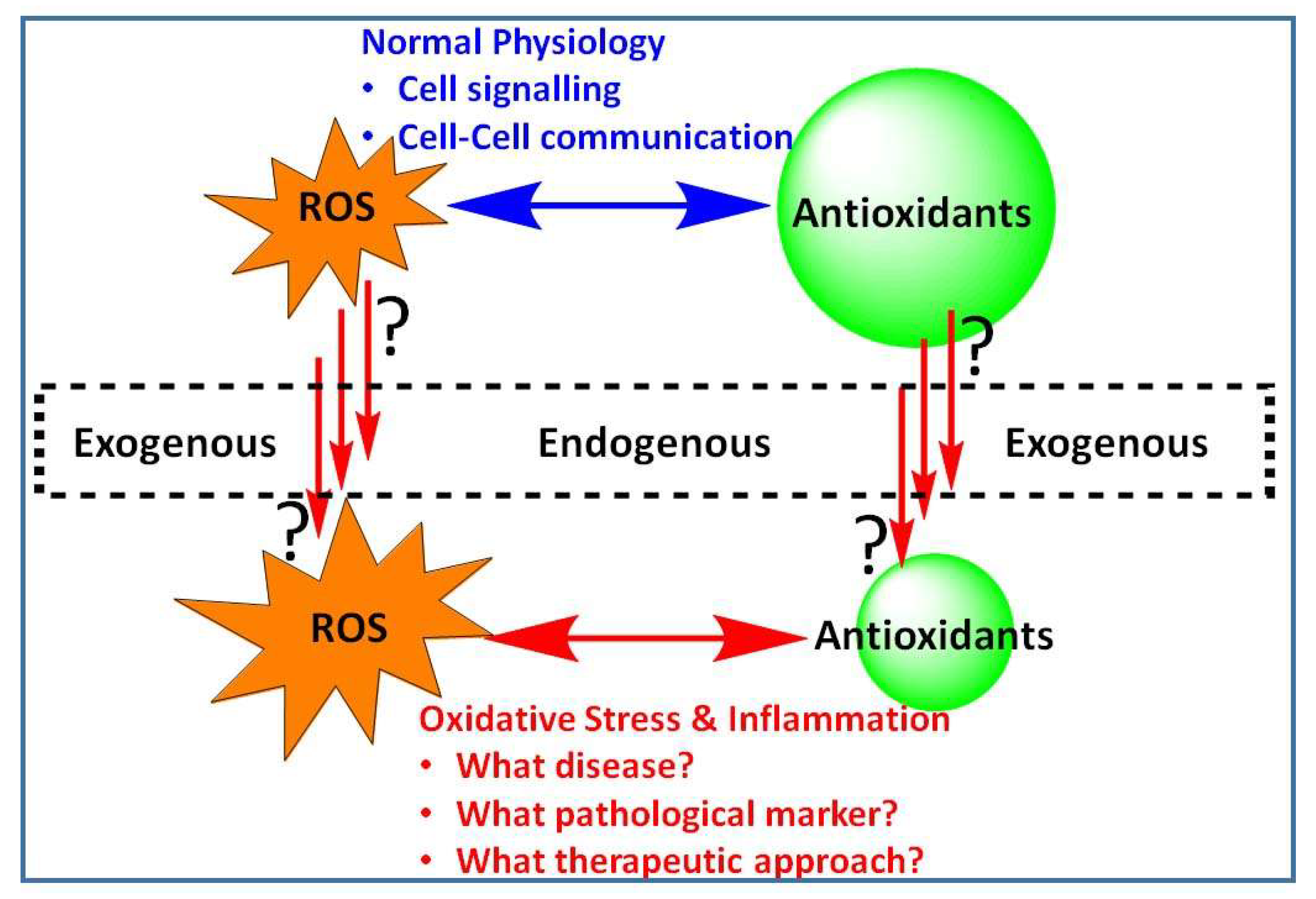
Modulation of Reactive Oxygen Species in Health and Disease

{kind=link}
Funding
Conflicts of Interest
References
- Rugemalira, E.; Roine, I.; Kuligowski, J.; Sánchez-Illana, Á.; Piñeiro-Ramos, J.D.; Andersson, S.; Peltola, H.; Leite Cruzeiro, M.; Pelkonen, T.; Vento, M. Protein oxidation biomarkers and myeloperoxidase activation in cerebrospinal fluid in childhood bacterial meningitis. Antioxidants 2019, 8, 441. [Google Scholar] [CrossRef] [PubMed]
- Kalita, J.; Misra, U.K.; Dubey, A.K. Role of oxidative stress in tuberculous meningitis: A clinico-radiological correlation. J. Mol. Neurosci. 2019, 68, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Matsuo, T.; Mitsunari, K.; Asai, A.; Ohba, K.; Sakai, H. A review of oxidative stress and urinary dysfunction caused by bladder outlet obstruction and treatments using antioxidants. Antioxidants 2019, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Choo, M.S.; Piao, S.; Oh, S.J. The preventive effect of a free radical scavenger on oxidative stress after the relief of partial bladder outlet obstruction in a rat model. PLoS ONE 2018, 13, e0199800. [Google Scholar] [CrossRef]
- Gu, M.; Liu, C.; Wan, X.; Yang, T.; Chen, Y.; Zhou, J.; Chen, Q.; Wang, Z. Epigallocatechin gallate attenuates bladder dysfunction via suppression of oxidative stress in a rat model of partial bladder outlet obstruction. Oxid. Med. Cell. Longev. 2018, 2018, 1393641. [Google Scholar] [CrossRef]
- Sezginer, E.K.; Yilmaz-Oral, D.; Lokman, U.; Nebioglu, S.; Aktan, F.; Gur, S. Effects of varying degrees of partial bladder outlet obstruction on urinary bladder function of rats: A novel link to inflammation, oxidative stress and hypoxia. Low. Urin. Tract. Symptoms. 2019, 11, O193–O201. [Google Scholar] [CrossRef]
- Todorović, A.; Pejić, S.; Gavrilović, L.; Pavlović, I.; Stojiljković, V.; Popović, N.; Pajović, S.B. Expression of antioxidant enzymes in patients with uterine polyp, myoma, hyperplasia, and adenocarcinoma. Antioxidants 2019, 8, 97. [Google Scholar] [CrossRef]
- Mallay, S.; Gill, R.; Young, A.; Mailloux, R.J. Sex-dependent differences in the bioenergetics of liver and muscle mitochondria from mice containing a deletion for glutaredoxin-2. Antioxidants 2019, 8, 245. [Google Scholar] [CrossRef]
- Kanaan, G.N.; Ichim, B.; Gharibeh, L.; Maharsy, W.; Patten, D.A.; Xuan, J.Y.; Reunov, A.; Marshall, P.; Veinot, J.; Menzies, K.; et al. Glutaredoxin-2 controls cardiac mitochondrial dynamics and energetics in mice, and protects against human cardiac pathologies. Redox Biol. 2018, 14, 509–521. [Google Scholar] [CrossRef]
- Wohua, Z.; Weiming, X. Glutaredoxin 2 (GRX2) deficiency exacerbates high fat diet (HFD)-induced insulin resistance, inflammation and mitochondrial dysfunction in brain injury: A mechanism involving GSK-3β. Biomed. Pharmacother. 2019, 118, 108940. [Google Scholar] [CrossRef]
- Wu, H.; Yu, Y.; David, L.; Ho, Y.S.; Lou, M.F. Glutaredoxin 2 (Grx2) gene deletion induces early onset of age-dependent cataracts in mice. J. Biol. Chem. 2014, 289, 36125–36139. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Lee, H.; Park, C.; Hong, S.H.; Hong, S.H.; Kim, G.Y.; Cha, H.J.; Kim, S.; Kim, H.S.; Hwang, H.J.; et al. Glutathione Induced Immune-Stimulatory Activity by Promoting M1-Like Macrophages Polarization via Potential ROS Scavenging Capacity. Antioxidants 2019, 8, 413. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Iridoids and Other Monoterpenes in the Alzheimer’s Brain: Recent Development and Future Prospects. Molecules 2018, 23, 117. [Google Scholar] [CrossRef]
- Habtemariam, S. Antioxidant and Anti-inflammatory Mechanisms of Neuroprotection by Ursolic Acid: Addressing Brain Injury, Cerebral Ischemia, Cognition Deficit, Anxiety, and Depression. Oxid. Med. Cell. Longev. 2019, 2019, 8512048. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Sureda, A.; Habtemariam, S.; Nabavi, S.M. Ginsenoside Rd and ischemic stroke; a short review of literatures. J. Ginseng Res. 2015, 39, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Espino, J.; Macedo, M.; Lozano, G.; Ortiz, Á.; Rodríguez, C.; Rodríguez, A.B.; Bejarano, I. Impact of Melatonin Supplementation in Women with Unexplained Infertility Undergoing Fertility Treatment. Antioxidants 2019, 8, 338. [Google Scholar] [CrossRef]
- Yang, Y.; Cheung, H.H.; Zhang, C.; Wu, J.; Chan, W.Y. Melatonin as potential targets for delaying ovarian aging. Curr. Drug Targets. 2019, 20, 16–28. [Google Scholar] [CrossRef]
- Kırıkçı, G.; Altunkaynak, B.Z.; Kaplan, S. Is melatonin, leptin or their combination more effective on oxidative stress and folliculogenesis in the obese rats? J. Obstet. Gynaecol. 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Zhang, X.; Xiong, Y.; Tan, Y.; Wang, L.; Li, R.; Zhang, Y.; Liu, X.; Lin, X.; Jin, L.; Hu, Y.; et al. Melatonin rescues impaired penetration ability of human spermatozoa induced by mitochondrial dysfunction. Reproduction 2019, REP-19-0231.R2. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, Q.; Wei, R.; Song, H.; Mi, J.; Lin, Z.; Yang, Y.; Sun, Z.; Zou, K. Melatonin protects spermatogonia from the stress of chemotherapy and oxidation via eliminating reactive oxidative species. Free Radic. Biol. Med. 2019, 137, 74–86. [Google Scholar] [CrossRef]
- Perumal, P.; Chang, S.; Baruah, K.K.; Srivastava, N. Administration of slow release exogenous melatonin modulates oxidative stress profiles and in vitro fertilizing ability of the cryopreserved mithun (Bos frontalis) spermatozoa. Theriogenology 2018, 120, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Moayeri, A.; Mokhtari, T.; Hedayatpour, A.; Abbaszadeh, H.A.; Mohammadpour, S.; Ramezanikhah, H.; Shokri, S. Impact of melatonin supplementation in the rat spermatogenesis subjected to forced swimming exercise. Andrologia 2018, 50, 50. [Google Scholar] [CrossRef]
- Habtemariam, S. Antioxidant and rutin content analysis of leaves of the common buckwheat (Fagopyrum esculentum Moench) grown in the United Kingdom: A case study. Antioxidants 2019, 8, 160. [Google Scholar] [CrossRef]
- Habtemariam, S.; Lentini, G. The therapeutic potential of rutin for diabetes: An update. Mini Rev. Med. Chem. 2015, 15, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Rutin as a natural therapy for Alzheimer’s disease: Insights into its mechanisms of action. Curr. Med. Chem. 2016, 23, 860–873. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habtemariam, S. Modulation of Reactive Oxygen Species in Health and Disease. Antioxidants 2019, 8, 513. https://doi.org/10.3390/antiox8110513
Habtemariam S. Modulation of Reactive Oxygen Species in Health and Disease. Antioxidants. 2019; 8(11):513. https://doi.org/10.3390/antiox8110513
Chicago/Turabian StyleHabtemariam, Solomon. 2019. "Modulation of Reactive Oxygen Species in Health and Disease" Antioxidants 8, no. 11: 513. https://doi.org/10.3390/antiox8110513
APA StyleHabtemariam, S. (2019). Modulation of Reactive Oxygen Species in Health and Disease. Antioxidants, 8(11), 513. https://doi.org/10.3390/antiox8110513