1. Introduction
Diabetes mellitus (DM) poses a serious public health issue [
1]. As of 2017, there were 425 million diagnosed diabetics worldwide, and this number may increase up to 629 million in 2045 according to an International Diabetes Federation report [
2]. Typically, diabetes can be classified as type 1 diabetes, type 2 diabetes, gestational diabetes, or as other types with different etiologies. Type 2 diabetes, which is characterized by hyperglycemia, insulin resistance, and disturbance in lipid, protein, and carbohydrate metabolism, is the most often diagnosed type in adults (about 90% of adult diabetics) [
3,
4,
5,
6]. In patients with diabetes, a range of diabetic complications may develop, namely diabetic cardiomyopathy, myocardial insufficiency, retinopathy, nephropathy, neuropathy, and hyperlipidemia [
5,
7,
8]. However, the cardiovascular complications of diabetes are considered the major factor resulting in deaths of diabetic patients [
3,
7,
9,
10].
It is believed that diabetic complications, such as diabetic-related cardiac dysfunctions, are directly connected with redox imbalance and oxidative stress [
9,
11,
12,
13,
14]. Oxidative stress can be described as the lack of ability of the organism to neutralize free radicals, accompanied by overproduction of the latter. When the endogenous antioxidative system becomes insufficient in order to cope with the excess of free radicals, it could be helpful to aid antioxidative capabilities of the body with exogenous antioxidants [
15]. Numerous scientific reports indicate that exogenous antioxidants improved oxidative stress-related parameters in the serum and various tissues [
16,
17,
18,
19,
20,
21,
22], including cardiac tissue [
23,
24,
25,
26,
27], in animals with experimentally-induced diabetes. Phenolic acids—rosmarinic acid and sinapic acid—may be considered as such exogenous antioxidants.
Both rosmarinic acid and sinapic acid are compounds commonly occurring in plants. These phenolic acids can be delivered to the organism on a daily basis in numerous herbs, vegetables, and fruits. Rosmarinic acid can be found mainly in Lamiaceae plants, such as rosemary, peppermint, lemon balm, or sage [
28,
29], whereas sinapic acid occurs in citrus fruits, strawberries, and Brassicaceae vegetables such as broccoli, turnip, kale, or tronchuda cabbage [
30,
31].
Scientific literature shows that both rosmarinic and sinapic acids have a beneficial effect on parameters connected with the glucose metabolism in diabetic rats [
32,
33,
34,
35,
36,
37,
38]. Additionally, rosmarinic acid positively affects several oxidative stress-related parameters and lipid profile in rats with experimentally induced diabetes [
32,
33]. Both these phenolic acids reveal a favorable effect on lipid metabolism and parameters related to oxidative stress in the serum of rats in the early stage of estrogen deficiency [
39,
40]. What is more, there are reports indicating a beneficial effect of rosmarinic acid and sinapic acid on cardiac dysfunction in several experimental rodent models [
41,
42,
43,
44,
45].
Based on scientific reports, we hypothesize that disturbance in redox balance in the serum and cardiac tissue may develop in long-term hyperglycemia in type 2 diabetic rats. Moreover, we suppose that rosmarinic acid and sinapic acid could mitigate hyperglycemia and hyperlipidemia and they may act as antioxidants in cardiac tissue. Thus, this experiment was conducted in order to evaluate the effect of rosmarinic acid and sinapic acid on the parameters connected with glucose and lipid metabolism in the serum and on the markers of oxidative stress in the serum and cardiac tissue of female rats with type 2 diabetes induced with high-fat diet and streptozotocin.
2. Materials and Methods
2.1. Animals and Drugs
The experiment was carried out on sexually mature (3-month-old) female Wistar rats. The study was approved by the Local Ethics Commission in Katowice, Poland (permission numbers: 38/2015, 148/2015 and 66/2016). The rats were provided by Center of Experimental Medicine, Medical University of Silesia (Katowice, Poland). The animals were kept in standard laboratory conditions (EU Directive 2010/63).
The following drugs and substances were used in the experiment: streptozotocin (STZ, Cayman Chemical Company, Ann Arbor, MI, USA); ketamine (Ketamina 10%, Biowet Puławy Sp. z o. o., Puławy, Poland); xylazine (Xylapan, Vetoquinol Biowet, Gorzów Wlkp., Poland); rosmarinic acid (Sigma-Aldrich, St. Louis, MO, USA); and sinapic acid (Sigma-Aldrich, St. Louis, MO, USA).
During a one-week acclimation period the animals were randomly assigned to six experimental groups (n = 10):
Control, non-diabetic rats (C group)
High-fat diet and streptozotocin (HFD + STZ)-induced type 2 diabetic control rats (DM2 group)
HFD + STZ diabetic rats administered orally with rosmarinic acid at a dose of 10 mg/kg (RA10 group)
HFD + STZ diabetic rats administered orally with rosmarinic acid at a dose of 50 mg/kg (RA50 group)
HFD + STZ diabetic rats administered orally with sinapic acid at a dose of 5 mg/kg (SA5 group)
HFD + STZ diabetic rats administered orally with sinapic acid at a dose of 25 mg/kg (SA25 group)
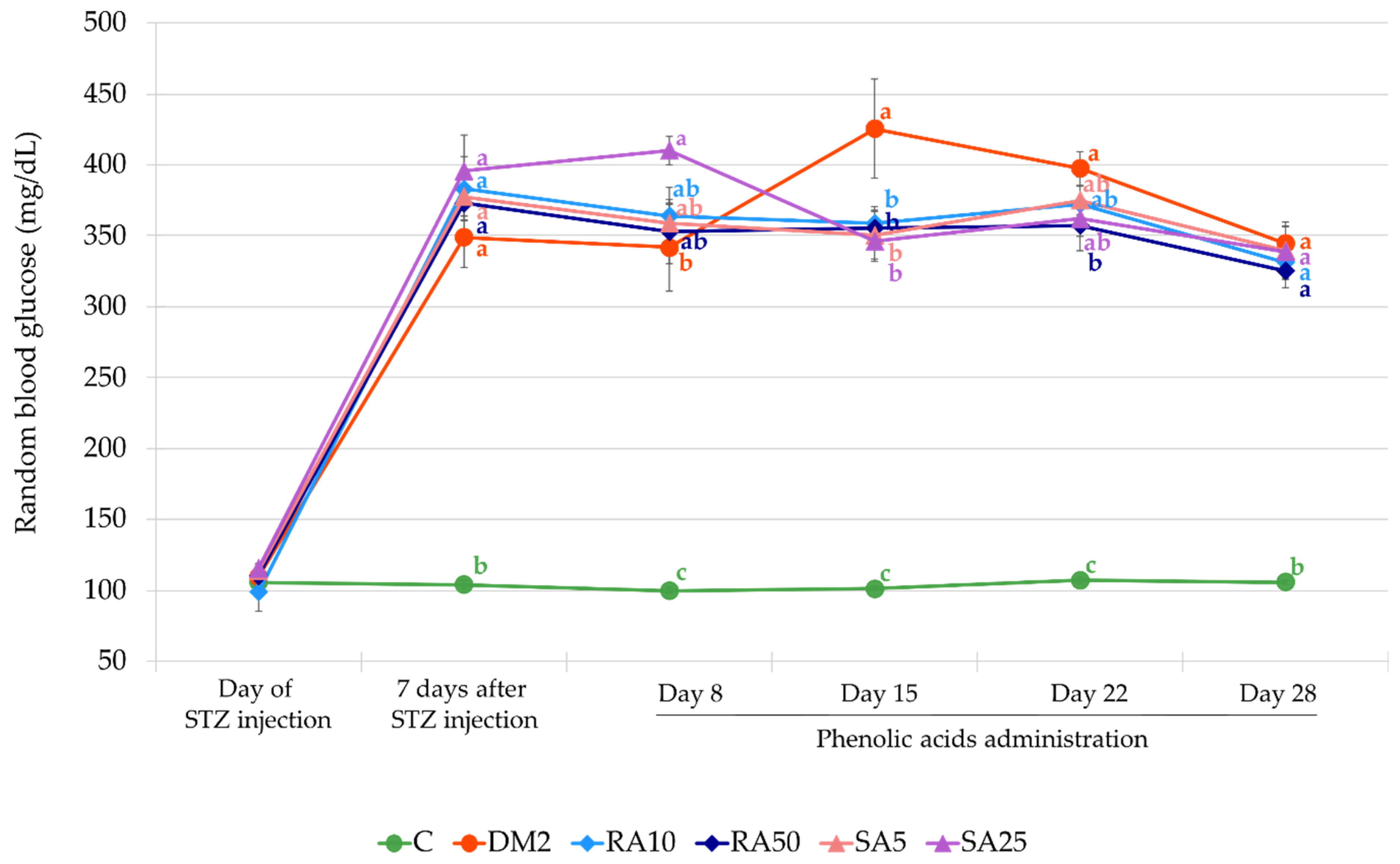
The animals were weighted at the beginning of the experiment, and then once a week. The last weight record was taken before an overnight fasting. From the day of STZ injection, a random blood glucose level from tail capillary vessels (obtained by cutting the end of the tail) was measured once a week by MicroDot glucometer equipped with test strips (Cambridge Sensor USA, Plainfield, IL, USA). During the whole experiment the animals had unlimited water supply and appropriate chow: rats in the C group were fed with a standard laboratory chow (Labofeed B, Wytwórnia Pasz “Morawski”, Kcynia, Poland) and the rats in the DM2, RA10, RA50, SA5, and SA25 groups were fed for 2 weeks before STZ injection with high-fat diet (HFD) enriched with 32% of fat from lard (Wytwórnia Pasz “Morawski”, Kcynia, Poland). The HFD in these groups was continued till the end of the experiment. The overall duration of HFD in the groups with experimentally induced type 2 diabetes was 49 days. Before a single injection with streptozotocin (40 mg/kg intraperitoneally) the rats were fasted for 8 h. Streptozotocin was dissolved in 0.1 M citric buffer and 1 mL/kg of this solution was injected. The rats from the C group were injected with an appropriate volume of 0.1 M citric buffer only. Seven days after the STZ injection random blood glucose level was measured. If the result of this test was higher than 200 mg/dL, the animal was considered diabetic. Administration of rosmarinic acid and sinapic acid started 7 days after STZ injection and lasted for 28 days. The phenolic acids suspended in water (with addition of Tween 20 quantum satis, max. 1 µL of Tween 20 per 1 mL of water) were administered by gavage once a day. Control rats from the C and DM2 groups received water with Tween 20 by gavage at the same volume as the rats from the rosmarinic acid and sinapic acid-treated groups received the suspension of phenolic acids (2 mL/kg). The timeline of the in vivo part of the experiment is presented in the
Figure 1.
24 h after the administration of the last dose of phenolic acids and overnight fasting, rats were euthanized by injection of ketamine and xylazine mixture and cardiac exsanguination. In order to separate the serum required for biochemical assays, the blood obtained from the heart was centrifuged. Vital organs such as the heart, liver, right kidney and the uterus were excised from the animals and weighted. Then, each heart was homogenized separately in phosphate buffered saline (PBS), pH 7.4 (10% w/v), and centrifuged (10,000× g, 15 min, +4 °C). The obtained supernatant was used for further examinations. All assays were measured in Tecan Infinite M200 PRO plate reader with Magellan 7.2 software (Tecan Austria, Grödig, Austria).
2.2. Biochemical Assays in the Serum
The levels of the estradiol and interleukin 18 (IL-18) were assessed with ELISA kits (DiaMetra, Segrate-Milano, Italy for estradiol and Cloud-Clone Corp. Houston, TX, USA for IL-18). Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) activities as well as uric acid and urea levels were measured spectrophotometrically with Biosystems kits (Costa Brava, Barcelona, Spain); the creatinine was assayed with Pointe Scientific Inc. kit (Canton, MI, USA).
2.3. Parameters Connected with Glucose Homeostasis in the Serum and Cardiac Tissue
Glucose and fructosamine levels in the serum were evaluated with Pointe Scientific Inc. kits. Insulin levels in the serum were measured with the BioVendor ELISA kit (Brno, Czech Republic). The homeostasis model assessment of insulin resistance (HOMA-IR) was calculated according to the following Equation:
while the quantitative insulin sensitivity check index (QUICKI) was calculated as follows:
Advanced glycation end products (AGEs) in both the serum and cardiac tissue were assayed with ELISA method (Cell Biolabs, San Diego, CA, USA).
2.4. Lipid Profile Assessment in the Serum
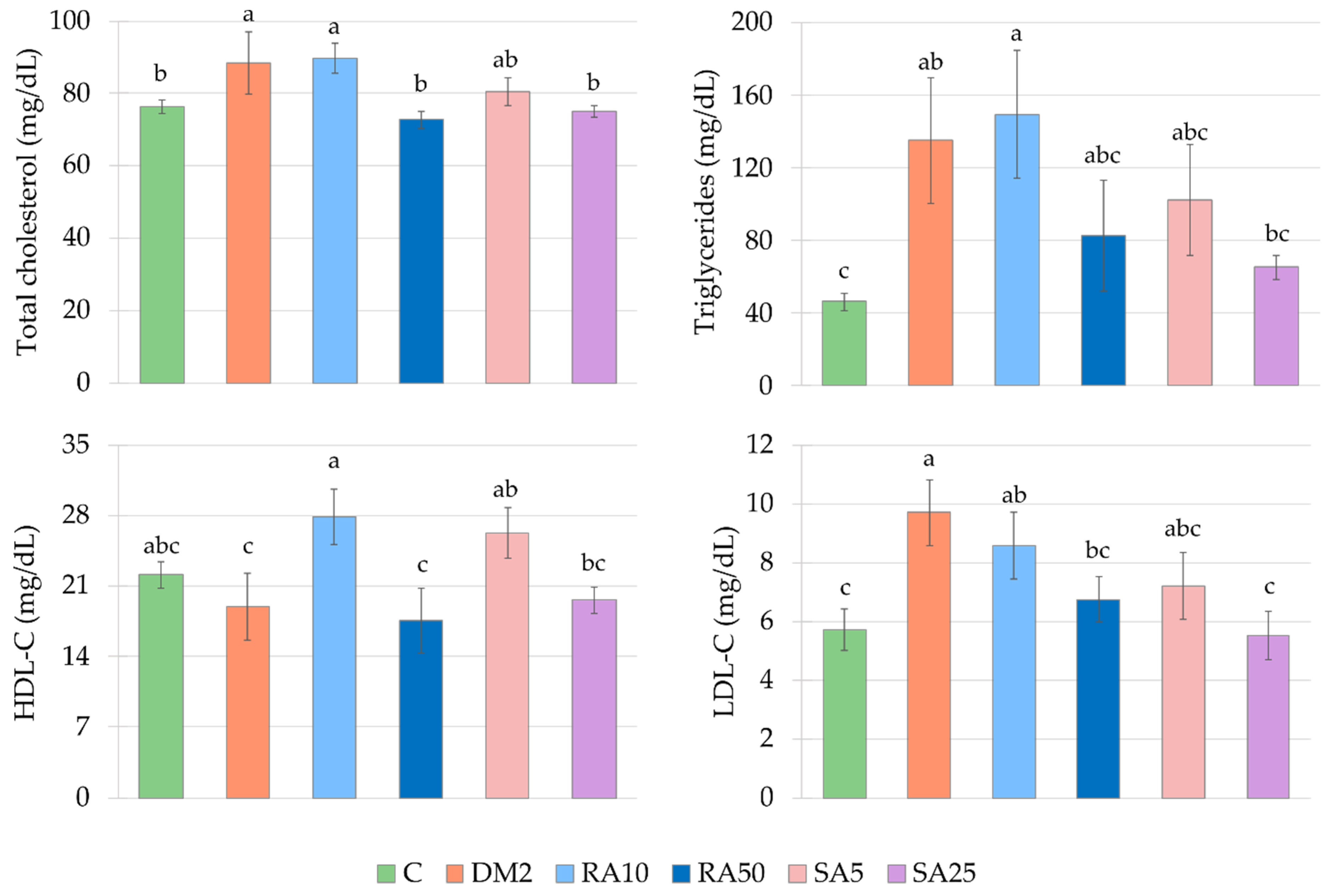
Total cholesterol, triglycerides, low-density lipoprotein cholesterol (LDL-C), and high-density lipoprotein cholesterol (HDL-C) levels were assayed spectrophotometrically with Pointe Scientific Inc kits (Canton, MI, USA).
2.5. Oxidative Stress-Related Parameters Assessment in the Serum and Cardiac Tissue
Activity of superoxide dismutase (SOD) and catalase (CAT) was measured in both the serum and cardiac tissue. Additionally, glutathione peroxidase (GPx) activity was evaluated in cardiac tissue. All the enzymes were assayed with the use of Cayman kits (Cayman Chemical, Ann Arbor, MI, USA) and their activity was expressed per 1 mg of protein. The protein level was assessed with the Pointe Scientific Inc kit (Canton, MI, USA).
Non-protein sulfhydryl groups (NPSH) levels in the serum and cardiac tissue were examined according to the protocol proposed by Sedlak and Lindsay [
46]. In this method, a colorful product of reaction between sulfhydryl groups and 5,5’-dithiobis-(2-nitrobenzoic acid) (Sigma-Aldrich, St Louis, MO, USA) is measured at 412 nm. The standard curve was prepared with reduced glutathione (Sigma-Aldrich, St Louis, MO, USA).
Thiobarbituric acid reactive substances (TBARS) levels in the serum and cardiac tissue were evaluated with the Buege and Aust [
47] method, in which thiobarbituric acid reacts with lipid peroxidation products, and the colorful product of this reaction is measured spectrophotometrically at 535 nm. Obtained results were calculated with the use of the extinction coefficient = 1.56 × 10
5/M cm.
The level of advanced oxidation protein products (AOPP) was obtained according to the Witko–Sarsat method [
48], and chloramine T (Sigma-Aldrich, St Louis, MO, USA) was used for standard curve. The absorbance was read at 340 nm. In the serum, the AOPP level was expressed as 1 nmol of chloramine T equivalent per 1 mL and in the cardiac tissue as 1 nmol of chloramine T equivalent per 1 mg of protein.
2.6. Statistical Analyses
The results are presented as arithmetical means ± SEM. To evaluate statistical significance, one-way ANOVA followed by Fisher’s Least Significant Difference (LSD) post-hoc test was used (Statistica 13 software, StatSoft, Kraków, Poland). The results were considered significant if
p < 0.05. Additionally, data obtained from the cardiac tissue and serum were subjected to principal component analysis (PCA) using PAST 3.26 software [
49]. The PCA data was subjected to MANOVA (Statistica 13;
Appendix A).
The number of rat subjects in each experimental group was equal at the beginning of the experiment (n = 10). However, in the DM2 control and phenolic acids-treated groups the number of rats varied based on the diabetes onset—only the rats classified as type 2 diabetic were included in analyses. After the classification, the final numbers were as follows: DM2, n = 7; RA10, n = 8; RA50, n = 6; SA5, n = 7; and SA25, n = 7, whereas C remained at n = 10.
4. Discussion
In the presented study we aimed to evaluate the effect of rosmarinic acid and sinapic acid on oxidative stress-related parameters in the cardiac tissue as well as parameters connected with glucose homeostasis, lipid profile and oxidative stress in the serum of the rats with experimentally induced type 2 diabetes. Among the existing mice and rat models of diabetes, the model in which the animals are treated with a single injection of relatively high dose of streptozotocin is the most frequently used [
50,
51]. This model is characterized by a very high glucose level, polydipsia, polyuria, and, in consequence, dehydration and general exhaustion. The damage of the vital organs, which is a result of such a severe hyperglycemia, reflects type 1 diabetes rather than the type 2. There are several genetic models of the type 2 diabetes in rodents, such as: db/db mice or Zucker Diabetic Fatty (ZDF) rats (fa/fa), Goto Kakizaki (GK) rats, and Otsuka Long Evans Tokushima (OLET) rats. However, these genetic models are imperfect. This is due to the fact that insulin resistance, obesity and hyperglycemia in these models are the result of a gene mutation (e.g., leptin receptor OB-r), which can rarely be observed in humans [
52]. Therefore, it can be concluded, that the model reflecting the changes observed in the type 2 diabetes in humans in the most adequate manner is a model in which a high-fat diet is combined with a single injection of a low dose of streptozotocin [
52,
53]. It should be noted that the majority of studies describing the effect of tested substances on parameters affected by both the type 1 and type 2 diabetes is based on the experiments conducted on male laboratory animals [
17,
19,
20,
21,
22,
23,
24,
25,
32,
33,
34,
35,
37,
38,
50,
51,
54,
55,
56,
57,
58,
59,
60,
61,
62]. Following the suggestion of The National Institutes of Health (NIH) to use female animals in the laboratory studies [
63] and reports about sex-dependent metabolic differences [
64,
65], our experiment was conducted on female rats.
In our experiment, the diabetes was induced by a single injection of streptozotocin (40 mg/kg intraperitoneally) preceded by a 14-day treatment with a high-fat diet (32% of fat from lard). After streptozotocin injection, the high-fat diet was continued in the diabetic rats till the end of the experiment, i.e., for additional 35 days. This treatment induced disturbance in the glucose and lipid metabolism. Similarly to other reports, we observed increased random blood glucose level as well as the levels of fasting serum glucose, serum fructosamine, total cholesterol, and triglycerides. Moreover, induction of the type 2 diabetes resulted in the HOMA-IR increase and QUICKI decrease [
32,
35,
54,
56,
57,
58,
66]. There were also changes in the oxidative stress-related parameters in the serum. Diabetes induced an increase in the SOD and CAT activities and in the level of the lipid peroxidation marker TBARS. The increase in the TBARS level is consistent with the other reports [
58,
59]. On the other hand, the reports on the effect of diabetes (including HFD + STZ-induced type 2 diabetes) on the activity of the antioxidant enzymes in the serum are incoherent. There are studies in which activity of these enzymes increases in the serum/plasma [
21,
55], and such which describe a decrease in the antioxidative enzymes activities [
22,
58,
60]. Among other things, this discrepancy may be a result of differences in the duration to free radical exposure [
67,
68]. It should be noted that in our study neither AOPP (a marker for protein oxidative damage) nor NPSH (of which reduced glutathione (GSH), an endogenous nonenzymatic antioxidant, is the main representative) changed after induction of the type 2 diabetes. Numerous studies indicate that in the course of diabetes the GSH level in the serum/plasma decreases and the level of protein damage markers increases [
21,
22,
32,
57]. Lack of such alterations observed in our study may result from the fact that in females, even in the type 2 diabetes, estrogens play protective role against oxidative stress [
69], and the estrogen level in rats in our study did not change significantly after induction of diabetes.
In this study the level of interleukin 18 (IL-18) was measured, since this interleukin is engaged in metabolic homeostasis and it is sex hormone-dependent [
70,
71]. The role of IL-18 in metabolic syndrome, obesity, or diabetes is not clear. On the one hand, in people with diabetes, metabolic syndrome, or obesity the IL-18 level is significantly higher than in healthy people [
72]. On the other hand, in mice with inhibited IL-18 synthesis or with deficiency of inflammasome NLPR1, which is responsible for IL-18 synthesis, obesity, insulin resistance and other metabolic syndrome symptoms were observed [
73,
74]. We observed that in type 2 diabetic rats, which demonstrated insulin resistance, the level of IL-18 was significantly lower than in control healthy rats. Our results contradict the studies describing the elevated level of IL-18 in the Otsuka Long-Evans Tokushima Fatty rats and fructose-fed rats [
75,
76]. These differences may result from sexual dimorphism, as these studies were conducted on male rats [
75,
76] and not on female rats. Such gender discrepancies in the plasma IL-18 level were previously observed in wild-type mice injected intraperitoneally with LPS [
77]. Another important factor which seems to affect the IL-18 level is diet. Russel et al. observed that the level IL-18 in the serum of ovariectomized rats depended on the isoflavone content in the chow [
78]. However, our study is consistent with another report, in which the IL-18 level in Spontaneously Diabetic Torii rats was lower than in control, non-diabetic rats [
79]. What is more, the authors of this study noticed, that IL-18 level differs in time after diabetes induction [
79]. Taking into consideration all abovementioned literature data it could be assumed that IL-18 level may depend on many factors, including gender, species, strain or genetic model of the animal used for research, time point in which the measurement was taken, or even diet composition.
In the presented study, rosmarinic acid and sinapic acid were administered to the type 2 diabetic rats at the doses of 10 and 50 mg/kg and 5 and 25 mg/kg, respectively. The lower doses of the tested phenolic acids correspond with the dietary achievable doses. The 5-fold higher doses were chosen in order to evaluate if they may have a better therapeutic effect than the dietary doses. The same doses for both the phenolic acids were used previously in ovariectomized rats. Both rosmarinic acid and sinapic acid lowered HOMA-IR in ovariectomized rats and rosmarinic acid additionally lowered fructosamine level in these rats [
39,
40]. In the current study, in which rats were in a permanent hyperglycemic state, neither rosmarinic acid nor sinapic acid affected the parameters connected with glucose metabolism. Even though the higher doses of phenolic acids lowered the fasting glucose level in the serum, the fructosamine level in the serum (a parameter which reflects long-term glycemia status [
80]), random blood glucose level or HOMA-IR were not decreased and QUICKI was not elevated. It should be noted that control of blood glucose level and monitoring the parameters representing long-term changes in glycemia (such as glycated hemoglobin A1c (HbA1c) or fructosamine) are a crucial element in prevention of diabetes complications [
81,
82,
83]. It has been proven that postprandial glucose as well as random glucose levels have a closer association to glycated hemoglobin than the level of fasting plasma glucose [
84,
85,
86]. As observed in a study conducted in type 2 diabetic patients, a greater emphasis should be put on regulation of the postprandial glucose level than on fasting glucose level, as it could produce better outcomes in general diabetes therapy e.g., lowering HbA1c [
87].
Rosmarinic acid and sinapic acid administered to the type 2 diabetic rats at the higher doses (50 and 25 mg/kg respectively) lowered total cholesterol in the serum of the rats, similarly to rats in the early stage of estrogen deficiency [
39,
40]. Moreover, in our present study the higher doses of phenolic acids lowered the LDL-C in the serum of the type 2 diabetic rats. The beneficial effect of rosmarinic acid on lipid profile in rats with diabetes induced with high-fat diet and streptozotocin was previously shown [
32,
33]. In contrast to our results, in these works was shown that rosmarinic acid revealed a beneficial effect on parameters describing glucose metabolism and oxidative stress [
32,
33,
35]. In two of these studies, administration of rosmarinic acid at a dose of 100 mg/kg resulted in a decrease in glycated hemoglobin and HOMA-IR [
32,
35] and in the third study, rosmarinic acid administered at a dose of 30 mg/kg lowered the AGEs level in the heart, liver, and kidney [
33]. There is also a study in which sinapic acid administered at the dose of 25 mg/kg to the HFD + STZ-induced diabetic rats demonstrated a positive effect on glucose metabolism parameters such as: HbA1c or HOMA-IR [
37]. However, it should be highlighted that all the aforementioned studies [
32,
33,
35,
37] were conducted on male rats and not on female rats like in our study. Therefore, it is possible that inconsistency in rosmarinic acid action results from sex differences. To the best of our knowledge, there were no studies describing the effect of sinapic acid on parameters connected with lipid profile in diabetic animals. Positive effect of sinapic acid on lipids level in the serum/plasma was observed in different experimental models, such as isoproterenol-induced myocardial infarcted rats [
44] or L-NAME-induced hypertensive rats [
43].
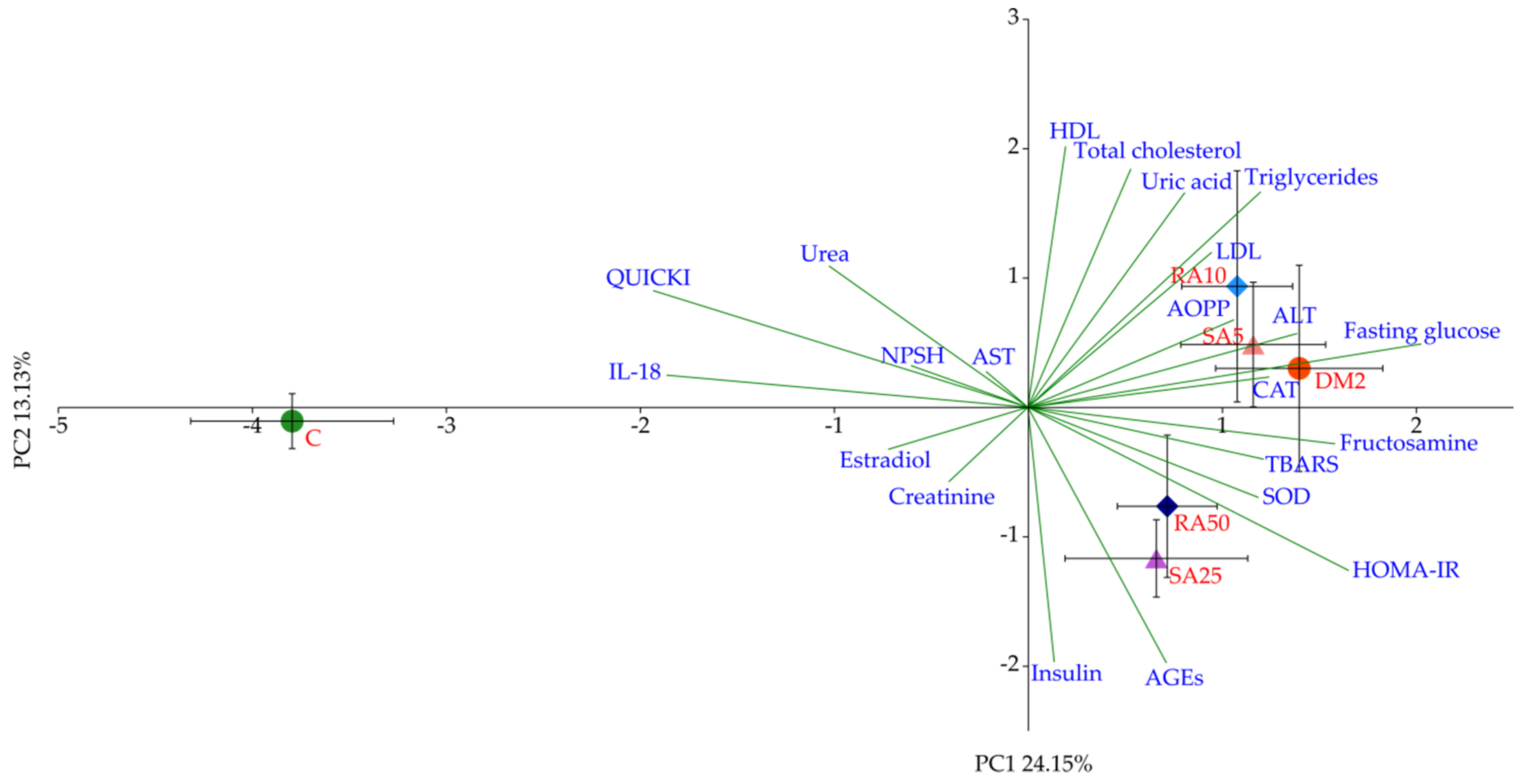
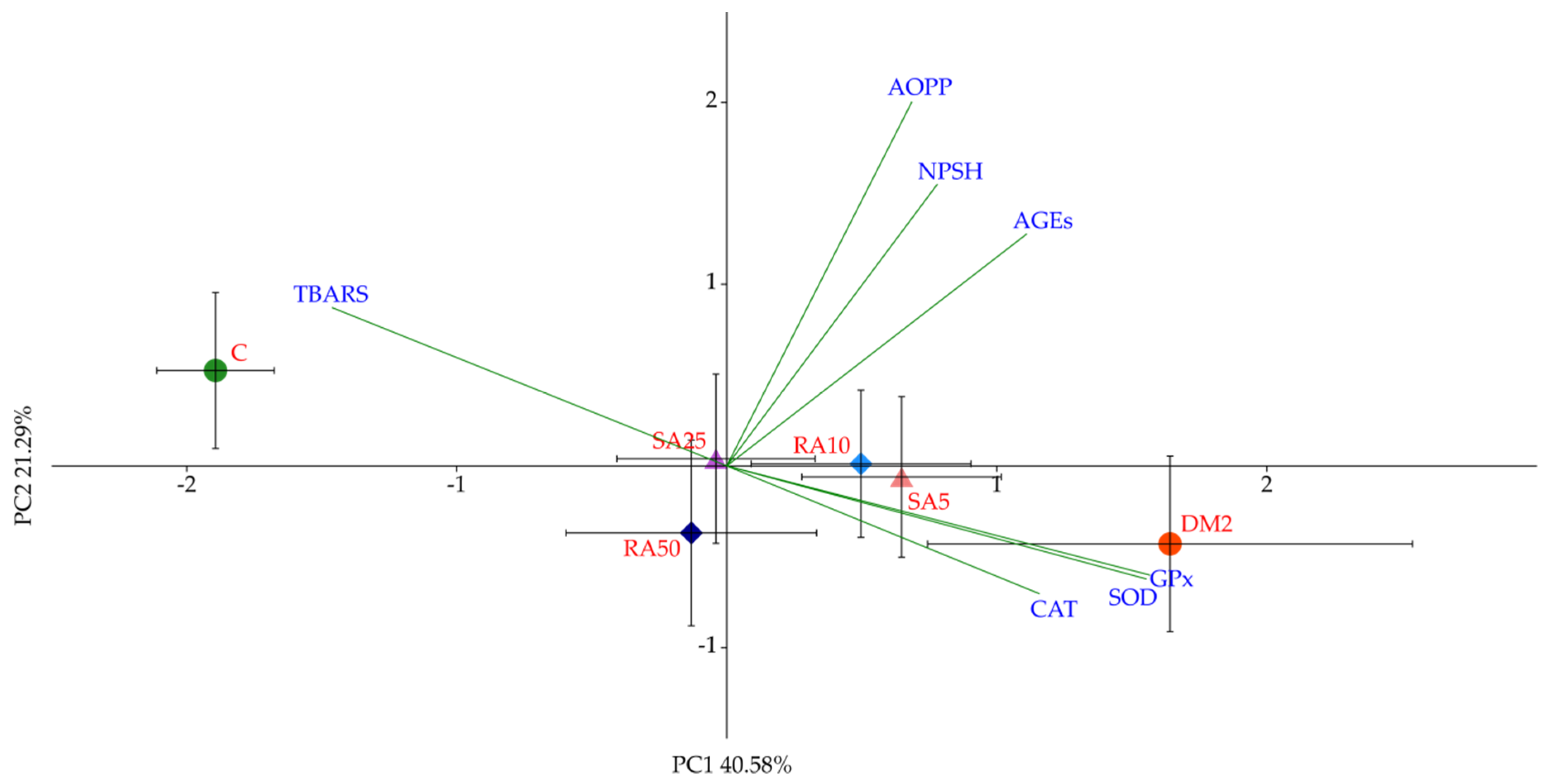
Based on the PCA it was determined that the variables predominantly responsible for the variance observed between the C group and all of the diabetic groups of rats were fasting glucose, fructosamine, HOMA-IR, QUICKI, and IL-18. There were no differences in these variables after the phenolic acids administration, thus no separation between these groups (RA10, RA50, SA5, and SA25) or DM2 was observed in the PCA. Even though there were minor differences in individual lipid profile markers, they were too weak to demonstrate the visible effect on the PCA, as much stronger effects were related to glucose metabolism. It is worth noting that random blood glucose measured in the capillary vessels was not affected after the phenolic acid administration for 28 days. This is in agreement with the PCA results.
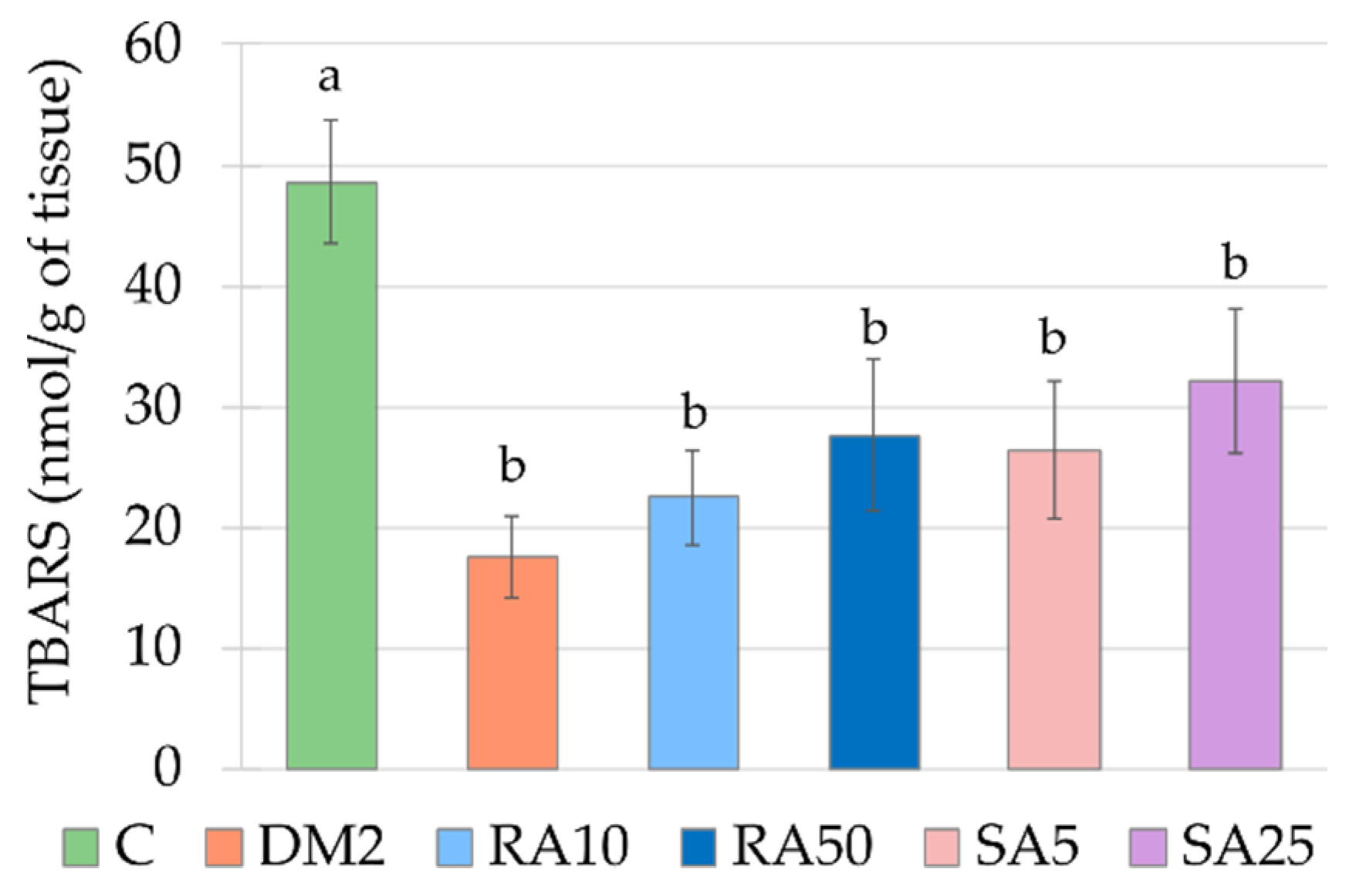
The results for oxidative stress parameters obtained from the cardiac tissue of the HFD + STZ-induced type 2 diabetic rats are surprising. The TBARS level in this tissue was significantly lower than in the control rats. The majority of the authors reports that the lipid peroxidation markers level increases in the heart during diabetes induced with streptozotocin alone [
26,
51] or with high-fat diet combined with streptozotocin [
27,
33,
88]. Only few publications indicate that TBARS level does not change or decreases in the cardiac tissue in diabetic rodents [
61,
89]. The lowered TBARS level in the heart was observed by the authors, who fed the rats with high-fat diet (with 30% of fat from palm oil) for 11 weeks [
90]. These authors explain the result by changes in polyunsaturated fatty acids (PUFA) profile in phospholipids of the heart membrane—there was a significant increase in arachidonic acid (AA) in the heart membrane of rats on palm oil-rich diet, compared to the membrane of rats fed with standard chow [
90]. In our study, rats were fed with diet enriched with 32% of fat from lard. The same research team who examined palm oil-rich diet in another study tested the changes in PUFA profiles after lard-rich diet. Similarly to palm oil-rich diet, the diet rich in lard also effected in an increase of AA in the heart membrane [
91]. The authors suggest that AA has a strong effect on vessel dilatation and its accumulation may promote coronary flow, which can result in an increased mechanical function of the heart [
90].
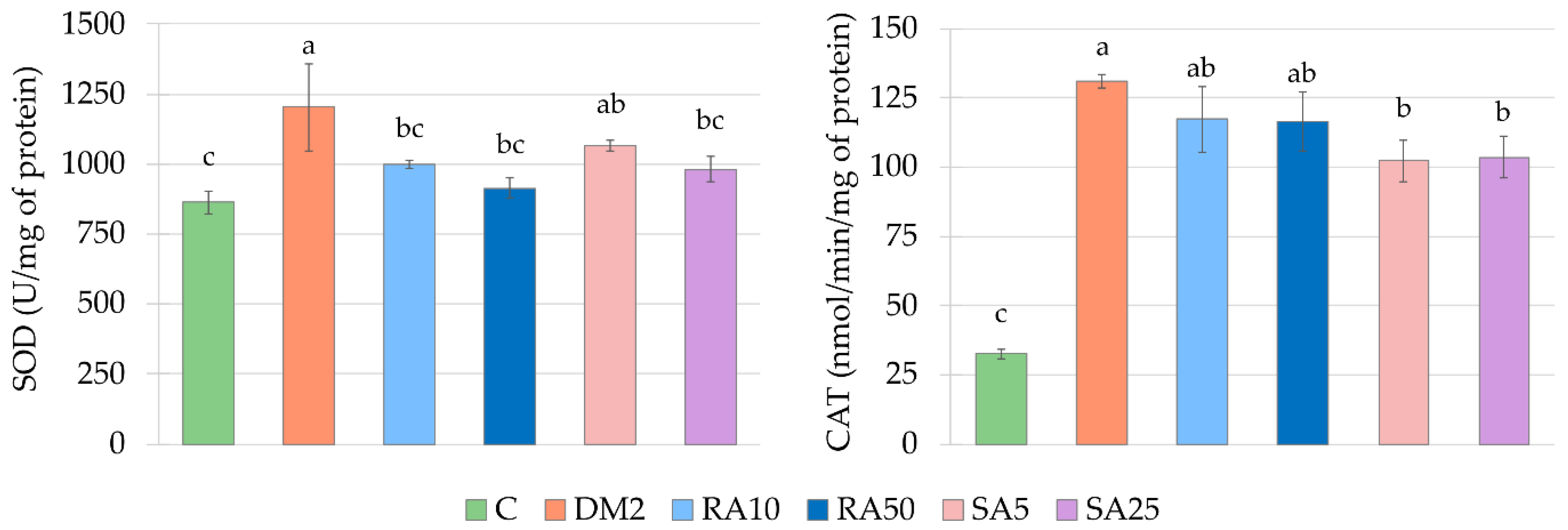
In the cardiac tissue, similarly to the serum, the activity of antioxidative enzymes was elevated. These results are in line with some other reports [
26,
61], but there are also reports indicating a decrease in antioxidative enzymes activity in the heart of diabetic animals [
24,
33,
58,
62]. In the study conducted by Okutan et al. regarding the effect of caffeic acid phenethyl ester on oxidative stress markers in the heart it was shown that similarly to our results, the activities of SOD and CAT were elevated in diabetic rats, while GPx activity was unchanged when compared to the non-diabetic animals. After administration of caffeic acid phenethyl ester, the activities of SOD and CAT was lowered. The authors concluded that administration of this derivative of phenolic acid reduced the diabetes-induced oxidative stress in the heart [
26]. Therefore, based on these findings, we also may conclude that lowering of SOD activity by both the doses of rosmarinic acid and the higher dose of sinapic acid, as well as lowering of CAT activity by both the doses of sinapic acid indicate a positive effect of these phenolic acids on cardiac tissue of type 2 diabetic rats.
In our experiment we have observed a diverse effect of individual phenolic acids on oxidative stress parameters in the cardiac tissue. Furthermore, for each compound there were discrepancies in the effects of individual doses on specific markers of oxidative stress. However, the multivariate analysis performed on the cardiac tissue-derived variables has confirmed a negligible or no effect of phenolic acids when administered in dietary doses of 10 mg/kg and 5 mg/kg for rosmarinic and sinapic acid, respectively. However, it should be noted that higher doses of the aforementioned compounds did beneficially affect the oxidative stress markers, as evidenced by the PCA. These differences were masked in individual ANOVAs. This might be due to the fact that classic analysis of variance takes into consideration only quantitative characteristics of measured variables, whereas the specifics of the PCA introduce a qualitative component to the study, as the effect (weight) of individual variables on the total observed variation between experimental groups along PC1 and PC2 axes is not equal [
92].
In the current research we focused on biochemical parameters in the serum and oxidative stress markers in the serum and cardiac tissue in the female diabetic rats. Due to this fact, this study has several limitations. Firstly, we did not conduct histological analyses or examine the molecular mechanisms underlying the action of tested phenolic acids in the cardiac tissue. We did not evaluate the effect of diabetes and phenolic acids administration on the markers connected to inflammation (e.g., IL-1β, IL-6, IL-10, IL-18, tumor necrosis factor α), apoptosis (for instance caspase 3 or p53), or cellular function (such as adenosine monophosphate-activated protein kinase, mitochondrial transcription factor A, or phosphoinositide 3-kinase) in the cardiac tissue. Secondly, further studies regarding rosmarinic acid or sinapic acid should include the cardiac mechanical function. Finally, in order to find out whether sex hormone environment is an important factor in the response to phenolic acids treatment, the next experiment should be conducted on both male and female rats.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








