Batryticatus Bombyx Protects Dopaminergic Neurons against MPTP-Induced Neurotoxicity by Inhibiting Oxidative Damage



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of BB Extract
2.2. Animals and Drug Administration
2.3. Rotarod and Pole Test
2.4. Brain Tissue Preparation
2.5. Immunohistochemistry (IHC) Analysis
2.6. Western Blot and Determination of Dopamine Content and SOD, CAT, and GSH Levels
2.7. Statistical Analyses
3. Results
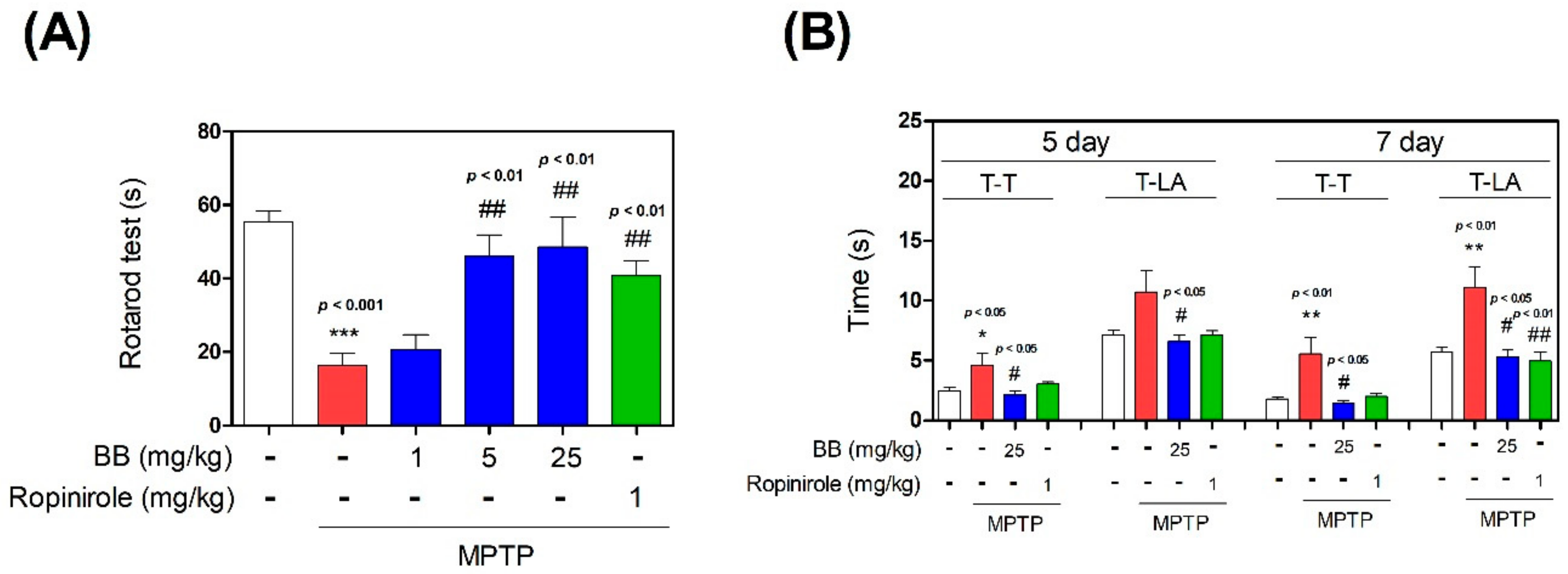
3.1. Effect of BB on MPTP-Induced Behavior Impairment
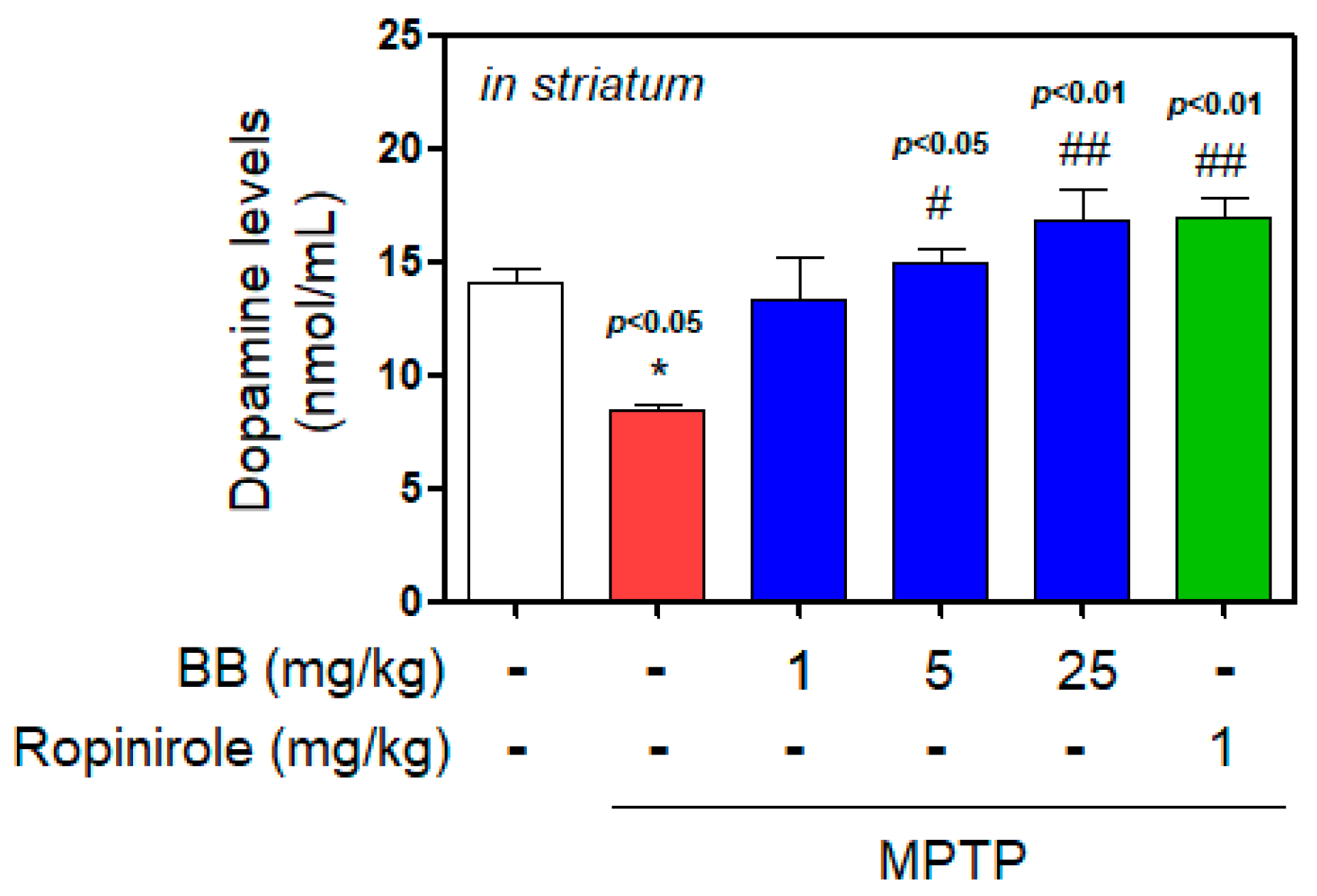
3.2. Effect of BB on MPTP-Induced Dopamine Depletion
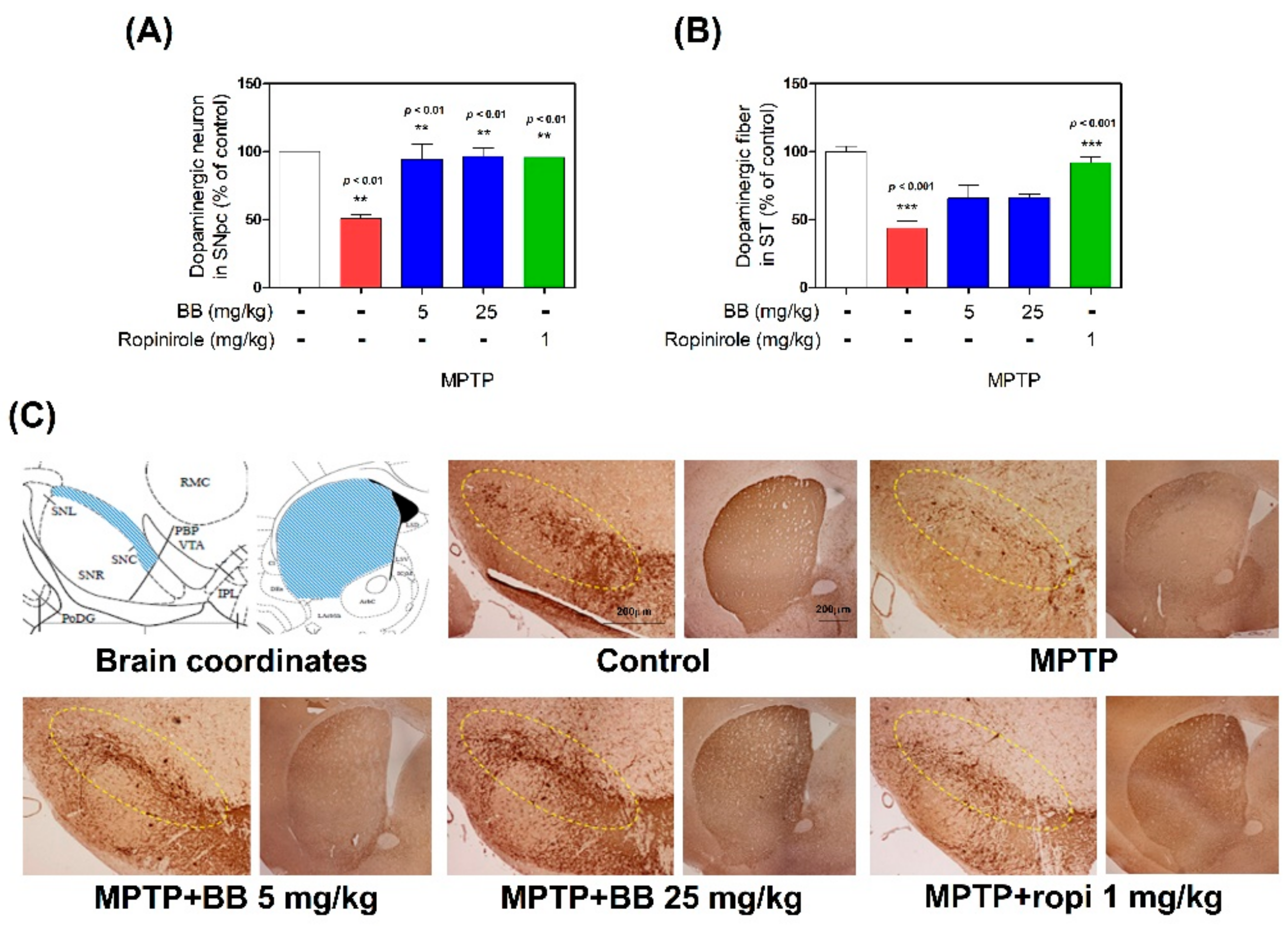
3.3. Effects of BB on MPTP-Induced Dopaminergic Neuronal Loss in SNpc and ST
3.4. Effects of BB on MPTP-Induced Expression Levels of Antioxidant Enzymes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The role of oxidative stress in neurodegenerative diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yu, S.; Zheng, Y.; Yang, H.; Zhang, J. Oxidative modification and its implications for the neurodegeneration of Parkinson’s disease. Mol. Neurobiol. 2017, 54, 1404–1418. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Park, G.; Kim, H.; Oh, D.S.; Kim, H.; Oh, M. In vitro and in vivo neuroprotective effects of walnut (Juglandis semen) in models of Parkinson’s disease. Int. J. Mol. Sci. 2016, 17, 108. [Google Scholar] [CrossRef] [PubMed]
- Sarrafchi, A.; Bahmani, M.; Shirzad, H.; Rafieian-Kopaei, M. Oxidative stress and Parkinson’s disease: New hopes in treatment with herbal antioxidants. Curr. Pharm. Des. 2016, 22, 238–246. [Google Scholar] [CrossRef]
- Coldwell, M.C.; Boyfield, I.; Brown, T.; Hagan, J.J.; Middlemiss, D.N. Comparison of the functional potencies of Ropinirolee and other dopamine receptor agonists at human D2 (long), D3 and D4.4 receptors expressed in Chinese hamster ovary cells. Br. J. Pharmacol. 1999, 127, 1696–1702. [Google Scholar] [CrossRef]
- Fukuzaki, K.; Kamenosono, T.; Nagata, R. Effects of Ropinirolee on various parkinsonian models in mice, rats, and cynomolgus monkeys. Pharmacol. Biochem. Behav. 2000, 65, 503–508. [Google Scholar] [CrossRef]
- Hu, M.; Yu, Z.; Wang, J.; Fan, W.; Liu, Y.; Li, J.; Xiao, H.; Li, Y.; Peng, W.; Wu, C. Traditional uses; origins; chemistry and pharmacology of Bombyx batryticatus: A review. Molecules 2017, 22, 1779. [Google Scholar] [CrossRef]
- Kwon, H.C.; Lee, K.C.; Cho, O.R.; Jung, I.Y.; Cho, S.Y.; Kim, S.Y.; Lee, K.R. Sphingolipids from Bombycis Corpus 101A and their neurotrophic effects. J. Nat. Prod. 2003, 66, 466–469. [Google Scholar] [CrossRef]
- Kwon, H.C.; Jung, I.Y.; Cho, S.Y.; Cho, O.R.; Yang, M.C.; Lee, S.O.; Hur, J.Y.; Kim, S.Y.; Yang, J.B.; Lee, K.R. Phospholipids from Bombycis corpus and their neurotrophic effects. Arch. Pharma. Res. 2003, 26, 471–477. [Google Scholar] [CrossRef]
- Hu, M.; Liu, Y.; He, L.; Yuan, X.; Peng, W.; Wu, C. Antiepileptic Effects of Protein-Rich Extract from Bombyx batryticatus on Mice and Its Protective Effects against H2O2-Induced Oxidative Damage in PC12 Cells via Regulating PI3K/Akt Signaling Pathways. Oxid. Med. Cell. Longev. 2019, 2019, 7897584. [Google Scholar] [CrossRef]
- Park, G.; Park, Y.J.; Yang, H.O.; Oh, M.S. Ropinirole protects against 1-methyl-4-phenyl-1; 2; 3; 6-tetrahydropyridine (MPTP)-induced neurotoxicity in mice via anti-apoptotic mechanism. Pharmacol. Biochem. Behav. 2013, 104, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Lee, S.H.; Oh, D.S.; Kim, Y.U. Melatonin inhibits neuronal dysfunction-associated with neuroinflammation by atopic psychological stress in NC/Nga atopic-like mouse models. J. Pineal Res. 2017, 63, e12420. [Google Scholar] [CrossRef] [PubMed]
- Rozas, G.; López-Martın, E.; Guerra, M.; Labandeira-Garcıa, J. The overall rod performance test in the MPTP-treated-mouse model of Parkinsonism. J. Neurosci. Methods 1998, 83, 165–175. [Google Scholar] [CrossRef]
- Hauser, D.N.; Hastings, T.G. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease and monogenic parkinsonism. Neurobiol. Dis. 2013, 51, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Sherer, T.B.; Betarbet, R.; Greenamyre, J.T. Environment; mitochondria; and Parkinson’s disease. Neurosci. 2002, 8, 192–197. [Google Scholar]
- Daubner, S.C.; Le, T.; Wang, S. Tyrosine hydroxylase and regulation of dopamine synthesis. Arch. Biochem. Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef]
- Darmopil, S.; Muñetón-Gómez, V.C.; De Ceballos, M.L.; Bernson, M.; Moratalla, R. Tyrosine hydroxylase cells appearing in the mouse striatum after dopamine denervation are likely to be projection neurones regulated by L-DOPA. Eur. J. Neurosci. 2008, 27, 580–592. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef]
- Fiskum, G.; Starkov, A.; Polster, B.M.; Chinopoulos, C. Mitochondrial mechanisms of neural cell death and neuroprotective interventions in Parkinson’s disease. Ann. N. Y. Acad. Sci. 2003, 991, 111–119. [Google Scholar] [CrossRef]
- Knight, J.A. Reactive oxygen species and the neurodegenerative disorders. Ann. Clin. Lab. Sci. 1997, 27, 11–25. [Google Scholar]
- Chao, C.; Lee, E.H. Neuroprotective mechanism of glial cell line-derived neurotrophic factor on dopamine neurons: Role of antioxidation. Neuropharmacology 1999, 38, 913–916. [Google Scholar] [CrossRef]
- Albarracin, S.L.; Stab, B.; Casas, Z.; Sutachan, J.J.; Samudio, I.; Gonzalez, J.; Gonzalo, L.; Capani, F.; Morales, L.; Barreto, G.E. Effects of natural antioxidants in neurodegenerative disease. Nutr. Neurosci. 2012, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Barber, S.C.; Shaw, P.J. Oxidative stress in ALS: Key role in motor neuron injury and therapeutic target. Free Radic. Biol. Med. 2010, 48, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Dexter, D.; Carter, C.; Wells, F.; Javoy-Agid, F.; Agid, Y.; Lees, A.; Jenner, P.; Marsden, C.D. Basal lipid peroxidation in substantia nigra is increased in Parkinson’s disease. J. Neurochem. 1989, 52, 381–389. [Google Scholar] [CrossRef]
- Qi, D.; Ouyang, C.; Wang, Y.; Zhang, S.; Ma, X.; Song, Y.; Yu, H.; Tang, J.; Fu, W.; Sheng, L. HO-1 attenuates hippocampal neurons injury via the activation of BDNF–TrkB–PI3K/Akt signaling pathway in stroke. Brain Res. 2014, 1577, 69–76. [Google Scholar] [CrossRef]
- Chen, K.; Gunter, K.; Maines, M.D. Neurons overexpressing heme oxygenase-1 resist oxidative stress-mediated cell death. J. Neurochem. 2000, 75, 304–313. [Google Scholar] [CrossRef]
- Shi, Y.; Liang, X.C.; Zhang, H.; Wu, Q.L.; Qu, L.; Sun, Q. Quercetin protects rat dorsal root ganglion neurons against high glucose-induced injury in vitro through Nrf-2/HO-1 activation and NF-κB inhibition. Acta Pharmacol. Sin. 2013, 34, 1140. [Google Scholar] [CrossRef]
- Barone, E.; Di Domenico, F.; Sultana, R.; Coccia, R.; Mancuso, C.; Perluigi, M.; Butterfield, D.A. Heme oxygenase-1 posttranslational modifications in the brain of subjects with Alzheimer disease and mild cognitive impairment. Free Radic. Biol. Med. 2012, 52, 2292–2301. [Google Scholar] [CrossRef]
- Ross, D.; Kepa, J.K.; Winski, S.L.; Beall, H.D.; Anwar, A.; Siegel, D. NAD (P) H: Quinone oxidoreductase 1 (NQO1): Chemoprotection; bioactivation; gene regulation and genetic polymorphisms. Chem. Biol. Interact. 2000, 129, 77–97. [Google Scholar] [CrossRef]
- Zhao, F.; Wu, T.; Lau, A.; Jiang, T.; Huang, Z.; Wang, X.J.; Chen, W.; Wong, P.K.; Zhang, D.D. Nrf2 promotes neuronal cell differentiation. Free Radic. Biol. Med. 2009, 47, 867–879. [Google Scholar] [CrossRef]
- Jakel, R.J.; Townsend, J.A.; Kraft, A.D.; Johnson, J.A. Nrf2-mediated protection against 6-hydroxydopamine. Brain Res. 2007, 1144, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.C.; Vargas, M.R.; Pani, A.K.; Smeyne, R.J.; Johnson, D.A.; Kan, Y.W.; Johnson, J.A. Nrf2-mediated neuroprotection in the MPTP mouse model of Parkinson’s disease: Critical role for the astrocyte. Proc. Natl. Acad. Sci. USA 2009, 106, 2933–2938. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.-S.; Kim, J.-S.; Moon, B.C.; Ryu, S.M.; Lee, J.; Park, G. Batryticatus Bombyx Protects Dopaminergic Neurons against MPTP-Induced Neurotoxicity by Inhibiting Oxidative Damage. Antioxidants 2019, 8, 574. https://doi.org/10.3390/antiox8120574
Lim H-S, Kim J-S, Moon BC, Ryu SM, Lee J, Park G. Batryticatus Bombyx Protects Dopaminergic Neurons against MPTP-Induced Neurotoxicity by Inhibiting Oxidative Damage. Antioxidants. 2019; 8(12):574. https://doi.org/10.3390/antiox8120574
Chicago/Turabian StyleLim, Hye-Sun, Joong-Sun Kim, Byeong Cheol Moon, Seung Mok Ryu, Jun Lee, and Gunhyuk Park. 2019. "Batryticatus Bombyx Protects Dopaminergic Neurons against MPTP-Induced Neurotoxicity by Inhibiting Oxidative Damage" Antioxidants 8, no. 12: 574. https://doi.org/10.3390/antiox8120574
APA StyleLim, H.-S., Kim, J.-S., Moon, B. C., Ryu, S. M., Lee, J., & Park, G. (2019). Batryticatus Bombyx Protects Dopaminergic Neurons against MPTP-Induced Neurotoxicity by Inhibiting Oxidative Damage. Antioxidants, 8(12), 574. https://doi.org/10.3390/antiox8120574