Functional Analysis of Macromolecular Polysaccharides: Whitening, Moisturizing, Anti-Oxidant, and Cell Proliferation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Extraction from Giant African Land Snail (Achatina Fulica) and Heimiella Retispora
2.3. Evaporation Rate of A. fulica Extracts and H. retispora Extracts
2.4. Determination of 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) Radical Scavenging Capacity
2.5. Metal Chelating Activity
2.6. Reducing Power
2.7. Cytotoxicity Examinations
2.8. B16-F10 Cellular Tyrosinase Activity
2.9. Detection of ROS by 2’,7’-dichlorodihydrofluorescein Diacetate (DCFDA) Stain
2.10. Statistical Analysis
3. Results and Discussion
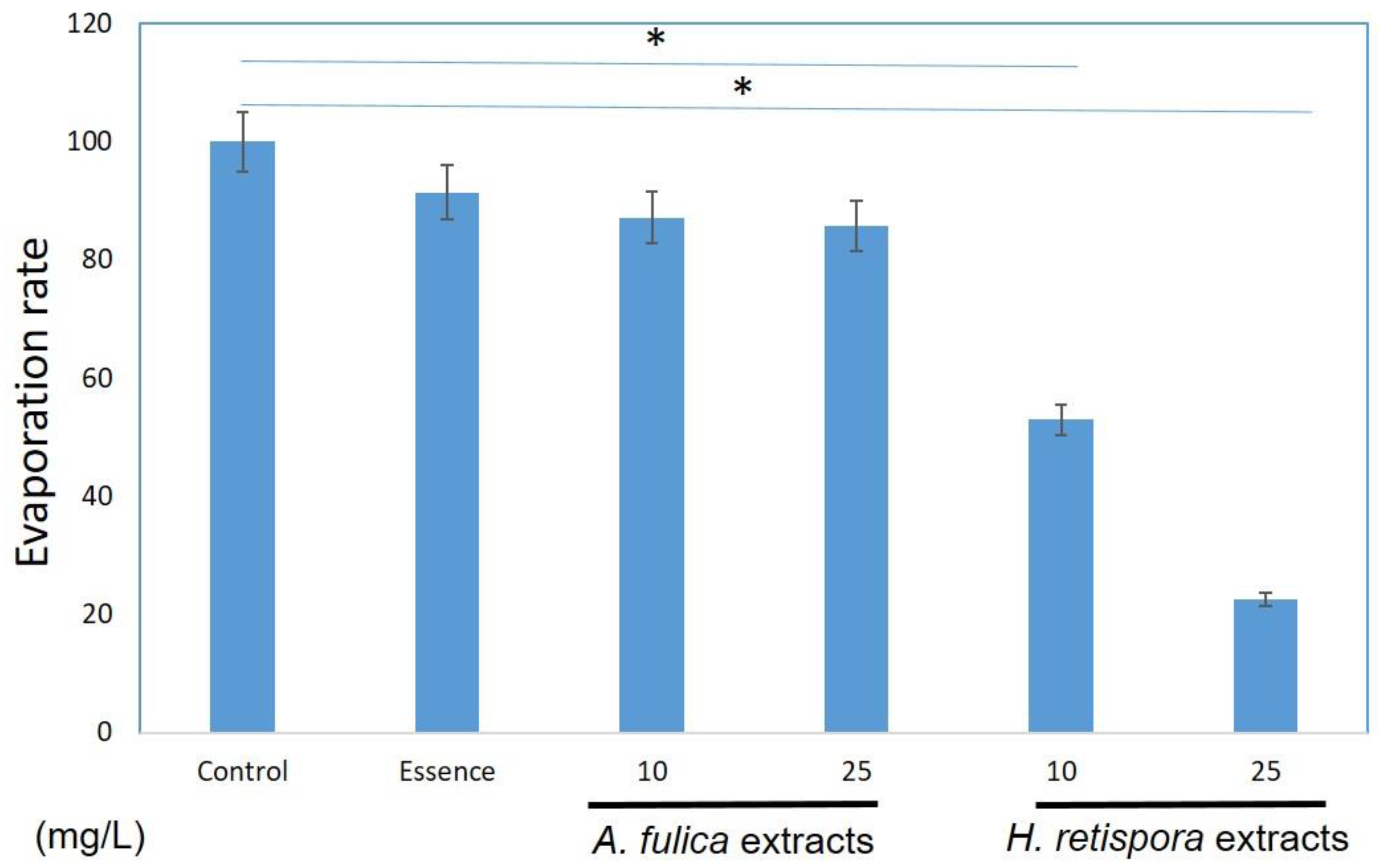
3.1. Moisturizing Activities
3.2. Anti-Oxidative Properties of the A. fulica Extracts and H. retispora Extracts
3.2.1. A. fulica Extracts and H. retispora Extracts Had No DPPH Free Radical Scavenging Activity
3.2.2. Ferrous Ions Chelating Capacity Measurements
3.2.3. Ferric Reducing Antioxidant Power (FRAP) Index Assessments
3.2.4. A. fulica Extracts and H. retispora Extracts Inhibit Intracellular ROS Accumulation
3.3. Cell Growth of A. fulica Extracts and H. retispora Extracts Treated in Human Fibroblasts
3.4. Collagen Productions in Sirius Red Assays
3.5. A. fulica Extracts and H. retispora Extracts on B16-F10 Cellular Tyrosinase Activity
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Data Availability Statement
References
- Li, P.H.; Liu, L.H.; Chang, C.C.; Gao, R.; Leung, C.H.; Ma, D.L.; David Wang, H.M. Silencing stem cell factor gene in fibroblasts to regulate paracrine factor productions and enhance c-Kit expression in melanocytes on melanogenesis. Int. J. Mol. Sci. 2018, 19, 1475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Murawsky, M.; LaCount, T.; Kasting, G.B.; Li, S.K. Transepidermal water loss and skin conductance as barrier integrity tests. Toxicol. Vitr. 2018, 51, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.; Brown, S.; Danby, S.; Flohr, C. Research techniques made simple: Transepidermal water loss measurement as a research tool. J. Investig. Dermatol. 2018, 138, 2295–2300.e1. [Google Scholar] [CrossRef] [PubMed]
- Grubauer, G.; Elias, P.M.; Feingold, K.R. Transepidermal water loss: The signal for recovery of barrier structure and function. J. Lipid Res. 1989, 30, 323–333. [Google Scholar] [PubMed]
- Wang, Y.-C.; Haung, X.-Y.; Chiu, C.-C.; Lin, M.-Y.; Lin, W.-H.; Chang, W.-T.; Tseng, C.-C.; Wang, H.-M.D. Inhibitions of melanogenesis via phyllanthus emblica fruit extract powder in B16F10 cells. Food Biosci. 2019, 28, 177–182. [Google Scholar] [CrossRef]
- Wu, P.Y.; Lin, T.Y.; Hou, C.W.; Chang, Q.X.; Wen, K.C.; Lin, C.Y.; Chiang, H.M. 1,2-Bis[(3-Methoxyphenyl)Methyl]Ethane-1,2-Dicarboxylic Acid Reduces UVB-Induced Photodamage In Vitro and In Vivo. Antioxidants 2019, 8, 452. [Google Scholar] [CrossRef]
- Seo, S.Y.; Sharma, V.K.; Sharma, N. Mushroom tyrosinase: Recent prospects. J. Agric. Food Chem. 2003, 51, 2837–2853. [Google Scholar] [CrossRef]
- Jimbow, K.; Obata, H.; Pathak, M.A.; Fitzpatrick, T.B. Mechanism of depigmentation by hydroquinone. J. Investig. Dermatol. 1974, 62, 436–449. [Google Scholar] [CrossRef]
- Omarsdottir, S.; Olafsdottir, E.S.; Freysdottir, J. Immunomodulating effects of lichen-derived polysaccharides on monocyte-derived dendritic cells. Int. Immunopharmacol. 2006, 6, 1642–1650. [Google Scholar] [CrossRef]
- Nishigori, C.; Yarosh, D.B.; Donawho, C.; Kripke, M.L. The immune system in ultraviolet carcinogenesis. J. Investig. Dermatol. Symp. Proc. 1996, 1, 143–146. [Google Scholar] [CrossRef]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Svobodova, A.R.; Galandakova, A.; Sianska, J.; Dolezal, D.; Ulrichova, J.; Vostalova, J. Acute exposure to solar simulated ultraviolet radiation affects oxidative stress-related biomarkers in skin, liver and blood of hairless mice. Biol. Pharm. Bull. 2011, 34, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Marostica, H.V.; Castro, L.S.; Goncalves, G.A.; Silva, F.M.S.; Bracht, L.; Bersani-Amado, C.A.; Peralta, R.M.; Comar, J.F.; Bracht, A.; Sa-Nakanishi, A.B. Methyl Jasmonate Reduces Inflammation and Oxidative Stress in the Brain of Arthritic Rats. Antioxidants 2019, 8, 485. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shang, F.; Yang, Z.; Wu, M.; Zhao, J. Structural analysis of a homogeneous polysaccharide from Achatina fulica. Int. J. Biol. Macromol. 2017, 98, 786–792. [Google Scholar] [CrossRef]
- Paszkiewicz, W.; Kozyra, I.; Rzeżutka, A. A refinement of an international standard method (ISO/TS 15216–2: 2013) to allow extraction and concentration of human enteric viruses from tissues of edible snail species. Food Anal. Method 2015, 8, 799–806. [Google Scholar] [CrossRef]
- Kim, Y.S.; Jo, Y.Y.; Chang, I.M.; Toida, T.; Park, Y.; Linhardt, R.J. A new glycosaminoglycan from the giant African snail Achatina fulica. J. Biol. Chem. 1996, 271, 11750–11755. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, Y.; Liu, H.X.; Hu, L.; Yang, K.; Steinmann, P.; Chen, Z.; Wang, L.Y.; Utzinger, J.; Zhou, X.N. Invasive snails and an emerging infectious disease: Results from the first national survey on Angiostrongylus cantonensis in China. PLoS Negl. Trop. Dis. 2009, 3, e368. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y. Structure and immunological activity of a novel polysaccharide from the spores of Ganoderma lucidum. Afr. J. Biotechnol. 2011, 10, 10923–10929. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Liu, Y.; Zhao, X.; Tang, Q.; Dernedde, J.; Zhang, J.; Fan, H. Anti-inflammatory properties of GLPss58, a sulfated polysaccharide from Ganoderma lucidum. Int. J. Biol. Macromol. 2018, 107, 486–493. [Google Scholar] [CrossRef]
- Xu, S.; Dou, Y.; Ye, B.; Wu, Q.; Wang, Y.; Hu, M.; Ma, F.; Rong, X.; Guo, J. Ganoderma lucidum polysaccharides improve insulin sensitivity by regulating inflammatory cytokines and gut microbiota composition in mice. J. Funct. Foods 2017, 38, 545–552. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, D.; Liu, Y.; Li, C.; Zhao, X.; Li, Y.; Li, W. Ganoderma lucidum polysaccharide inhibits prostate cancer cell migration via the protein arginine methyltransferase 6 signaling pathway. Mol. Med. Rep. 2018, 17, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.C.; Chen, C.Y.; Kuo, C.H.; Lin, Y.S.; Hwang, B.H.; Wang, T.K.; Kuo, Y.H.; Wang, H.D. 36H: A Novel Potent Inhibitor for Antimelanogenesis. Oxidative Med. Cell. Longev. 2018, 2018, 6354972. [Google Scholar] [CrossRef] [PubMed]
- Li, P.H.; Chiu, Y.P.; Shih, C.C.; Wen, Z.H.; Ibeto, L.K.; Huang, S.H.; Chiu, C.C.; Ma, D.L.; Leung, C.H.; Chang, Y.N.; et al. Biofunctional Activities of Equisetum ramosissimum Extract: Protective Effects against Oxidation, Melanoma, and Melanogenesis. Oxidative Med. Cell. Longev. 2016, 2016, 2853543. [Google Scholar] [CrossRef] [PubMed]
- Panchuk, R.R.; Skorokhyd, N.R.; Kozak, Y.S.; Lehka, L.V.; Moiseenok, A.G.; Stoika, R.S. Tissue-protective activity of selenomethionine and D-panthetine in B16 melanoma-bearing mice under doxorubicin treatment is not connected with their ROS scavenging potential. Croat. Med. J. 2017, 58, 171. [Google Scholar] [CrossRef] [PubMed]
- Lorz, L.R.; Yoo, B.C.; Kim, M.-Y.; Cho, J.Y. Anti-Wrinkling and Anti-Melanogenic Effect of Pradosia mutisii Methanol Extract. Int. J. Mol. Sci. 2019, 20, 1043. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, D.; Park, C.; Park, T.; Park, B. Anti-Inflammatory and Skin-Moisturizing Effects of a Flavonoid Glycoside Extracted from the Aquatic Plant Nymphoides indica in Human Keratinocytes. Molecules 2018, 23, 2342. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Tung, C.-H.; Chang, J.-H.; Hsieh, Y.-H.; Hsu, J.-C.; Ellis, A.V.; Liu, W.-C.; Yan, R.-H. Comparison of hydroxyl radical yields between photo-and electro-catalyzed water treatments. J. Taiwain Inst. Chem. Eng. 2014, 45, 1649–1654. [Google Scholar] [CrossRef]
- Ohshima, H.; Yoshie, Y.; Auriol, S.; Gilibert, I. Antioxidant and pro-oxidant actions of flavonoids: Effects on DNA damage induced by nitric oxide, peroxynitrite and nitroxyl anion. Free Radic. Biol. Med. 1998, 25, 1057–1065. [Google Scholar] [CrossRef]
- Chou, H.Y.; Lee, C.; Pan, J.L.; Wen, Z.H.; Huang, S.H.; Lan, C.W.; Liu, W.T.; Hour, T.C.; Hseu, Y.C.; Hwang, B.H.; et al. Enriched Astaxanthin Extract from Haematococcus pluvialis Augments Growth Factor Secretions to Increase Cell Proliferation and Induces MMP1 Degradation to Enhance Collagen Production in Human Dermal Fibroblasts. Int. J. Mol. Sci. 2016, 17, 955. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, B.; Webb, D.J. Cancer-associated fibroblasts modulate growth factor signaling and extracellular matrix remodeling to regulate tumor metastasis. Biochem. Soc. Trans. 2017, 45, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuzhalin, A.E.; Lim, S.Y.; Kutikhin, A.G.; Gordon-Weeks, A.N. Dynamic matrisome: ECM remodeling factors licensing cancer progression and metastasis. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Matsubayashi, K.; Miwa, N. Inhibitory effect of the water-soluble polymer-wrapped derivative of fullerene on UVA-induced melanogenesis via downregulation of tyrosinase expression in human melanocytes and skin tissues. Arch. Dermatol. Res. 2007, 299, 245–257. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Concentration (mg/L) | DPPH (%) | Chelating (%) | Reducing Power (OD700) |
|---|---|---|---|---|
| Vitamin C | 100 μM | 87.4 ± 0.1 | - | - |
| EDTA | 100 μM | - | 85.6 ± 0.1 | - |
| BHA | 100 μM | - | - | 0.67 ± 0.02 |
| A. fulica extracts | 10 | 21.83 ± 0.06 | N/A | 0.25 ± 0.06 |
| 25 | 40.15 ± 0.03 | N/A | 0.24 ± 0.02 | |
| H. retispora extracts | 10 | 14.48 ± 0.05 | N/A | 0.17 ± 0.01 |
| 25 | 28.03 ± 0.04 | N/A | 0.18 ± 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kao, C.-J.; Chou, H.-Y.; Lin, Y.-C.; Liu, Q.; David Wang, H.-M. Functional Analysis of Macromolecular Polysaccharides: Whitening, Moisturizing, Anti-Oxidant, and Cell Proliferation. Antioxidants 2019, 8, 533. https://doi.org/10.3390/antiox8110533
Kao C-J, Chou H-Y, Lin Y-C, Liu Q, David Wang H-M. Functional Analysis of Macromolecular Polysaccharides: Whitening, Moisturizing, Anti-Oxidant, and Cell Proliferation. Antioxidants. 2019; 8(11):533. https://doi.org/10.3390/antiox8110533
Chicago/Turabian StyleKao, Chien-Jen, Hsin-Yu Chou, Yu-Chen Lin, Qinghong Liu, and Hui-Min David Wang. 2019. "Functional Analysis of Macromolecular Polysaccharides: Whitening, Moisturizing, Anti-Oxidant, and Cell Proliferation" Antioxidants 8, no. 11: 533. https://doi.org/10.3390/antiox8110533
APA StyleKao, C.-J., Chou, H.-Y., Lin, Y.-C., Liu, Q., & David Wang, H.-M. (2019). Functional Analysis of Macromolecular Polysaccharides: Whitening, Moisturizing, Anti-Oxidant, and Cell Proliferation. Antioxidants, 8(11), 533. https://doi.org/10.3390/antiox8110533