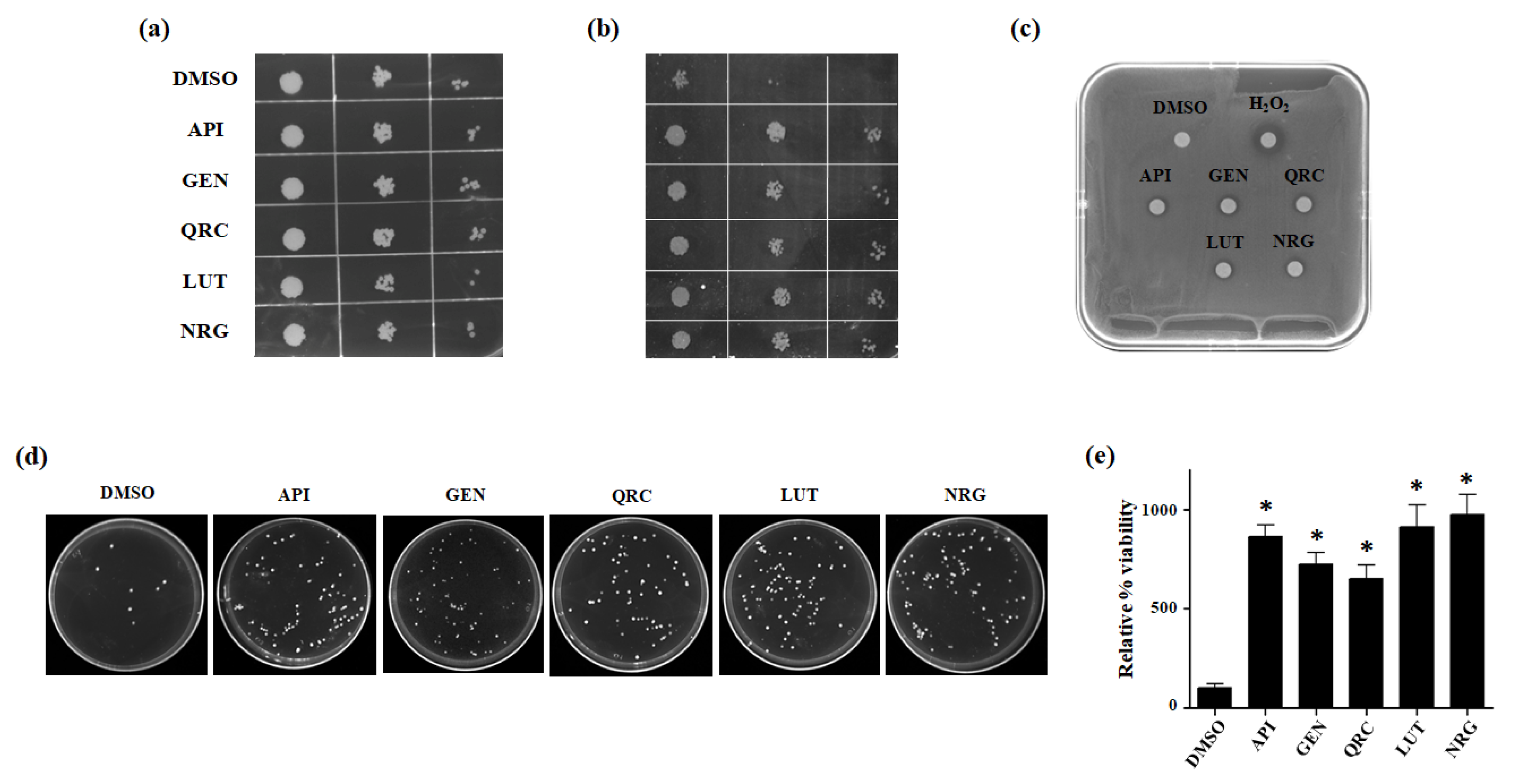
3.2. Effect of Selected Flavonoids on Oxidative Stress Tolerance
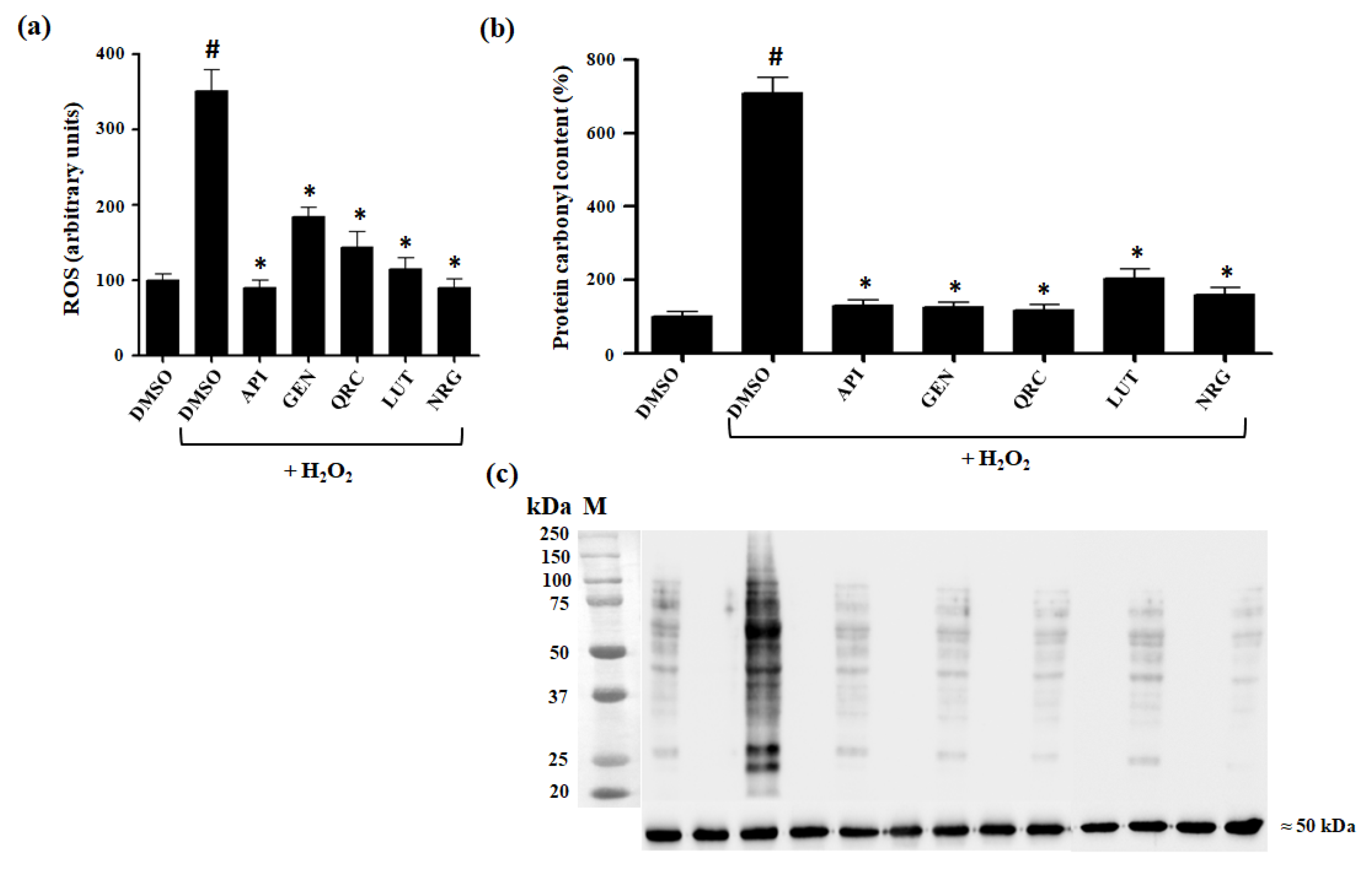
3.3. Effect of Selected Flavonoids on Intracelluar Oxidation
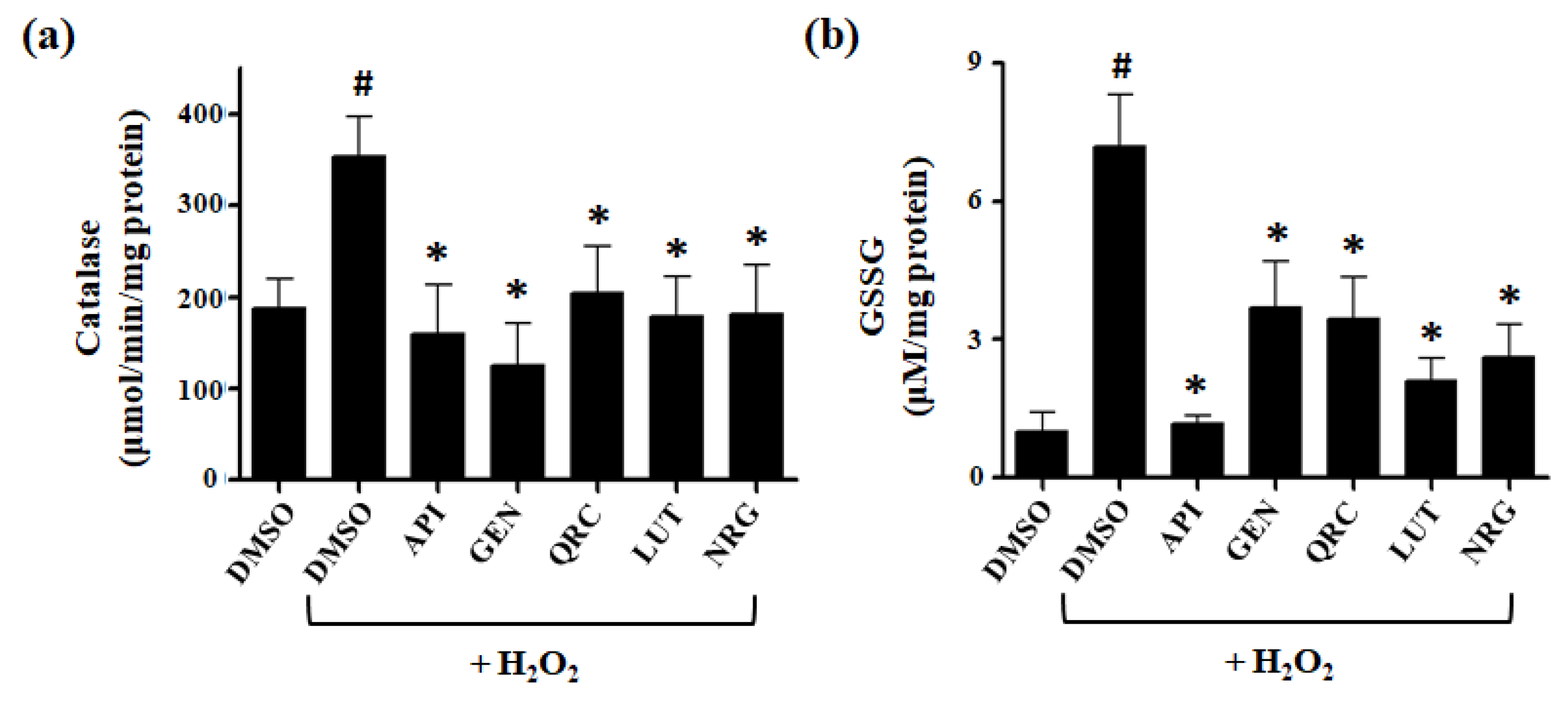
3.4. Effect of Selected Flavonoids on the Activity of Antioxidant Defense Systems
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, D.; Ding, Z.; Du, K.; Ye, X.; Cheng, S. Reactive Oxygen Species as a Link between Antioxidant Pathways and Autophagy. Oxid. Med. Cell Longev. 2021, 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-R.; Zweier, J.L. Cardiac Mitochondria and Reactive Oxygen Species Generation. Circ. Res. 2014, 114, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by Mitochondrial Reactive Oxygen Species in Physiology and Pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining Roles of Specific Reactive Oxygen Species (ROS) in Cell Biology and Physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Ferramosca, A.; Pinto Provenzano, S.; Montagna, D.D.; Coppola, L.; Zara, V. Oxidative Stress Negatively Affects Human Sperm Mitochondrial Respiration. Urology 2013, 82, 78–83. [Google Scholar] [CrossRef]
- Darenskaya, M.A.; Kolesnikova, L.I.; Kolesnikov, S.I. Oxidative Stress: Pathogenetic Role in Diabetes Mellitus and Its Complications and Therapeutic Approaches to Correction. Bull. Exp. Biol. Med. 2021, 171, 179–189. [Google Scholar] [CrossRef]
- Jelic, M.; Mandic, A.; Maricic, S.; Srdjenovic, B. Oxidative Stress and Its Role in Cancer. J. Cancer Res. Ther. 2021, 17, 22. [Google Scholar] [CrossRef] [PubMed]
- Shaito, A.; Aramouni, K.; Assaf, R.; Parenti, A.; Orekhov, A.; El Yazbi, A.; Pintus, G.; Eid, A.H. Oxidative Stress-Induced Endothelial Dysfunction in Cardiovascular Diseases. Front. Biosci. Landmark 2022, 27, 0105. [Google Scholar] [CrossRef]
- Briyal, S.; Ranjan, A.K.; Gulati, A. Oxidative Stress: A Target to Treat Alzheimer’s Disease and Stroke. Neurochem. Int. 2023, 165, 105509. [Google Scholar] [CrossRef]
- Dai, X.; Hu, Y.; Jiang, L.; Lei, L.; Fu, C.; Wu, S.; Zhang, X.; Zhu, L.; Zhang, F.; Chen, J.; et al. Decreased Oxidative Stress Response and Oxidant Detoxification of Skin during Aging. Mech. Ageing Dev. 2023, 216, 111878. [Google Scholar] [CrossRef] [PubMed]
- Zara, V.; Assalve, G.; Ferramosca, A. Insights into the Malfunctioning of the Mitochondrial Citrate Carrier: Implications for Cell Pathology. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2023, 1869, 166758. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, S.; Lin, C. Effect of Resveratrol and Pterostilbene on Aging and Longevity. BioFactors 2018, 44, 69–82. [Google Scholar] [CrossRef]
- Ferramosca, A.; Di Giacomo, M.; Zara, V. Antioxidant Dietary Approach in Treatment of Fatty Liver: New Insights and Updates. World J. Gastroenterol. 2017, 23, 4146. [Google Scholar] [CrossRef]
- Nani, A.; Murtaza, B.; Sayed Khan, A.; Khan, N.A.; Hichami, A. Antioxidant and Anti-Inflammatory Potential of Polyphenols Contained in Mediterranean Diet in Obesity: Molecular Mechanisms. Molecules 2021, 26, 985. [Google Scholar] [CrossRef]
- Omidifar, N.; Moghadami, M.; Mousavi, S.M.; Hashemi, S.A.; Gholami, A.; Shokripour, M.; Sohrabi, Z. Trends in Natural Nutrients for Oxidative Stress and Cell Senescence. Oxid. Med. Cell Longev. 2021, 2021, 1–7. [Google Scholar] [CrossRef]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and Human Health—A Focus on Oxidative Stress, Inflammation and Disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Di Giacomo, M.; Zara, V.; Bergamo, P.; Ferramosca, A. Crosstalk between Mitochondrial Metabolism and Oxidoreductive Homeostasis: A New Perspective for Understanding the Effects of Bioactive Dietary Compounds. Nutr. Res. Rev. 2020, 33, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Zara, V. Diet and Male Fertility: The Impact of Nutrients and Antioxidants on Sperm Energetic Metabolism. Int. J. Mol. Sci. 2022, 23, 2542. [Google Scholar] [CrossRef]
- Rajendran, M.; Manisankar, P.; Gandhidasan, R.; Murugesan, R. Free Radicals Scavenging Efficiency of a Few Naturally Occurring Flavonoids: A Comparative Study. J. Agric. Food Chem. 2004, 52, 7389–7394. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Lorenzetti, S.; Di Giacomo, M.; Lunetti, P.; Murrieri, F.; Capobianco, L.; Dolce, V.; Coppola, L.; Zara, V. Modulation of Human Sperm Mitochondrial Respiration Efficiency by Plant Polyphenols. Antioxidants 2021, 10, 217. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-I.; Hu, W.-S.; Hung, M.-Y.; Ou, H.-C.; Huang, S.-H.; Hsu, P.-T.; Day, C.-H.; Lin, K.-H.; Viswanadha, V.P.; Kuo, W.-W.; et al. Protective Effects of Luteolin against Oxidative Stress and Mitochondrial Dysfunction in Endothelial Cells. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 1032–1043. [Google Scholar] [CrossRef]
- Kashyap, P.; Shikha, D.; Thakur, M.; Aneja, A. Functionality of Apigenin as a Potent Antioxidant with Emphasis on Bioavailability, Metabolism, Action Mechanism and in Vitro and in Vivo Studies: A Review. J. Food Biochem. 2022, 46, e13950. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.-J.; Chen, R.-J.; Chen, L.; Chen, S.; Yang, X.-F.; Min, J.-W. Genistein Mitigates Oxidative Stress and Inflammation by Regulating Nrf2/HO-1 and NF-ΚB Signaling Pathways in Hypoxic-Ischemic Brain Damage in Neonatal Mice. Ann. Transl. Med. 2022, 10, 32. [Google Scholar] [CrossRef]
- Saberi-Hasanabadi, P.; Sedaghatnejad, R.; Mohammadi, H. Protective Effect of Quercetin against Paraquat-Induced Brain Mitochondrial Disruption in Mice. Curr. Drug Saf. 2024, 19, 44–50. [Google Scholar] [CrossRef]
- Yuan, R.; Venitz, J. Effect of Chronic Renal Failure on the Disposition of Highly Hepatically Metabolized Drugs. Int. J. Clin. Pharmacol. Ther. 2000, 38, 245–253. [Google Scholar] [CrossRef]
- Katoch, B.; Begum, R. Biochemical Basis of the High Resistance to Oxidative Stress InDictyostelium Discoideum. J. Biosci. 2003, 28, 581–588. [Google Scholar] [CrossRef]
- González Siso, M.I.; Cerdán, M.E. Kluyveromyces Lactis: A Suitable Yeast Model to Study Cellular Defense Mechanisms against Hypoxia-Induced Oxidative Stress. Oxid. Med. Cell Longev. 2012, 2012, 1–14. [Google Scholar] [CrossRef]
- Bayliak, M.M.; Burdylyuk, N.I.; Lushchak, V.I. Quercetin Increases Stress Resistance in the Yeast Saccharomyces Cerevisiae Not Only as an Antioxidant. Ann. Microbiol. 2016, 66, 569–576. [Google Scholar] [CrossRef]
- Xing, J.; Liu, P.; Zhao, L.; Huang, F. Deletion of CGLD1 Impairs PSII and Increases Singlet Oxygen Tolerance of Green Alga Chlamydomonas Reinhardtii. Front. Plant Sci. 2017, 8, 2154. [Google Scholar] [CrossRef] [PubMed]
- Agus, H.H.; Sengoz, C.O.; Yilmaz, S. Oxidative Stress-Mediated Apoptotic Cell Death Induced by Camphor in Sod1-Deficient Schizosaccharomyces pombe. Toxicol. Res. 2019, 8, 216–226. [Google Scholar] [CrossRef]
- Gao, Y.; Fang, L.; Wang, X.; Lan, R.; Wang, M.; Du, G.; Guan, W.; Liu, J.; Brennan, M.; Guo, H.; et al. Antioxidant Activity Evaluation of Dietary Flavonoid Hyperoside Using Saccharomyces Cerevisiae as a Model. Molecules 2019, 24, 788. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, H.; Watchaputi, K.; Hata, J.; Pabuprapap, W.; Suksamrarn, A.; Chua, L.S.; Soontorngun, N. Model Yeast as a Versatile Tool to Examine the Antioxidant and Anti-Ageing Potential of Flavonoids, Extracted from Medicinal Plants. Front. Pharmacol. 2022, 13, 980066. [Google Scholar] [CrossRef]
- Gosselin-Monplaisir, T.; Dagkesamanskaya, A.; Rigal, M.; Floch, A.; Furger, C.; Martin-Yken, H. A New Role for Yeast Cells in Health and Nutrition: Antioxidant Power Assessment. Int. J. Mol. Sci. 2023, 24, 11800. [Google Scholar] [CrossRef]
- Gröger, A.; Martínez-Albo, I.; Albà, M.M.; Ayté, J.; Vega, M.; Hidalgo, E. Comparing Mitochondrial Activity, Oxidative Stress Tolerance, and Longevity of Thirteen Ascomycota Yeast Species. Antioxidants 2023, 12, 1810. [Google Scholar] [CrossRef]
- Assalve, G.; Lunetti, P.; Zara, V.; Ferramosca, A. Ctp1 and Yhm2: Two Mitochondrial Citrate Transporters to Support Metabolic Flexibility of Saccharomyces Cerevisiae. Int. J. Mol. Sci. 2024, 25, 1870. [Google Scholar] [CrossRef] [PubMed]
- Magistrati, M.; Gilea, A.I.; Gerra, M.C.; Baruffini, E.; Dallabona, C. Drug Drop Test: How to Quickly Identify Potential Therapeutic Compounds for Mitochondrial Diseases Using Yeast Saccharomyces Cerevisiae. Int. J. Mol. Sci. 2023, 24, 10696. [Google Scholar] [CrossRef]
- Sudharshan, S.; Veerabhadrappa, B.; Subramaniyan, S.; Dyavaiah, M. Astaxanthin Enhances the Longevity of Saccharomyces Cerevisiae by Decreasing Oxidative Stress and Apoptosis. FEMS Yeast Res. 2019, 19, foy113. [Google Scholar] [CrossRef]
- De Blasi, G.; Lunetti, P.; Zara, V.; Ferramosca, A. Mitochondrial Citrate Transporters Ctp1-Yhm2 and Respiratory Chain: A Coordinated Functional Connection in Saccharomyces Cerevisiae Metabolism. Int. J. Biol. Macromol. 2024, 270, 132364. [Google Scholar] [CrossRef] [PubMed]
- Martins, D.; English, A.M. Catalase Activity Is Stimulated by H2O2 in Rich Culture Medium and Is Required for H2O2 Resistance and Adaptation in Yeast. Redox Biol. 2014, 2, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Jamnik, P.; Medved, P.; Raspor, P. Increased Glutathione Content in YeastSaccharomyces Cerevisiae Exposed to NaCl. Ann. Microbiol. 2006, 56, 175–178. [Google Scholar] [CrossRef]
- Owen, J.B.; Butterfield, D.A. Measurement of Oxidized/Reduced Glutathione Ratio. Methods Mol. Biol. 2010, 648, 269–277. [Google Scholar] [CrossRef]
- Zhao, Z. Hydroxyl Radical Generations Form the Physiologically Relevant Fenton-like Reactions. Free Radic. Biol. Med. 2023, 208, 510–515. [Google Scholar] [CrossRef]
- Rota, C.; Fann, Y.C.; Mason, R.P. Phenoxyl Free Radical Formation during the Oxidation of the Fluorescent Dye 2′,7′-Dichlorofluorescein by Horseradish Peroxidase. J. Biol. Chem. 1999, 274, 28161–28168. [Google Scholar] [CrossRef]
- Fernandes, P.N.; Mannarino, S.C.; Silva, C.G.; Pereira, M.D.; Panek, A.D.; Eleutherio, E.C.A. Oxidative Stress Response in Eukaryotes: Effect of Glutathione, Superoxide Dismutase and Catalase on Adaptation to Peroxide and Menadione Stresses in Saccharomyces cerevisiae. Redox Rep. 2007, 12, 236–244. [Google Scholar] [CrossRef]
- Jamieson, D.J. Oxidative Stress Responses of the Yeast Saccharomyces Cerevisiae. Yeast 1998, 14, 1511–1527. [Google Scholar] [CrossRef]
- Battino, M.; Forbes-Hernández, T.Y.; Gasparrini, M.; Afrin, S.; Cianciosi, D.; Zhang, J.; Manna, P.P.; Reboredo-Rodríguez, P.; Varela Lopez, A.; Quiles, J.L.; et al. Relevance of Functional Foods in the Mediterranean Diet: The Role of Olive Oil, Berries and Honey in the Prevention of Cancer and Cardiovascular Diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 893–920. [Google Scholar] [CrossRef]
- Koleva, D.I.; Petrova, V.Y.; Kujumdzieva, A.V. Comparison of Enzymatic Antioxidant Defence Systems in Different Metabolic Types of Yeasts. Can. J. Microbiol. 2008, 54, 957–963. [Google Scholar] [CrossRef]
- Costa, V.; Moradas-Ferreira, P. Oxidative Stress and Signal Transduction in Saccharomyces Cerevisiae: Insights into Ageing, Apoptosis and Diseases. Mol. Aspects Med. 2001, 22, 217–246. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of Its Protective Roles, Measurement, and Biosynthesis. Mol. Aspects Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Izawa, S.; Inoue, Y.; Kimura, A. Oxidative Stress Response in Yeast: Effect of Glutathione on Adaptation to Hydrogen Peroxide Stress in Saccharomyces cerevisiae. FEBS Lett. 1995, 368, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Penninckx, M. An Overview on Glutathione in versus Non-Conventional Yeasts. FEMS Yeast Res. 2002, 2, 295–305. [Google Scholar] [CrossRef]
- Inoue, Y.; Matsuda, T.; Sugiyama, K.; Izawa, S.; Kimura, A. Genetic Analysis of Glutathione Peroxidase in Oxidative Stress Response of Saccharomyces Cerevisiae. J. Biol. Chem. 1999, 274, 27002–27009. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Rate (h−1) | Doubling Time (h) | ||
|---|---|---|---|
| −H2O2 | DMSO | 0.120 ± 0.012 | 5.78 |
| +H2O2 | DMSO | 0.047 ± 0.015 * | 14.61 |
| API | 0.143 ± 0.012 | 4.87 | |
| GEN | 0.109 ± 0.009 | 6.37 | |
| QRC | 0.118 ± 0.011 | 5.88 | |
| LUT | 0.145 ± 0.011 | 4.79 | |
| NRG | 0.119 ± 0.013 | 5.82 | |
| API | 0.143 ± 0.012 | 4.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assalve, G.; Lunetti, P.; Zara, V.; Ferramosca, A. In Vivo Antioxidant Activity of Common Dietary Flavonoids: Insights from the Yeast Model Saccharomyces cerevisiae. Antioxidants 2024, 13, 1103. https://doi.org/10.3390/antiox13091103
Assalve G, Lunetti P, Zara V, Ferramosca A. In Vivo Antioxidant Activity of Common Dietary Flavonoids: Insights from the Yeast Model Saccharomyces cerevisiae. Antioxidants. 2024; 13(9):1103. https://doi.org/10.3390/antiox13091103
Chicago/Turabian StyleAssalve, Graziana, Paola Lunetti, Vincenzo Zara, and Alessandra Ferramosca. 2024. "In Vivo Antioxidant Activity of Common Dietary Flavonoids: Insights from the Yeast Model Saccharomyces cerevisiae" Antioxidants 13, no. 9: 1103. https://doi.org/10.3390/antiox13091103