1. Introduction
The basic cause of obesity is the imbalance between energy intake and energy expenditure. Accumulation of fat tissue, especially the accumulation of visceral fat tissue, is a possible primary cause of “metabolic syndrome”, which is the combination of multiple cardiovascular risk factors such as dyslipidemia, glucose intolerance, and hypertension. A number of epidemiological studies have reported positive associations between fat intake, level of blood cholesterol, and risk of coronary heart disease (CHD) and CHD mortality. Dyslipidemia and oxidative stress, which are closely related to diabetes, hypertension, obesity [
1,
2,
3,
4], and cancer [
5], might be consequences and causes of the high mortality of humans and animals from cardiovascular diseases [
5,
6]. Nutritional and pharmacological preparations that affect such physiological conditions, which accompany dyslipidemia, elevated plasma triglyceride levels, total low density lipoprotein (LDL) and very-low-density lipoprotein (VLDL) cholesterol, and a low level of high density lipoprotein (HDL) cholesterol, can regulate lipid metabolism and be effective in reducing the risk of lipid disorders, obesity, cardiovascular diseases, and consequent pathological complications [
7]. Recently, there has been renewed interest in the use of herbal bee products in the preventive treatment of many different chronic diseases, such as osteoporosis, cancer, psoriasis, and diabetes [
4,
5,
6,
7,
8,
9,
10,
11]. The advantages of using natural preparations are easy availability, better usability, lack of adverse effects, and economic cost-effectiveness. Propolis has been greatly popular as an agent in traditional medicine and as a food dietary supplement for human health in the world [
12,
13]. Propolis is produced by honeybees using collected extracts from leaves, buds, and exudates of various plant floras [
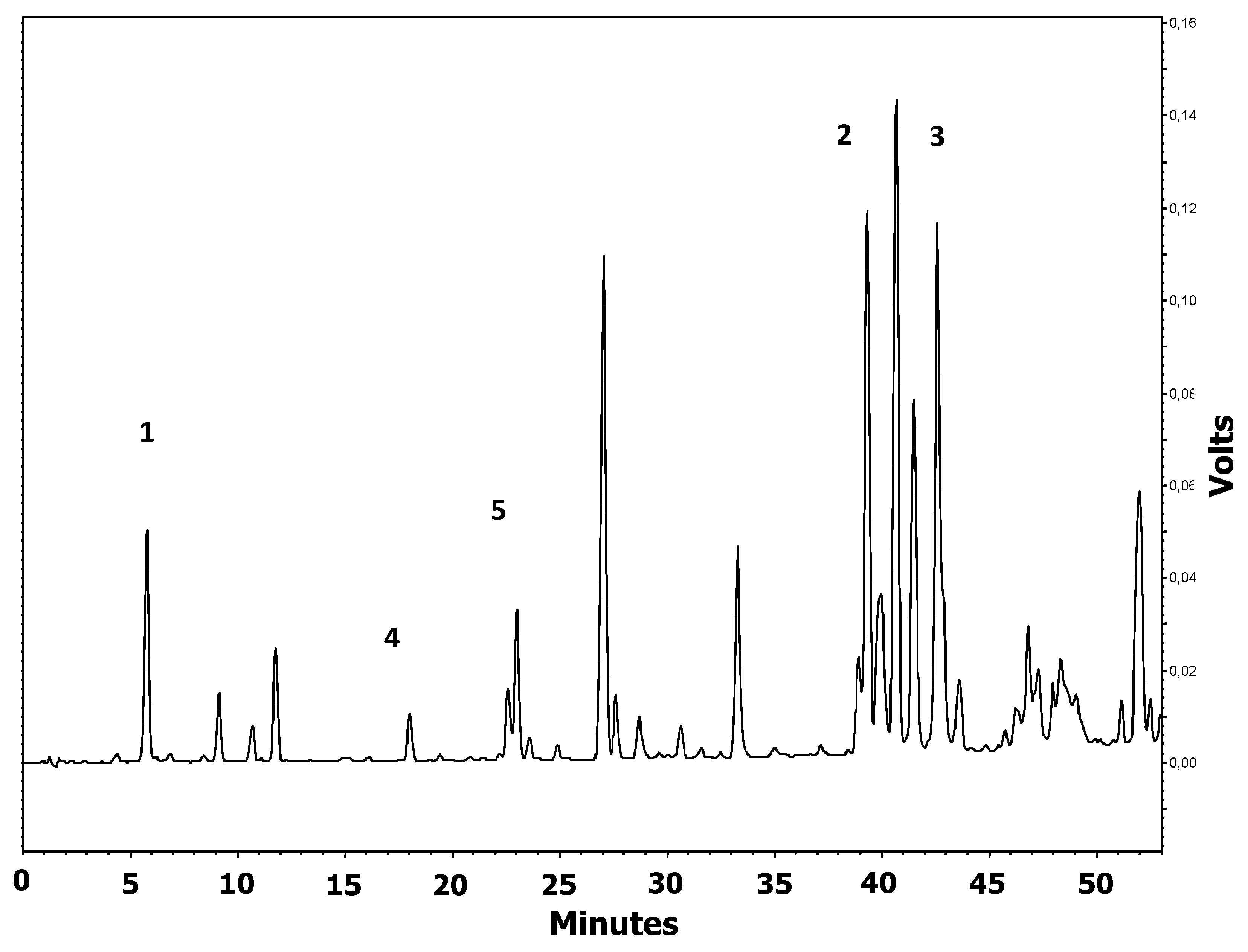
14]. Its antibacterial, antiviral, antitumor, anti-inflammatory, anticancer, antiallergic, antilipidemic, and vasodilatory actions, as well as its hepatoprotective and immunomodulatory effects, have been reported [
13,
15,
16]. In addition, flavonoids from propolis can inhibit lipid peroxidation, platelet aggregation, capillary permeability and fragility, and the activity of enzyme systems including cyclooxygenase and lipooxygenase [
5,
8,
10]. More than 300 molecular species, such as alkaloids, aromatic acids and their esters, free acids and their esters, phenolic acids and their esters, phenolic aldehydes, alcohols and ketones, sesquiterpenes, quinones, flavonoids, minerals, and vitamins are isolated from propolis [
17,
18], and their ratios and concentration depend on geographical origin. Flavonoids and phenolic compounds have strong antioxidant properties and the ability to protect cells from reactive radicals and protect important biological molecules, such as lipids, proteins, and DNA. Flavonoids can be aglycones, glycosides, and methylated derivatives, and their antioxidant activity depends on the arrangement of functional groups around the nuclear structure—in particular, the total number of hydroxyl groups [
17,
18]. For example, the hydroxyl configuration of the B ring is important for the removal of reactive oxygen species (ROS) and reactive nitrogen species (RNS) due to hydrogen and electron donation to hydroxyl, peroxyl, and peroxynitrite radicals, which stabilizes them and produces stable flavonoid radicals. It is believed that aglycones are more potent antioxidants than the corresponding glycosides, and that the degree of polymerization increases the antioxidant capacity, which is important for their biological and pharmacological efficacy [
3,
5,
19].
In spite of the multitude of ethnomedical applications against various pathophysiological conditions, we found scarce data in literature relating to the effect of propolis on obesity, plasma lipid profiles, and atherogenicity in a high-fat diet-induced obesity mouse model. In this connection, we conducted a study to find out whether propolis could reduce dyslipidemia induced by the intake of high-fat diet and thus protect the liver function and cardiovascular system. The effect of propolis on cardiovascular function was assessed by measuring lipid profile parameters that we used to calculate atherogenic indices, namely the atherogenic index of plasma (AIP), the cardiac risk ratio (CRR), and the atherogenic coefficient (AC). Due to the numerous antioxidant mechanisms of flavonoids present in propolis, including the free radical scavenging capacity, the inhibition of prooxidative enzymes, the metal chelating activity, and the synthesis of antioxidative enzymes (useful in protecting the body from hypertension, atherosclerosis, and cardiovascular diseases), we believe that it is worth investigating the effect of propolis on dyslipidemia induced by the intake of a high-fat diet. The goal is to try to prevent the consequences of dyslipidemia, e.g., cardiovascular diseases and hepatorenal disturbances.
3. Experimental Design
For the treatment group, edible sunflower seed oil (Zvijezda, Zagreb, Croatia) was used as the high-fat diet (HFD). A ratio of unsaturated/saturated = 7.3, 7% palmitic acid, 5% stearic acid, 19% oleic acid, 68% linoleic and 1% alpha linolenic acid was used. Through the whole experiment, on the days of treatment, the body weights of the animals were recorded, and food intake was monitored every two days. Each group contained nine animals. The first group was the control group treated with 0.2 mL/d of water with 1% of ethanol (vehicle). The second group was given the ethanol extract of propolis (EEP) (0.2 mL/d) at a dose of 50 mg/kg [
3]. The third group was treated with HFD (sunflowers seed oil, 0.1 mL/d) together with 0.1 mL/d of vehicle. The fourth group received the same amount (0.1 mL/d) of sunflower seed oil together with the ethanol extract of propolis (0.1 mL/d) at a dose of 50 mg/kg (HFD + EEP). All treated groups received (by gauge) a total volume of 0.2 mL/d repeatedly for 30 days. To avoid the false changes influenced by excessive blood loss caused by the frequent sampling, the whole blood samples and liver were collected on the 30th day of the experiment, respectively. On the 30th day, the animals were anesthetized (isoflurane, 2% in a flow of oxygen) and exsanguinated by intra-cardiac puncture without anticoagulant, as described in European Mouse Phenotyping Resource for Standardized Screens (EMPReSS), SOP (standard operating procedure) [
29] under veterinary supervision. After coagulation, the un-hemolyzed serum was collected and frozen at −80 ℃ until further processing. The liver was isolated from the animals instantly, and all further processing was done on ice. The liver weight was recorded. Livers were isolated and frozen at −80 ℃ until further processing. A sample of the whole liver (15–25 mg) was taken and processed for further triglyceride and total cholesterol assay protocols.
The whole experiment was repeated twice, and the statistical analysis showed no difference between the first and the second experimental setup (repetition).
3.1. Body Weight
During the study period of 30 days, the mice were weighed every day using the electronic balance, and their body weights were recorded. The body weight was expressed as a difference between the body weight on a certain day minus the weight on the initial day.
3.2. Estimation of the Serum Liver Enzyme Activity and Kidney Function
Serum biochemical parameters for renal and hepatic function included aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), urea, creatinine, blood glucose levels (glucose), lactate dehydrogenase (LDH), and total protein. After coagulation, the un-hemolyzed serum was collected and frozen at −80 ℃ until the further processing of biochemical parameters with the apparatus Alcylon 300 (Abbott, Lake Forest, IL, USA). The percentage of glycemic changes was calculated according to the formula:
where G
0 = blood glucose level at day zero, and G
x = blood glucose level at day 30.
3.3. Estimation of Lipid Profile Parameters in Serum and Liver
The experiment and the biochemistry analysis were conducted according to the recommendations of the International Federation of Clinical Chemistry (IFCC) methods in enzymology and were done with commercial kits (Merck, Darmstadt, Germany) on the Hitachi 717 automatic analyzer (Hitachi, Tokyo, Japan). All analyzed parameters were measured from the un-hemolyzed blood serum and liver homogenate at room temperature. Briefly, the activity of LDH (E.C.1.1.1.27) was measured under 340 nm by pyruvate to lactate continuous turnover measurement reaction [
30]. ALP (E.C.3.1.3.1.) was measured at 405 nm, substrate 4-nitrophenilphosphate [
31]. AST (E.C. 2.6.1.1.) and ALT (E.C.2.6.1.2.) were measured at 340 nm [
32]. Creatinine concentrations were measured with the method of alkaline picrate complex at 492 nm [
33]. Total protein concentration was measured by the method of Lowry [
34]. Triglycerides were measured by the colorimetric enzymatic method, which utilized the lipoprotein lipase for rapid and complete hydrolysis of triglycerides into glycerol followed by oxidation to dihydroxyacetone phosphate and hydrogen peroxide. The hydrogen peroxide reacted with 4-aminophenone and 4-chlorophenol under peroxidase catalytic action to form a red dye. The concentration of triglycerides was proportional to the intensity of the color generated and measured photometrically.
Total cholesterol was evaluated by enzymatic colorimetric method. Cholesterol esters were cleaved through the action of cholesterol esterase, producing free cholesterol and fatty acids. Cholesterol oxidase catalyzed the oxidation of cholesterol to cholest-4-en-3-one and hydrogen peroxide. In the presence of oxidase, the formed hydrogen peroxide affected the oxidative coupling of phenol and 4-aminoantipyrine, forming a quinone-imine red dye. The color intensity was directly proportional to cholesterol concentration, and the absorbance reading was at 512 nm.
High density lipoprotein cholesterol (HDL-c) was analyzed by the homogeneous colorimetric enzymatic method. In the presence of magnesium ions, dextran sulfate selectively formed water soluble compounds with low density lipoprotein cholesterols (LDL-c), very-low-density lipoprotein cholesterol (VLDL-c), and chylomicrons, which are resistant to polyethylene glycol-modified enzymes. Under the influence of the cholesterol enzyme, the cholesterol esters were quantitatively decomposed into free cholesterol and fatty acids. In the presence of peroxidation, the hydrogen peroxide generated reacted with 4-aminoantipyrine, forming a purple-bluish dye that was directly proportional to cholesterol concentration and was measured photometrically. All these parameters were measured using the Hitachi 717 automatic analyzer (Hitachi, Tokyo, Japan). The LDL-c concentrations were calculated from the Friedewald equation according to the manufacturer’s instructions [
35]:
3.4. Atherogenic Indices
After determining the concentration in mmol/L of total cholesterol (TC), total triglyceride (TG), HDL-c, and LDL-c fractions, atherogenic indices [AIP, CRR, AC, and cardioprotective indeks (CPI)] were calculated by using the values of lipid profile parameters in the following way:
3.5. Statistical Analysis
Statistical analyses were performed using Statistica 13.0 software (Stat-Soft, Tulsa, USA). Enzyme activity was expressed as the mean value of the group (±SD of the mean). Multiple comparisons between the control and the treated groups were done by ANOVA and Tukey post hoc tests. The level of statistical significance was set at
p ≤ 0.05 [
36].
5. Discussion
Obesity is associated with metabolic disorders such as dyslipidemia, atherosclerosis, and type 2 diabetes and is the result of energy imbalance—an increased caloric intake ratio compared to calorie loss. The study of natural preparations with hypoglycemic, hypotensive, and hypolipidemic potential is an interesting approach in ethno-pharmacological research. The present study was designed to identify the antioxidative capacity of propolis and its long-term hypoglycemic and hypolipidemic effects as well as its weight improving pattern in mice fed with the HFD.
According to a paper by Martins and Redgrave [
37], a diet-induced obesity animal model was developed to investigate human obesity. This model imitates the effect of human obesity better than the genetic obesity model and is based on a diet with a high fat content leading to obesity, hyperglycemia, and hyperlipidemia. In this work and other works, the elevated levels of TC, TG, and LDL-c in serum are factors of risk for the development of atherosclerosis and other cardiovascular diseases (CVD). In contrast, the elevated HDL-c level is associated with reduced risk for atherosclerosis.
Our results show that mice fed with the HFD and EEP over 30 days had higher levels of HDL-c compared with mice fed only with the HFD and consequently had lower levels of serum triglycerides and LDL-c, suggesting that EEP can effectively regulate metabolism triglycerides and cholesterol in HFD fed mice. Thus, propolis may be beneficial for treating patients with hypercholesterolemia and hypertriglyceridemia.
In this study, EEP increased plasma HDL cholesterol levels (
Table 2) in both normolipidemic and hyperlipidemic mice. This effect may be mediated by the presented polyphenols/flavonoids in propolis [
38]. However, coronary heart disease is accompanied and connected to high plasma LDL cholesterol [
39]. Oxidation of LDL into oxidated low-density lipoprotein (ox-LDL) indicates the first step of atherosclerosis in cardiovascular diseases, stimulating the immune and the inflammatory reactions that initiate the process of atherosclerotic plaque buildup. Oxidized LDL damages vascular endothelial walls, induces the production of local hormones from blood vessel walls, promotes inflammation, and attracts macrophages that uptake ox-LDL via scavenger receptors and transform macrophages into foam cells. High HDL reverses cholesterol transport by scavenging excess cholesterol from peripheral tissues to the liver for its metabolism and excretion [
40] and by inhibiting the oxidation of LDL. In addition, HDL can reduce or neutralize the atherogenic effects of oxidized LDL in artery walls. This effect may be increased by EEP and its antioxidative capacity (
Figure 3). EEP increases HDL-c levels while reducing LDL-c and has the properties to reduce oxidation of LDL by two different antioxidant mechanisms—the activation of the transcription factor NrF2 and the enhancement of the antioxidant enzymes such as heme oxygenase-1, phase II detoxification enzymes, and enzymes involved in glutathione (GSH) metabolism [
40,
41,
42,
43]. The secondary mechanism could refer to the neutralization of oxidative species, and it inhibits the activation of the nuclear transcription factor-
κB (NF-
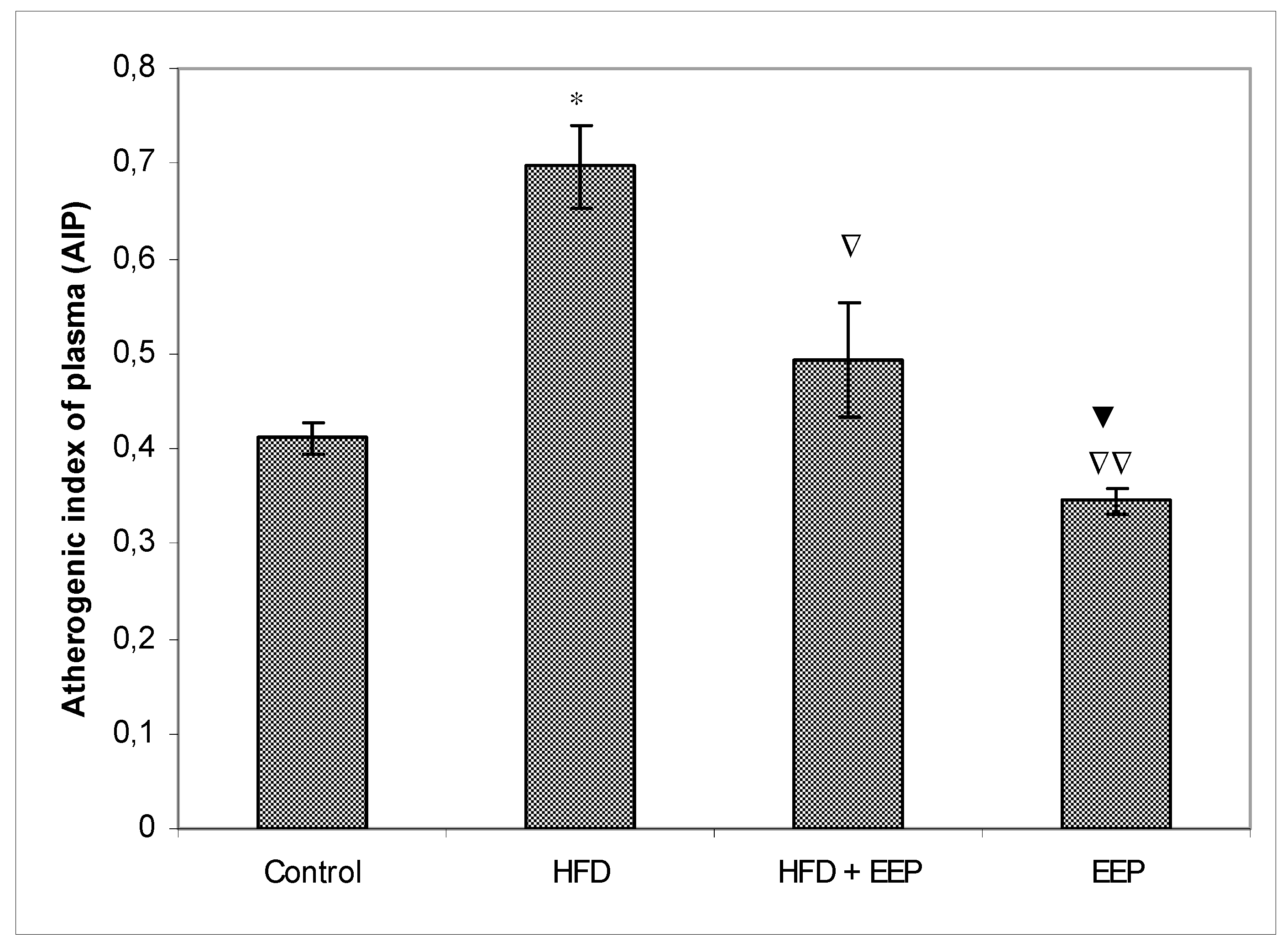
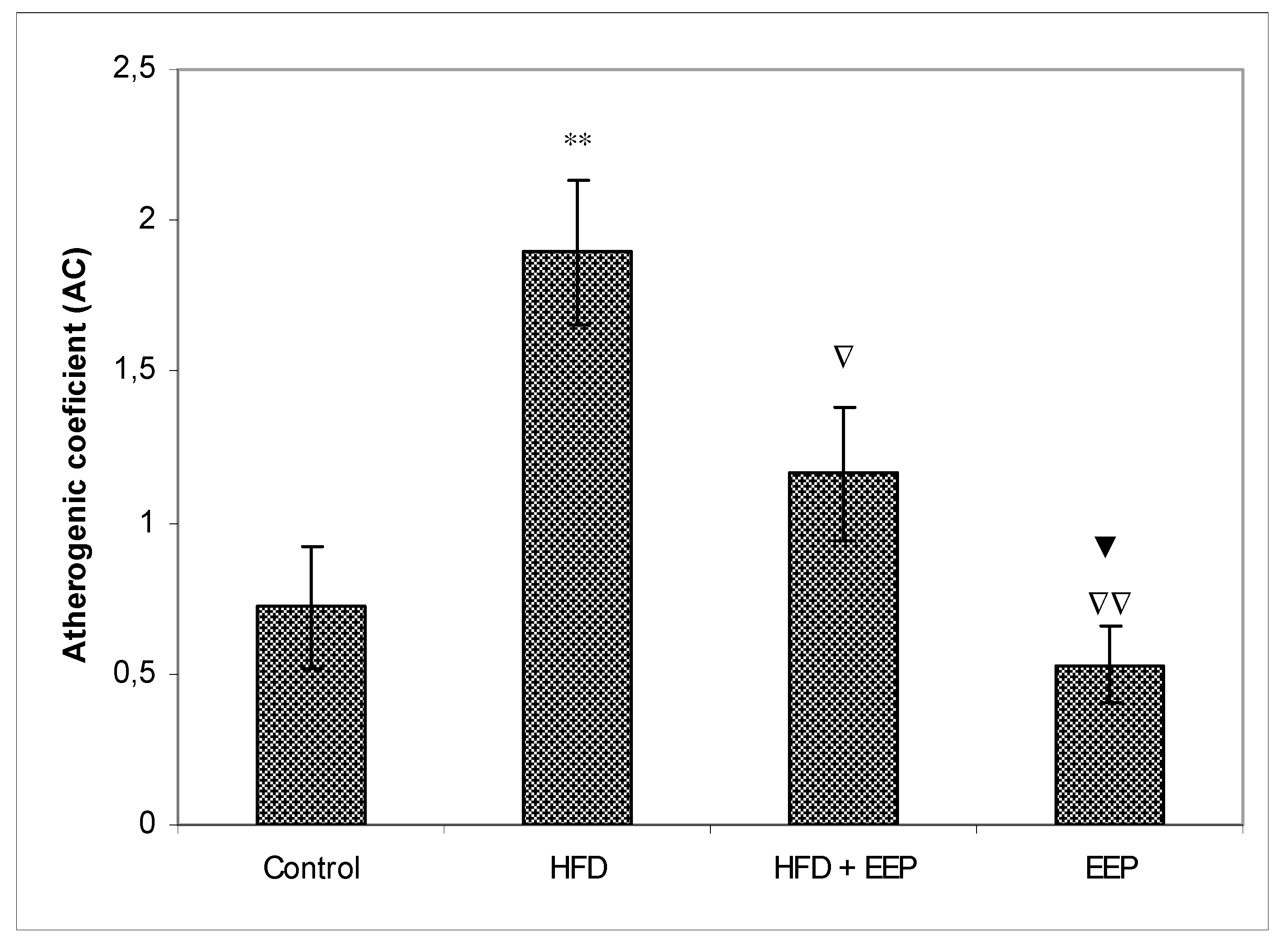
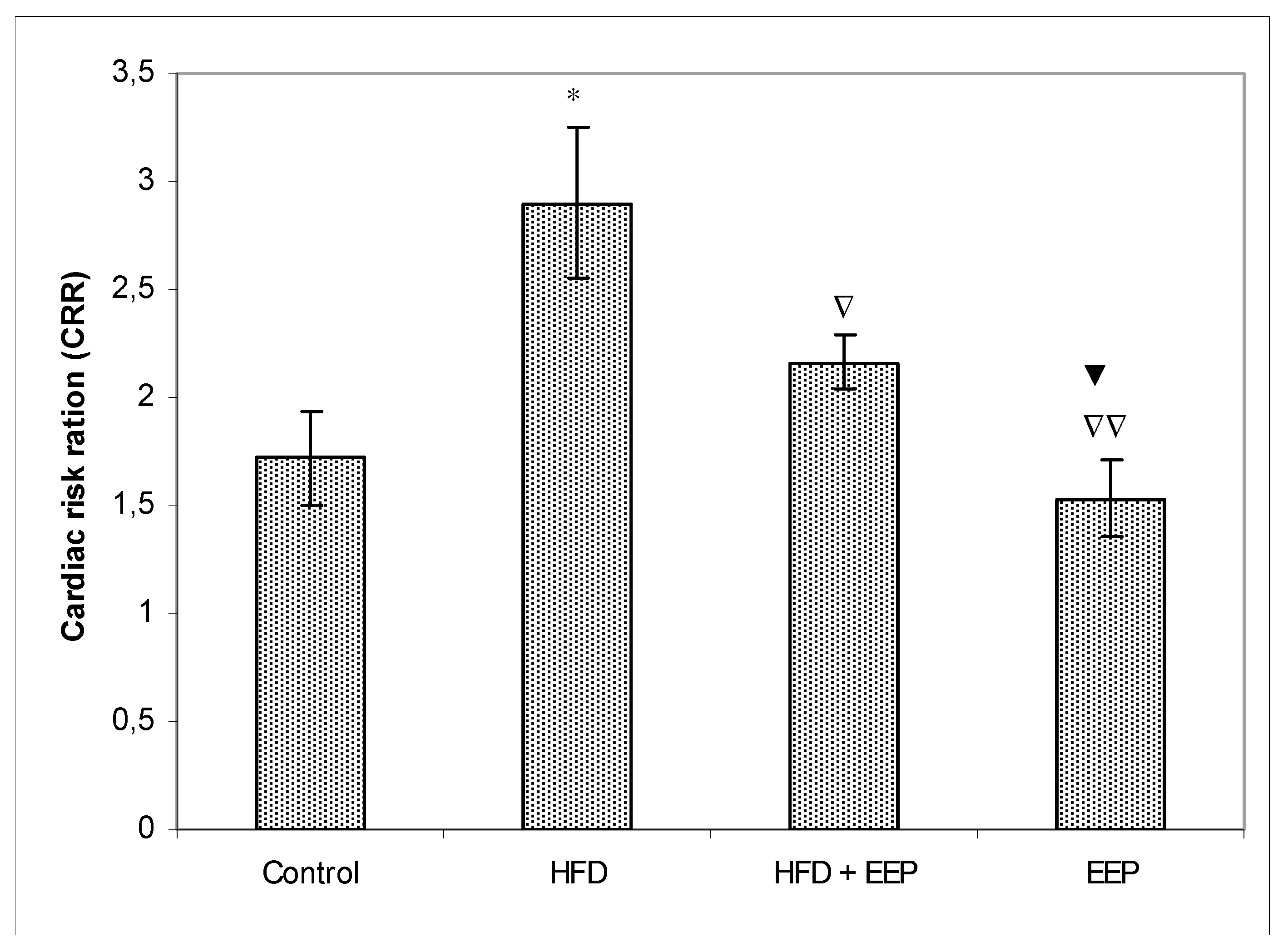
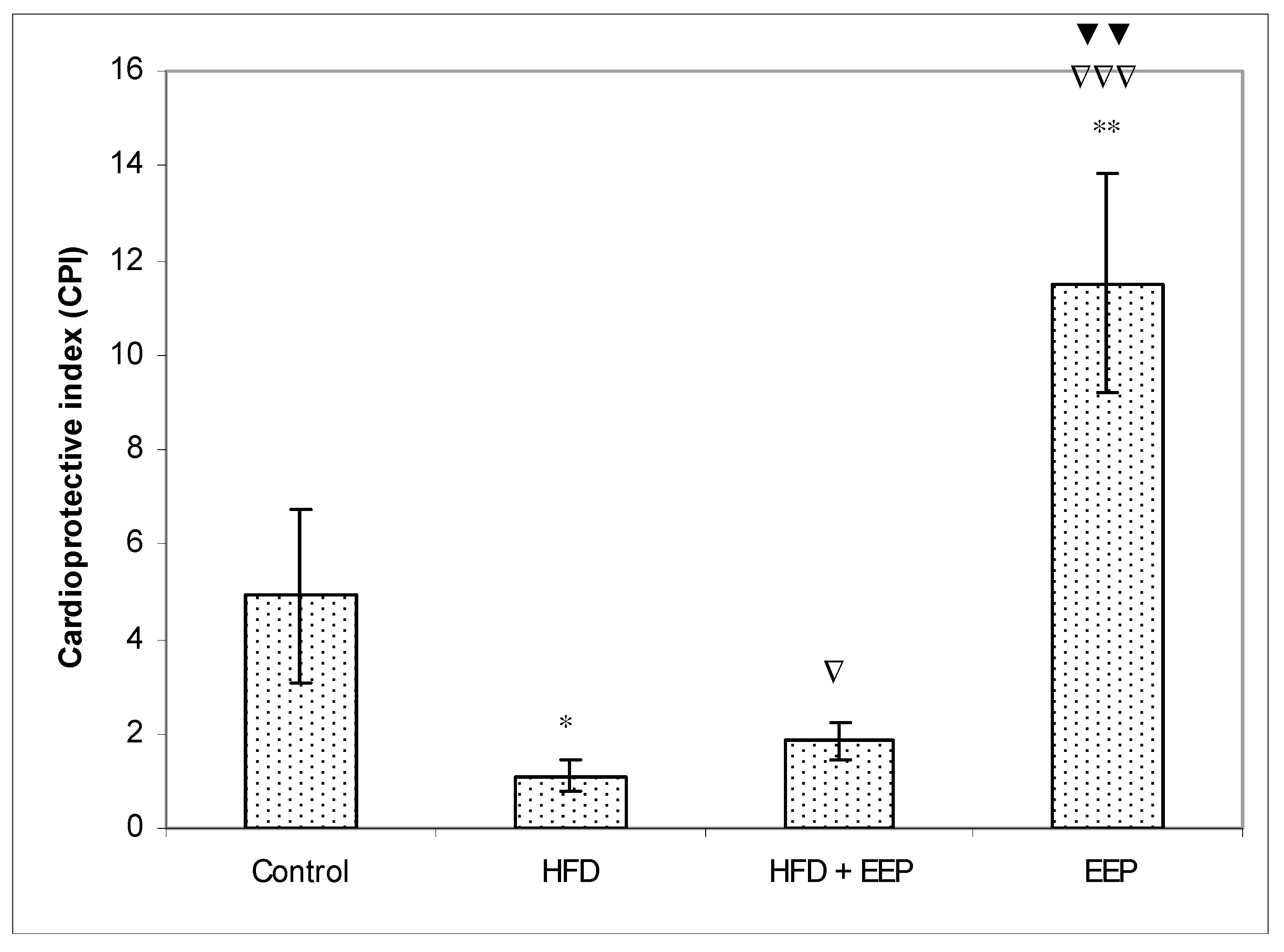
κB) signaling pathways. Reduced levels of LDL-c in HFD mice treated with EEP are the beneficial aspect of this current research in addition to showing the possible anti-atherosclerotic effects of EEP. This observation was consistent with the calculated indices such as AIP, AC, CRR, and CPI, which were all (exept CPI) attenuated following intervention with the EEP treatment (
Figure 5,
Figure 6,
Figure 7 and
Figure 8). These indices are valuable in assessing the risk of developing cardiovascular diseases; the more accurate the increase in AIP, AC, and CRR is, the more there is a predisposition for cardiovascular disease. CPI is highly improved in EEP treated normolipidemic and hyperlipidemic mice.
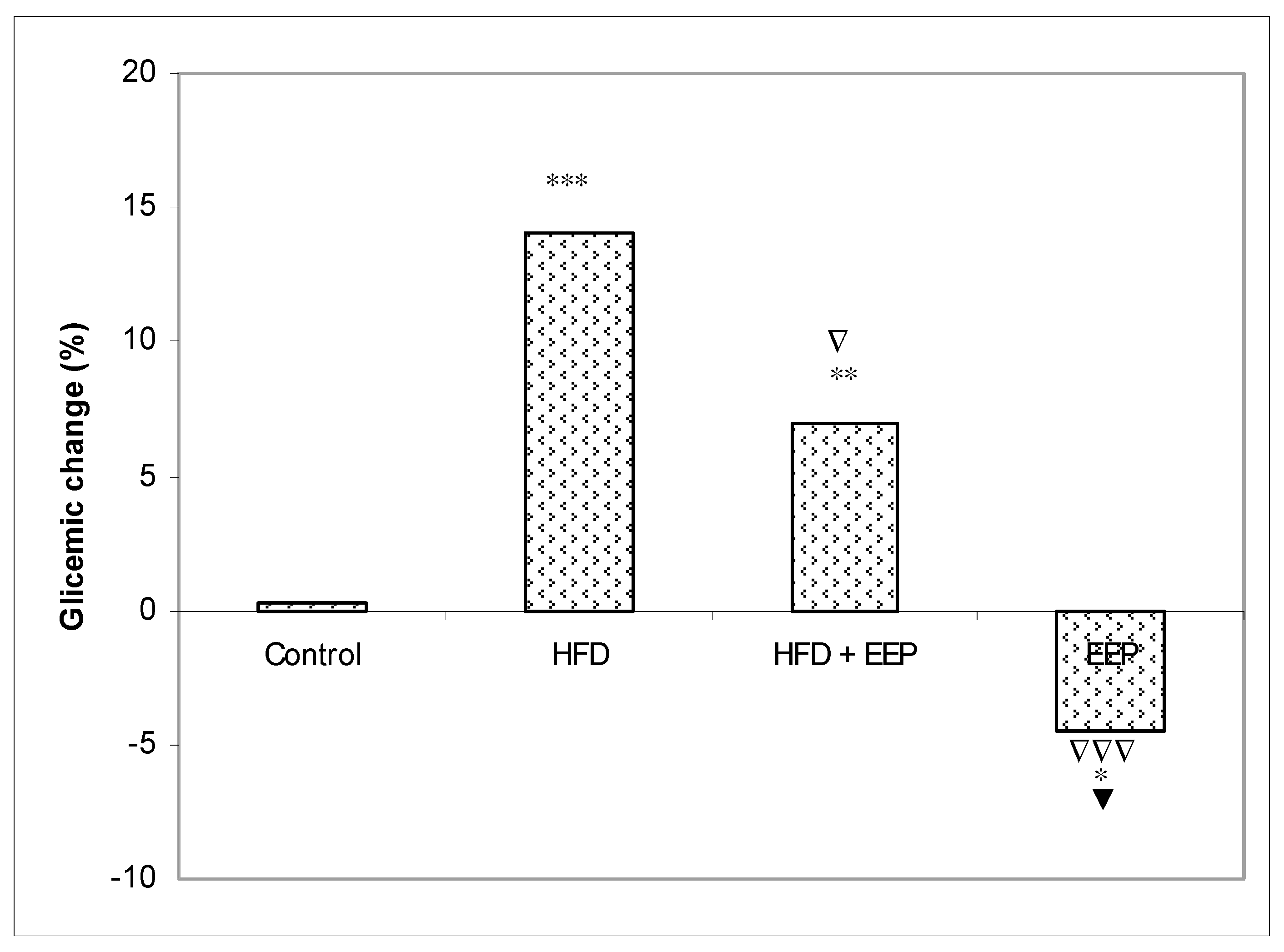
According to our results based on the antioxidative capacity of EEP, it seems that EEP with its compounds is a strong inhibitor of LDL oxidation. Oxidized LDL lipoproteins can be diminished with EEP by its antioxidant capacity, which can inactivate ROS and consequently counteract plasma LDL oxidation and reduce inflammation of the blood vessel endothelium. This antioxidative mechanism could contribute to the protective effect against cardiovascular diseases or other chronic diseases connected with oxidative stress. It seems that the consumption of the propolis diet was linked with hypolipidaemic effects specifically associated with decreased levels of LDL-c and TG and increased levels of HDL-c. EEP could exert health benefits not only by scavenging free radicals but also by hypoglycemic and regenerative effects. Blood glucose reduction may relate to the bioactive compounds of EEP and its protective effect on β-cells, which could enhance the production of insulin or enhance cellular sensitivity responses to insulin. The positive effect of EEP is also apparent in its hypoglycemic action (
Figure 9). The hypoglycemic effect of EEP (
Figure 9) may be due to: (i) an increase in glucose utilization by peripheral tissues; (ii) inhibiting the activity of enzymes included in cholesterol biosynthesis; (iii) decreased absorption of glucose in the intestine via the inhibition of the enzymatic activity of α-glucosidase; (iv) increased activity of the hepatic enzyme glucokinase, the markedly reduced enzymatic activities of hepatic glucose-6-phosphatase and phosphoenolpyruvate carboxykinase (PEPCK); or (v) the improvement in lipolysis by reducing the activity of hormone-sensitive lipase [
2,
44].
In addition, EEP improved the lipid profile of animals fed with the HFD diet and lowered their atherogenic indices closer to the levels of the control animals. A decrease in atherogenic indices is a positive physiological effect. The positive effects of propolis are induced by its bioactive components, especially the most explored polyphenol/flavonoid components such as epigallocatechin gallate, curcumin, reservatrol, quercetin, chrysin, naringenin, caffeic acid, caffeic acid phenethyl ester, proanthocyanidins, gallic acid, rosemary acid, isorhamnetin, camphorol, lutein, and pinocebrin [
2,
3,
6,
38,
42,
45,
46,
47].
Our results regarding HFD supplementation to the standard in an ad libitum diet are consistent with the data of other authors who used the same mice model [
48,
49,
50]. In addition to the potential to prevent hyperlipidemia and influence lipid metabolism, EEP also affected the overall body weight of the animals (
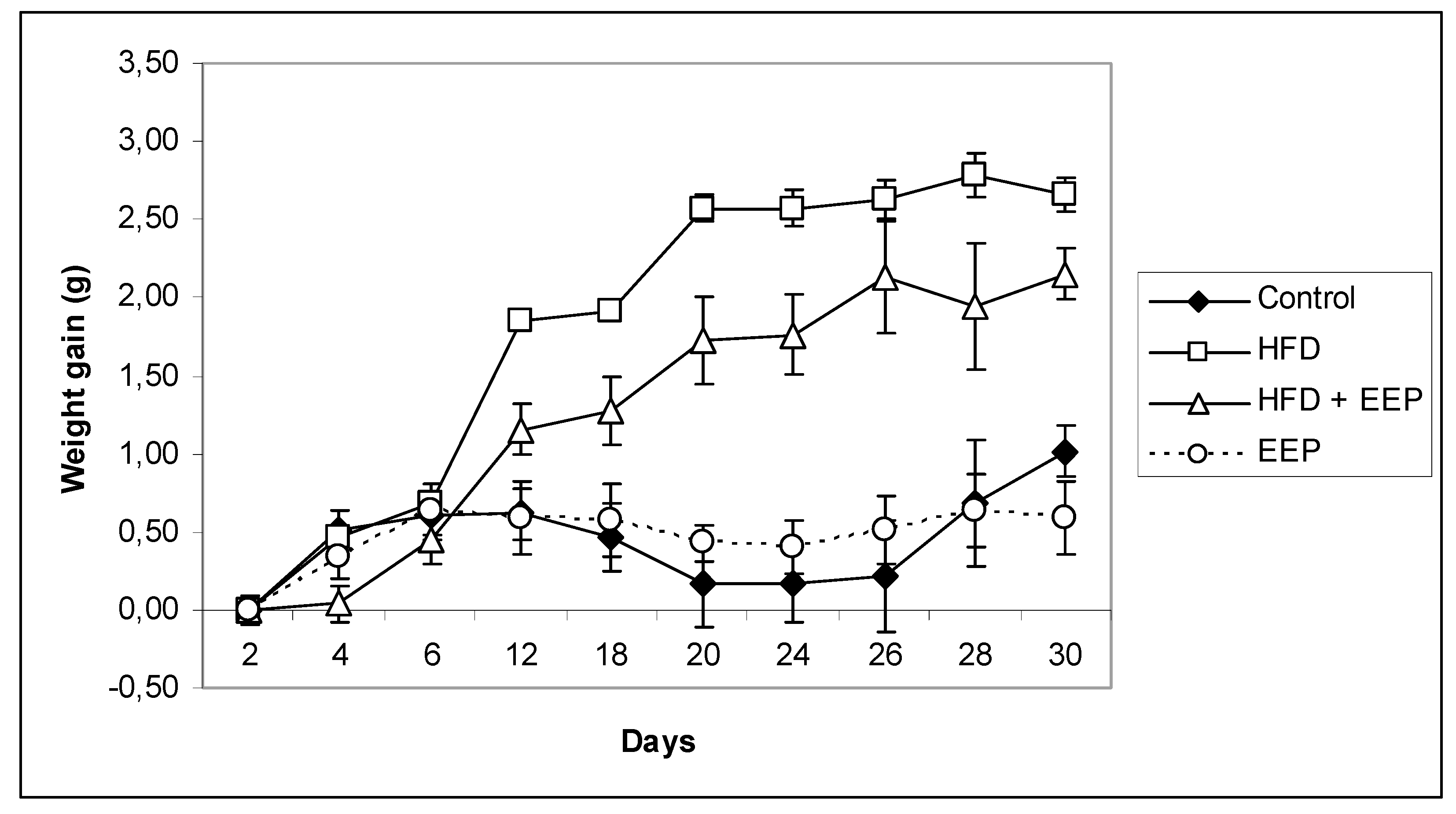
Figure 4). The results suggest that EEP is capable of reducing weight gain with no different food intake (
Table 3). This data confirms the role of EEP in the regulation of carbohydrate and lipid metabolism. Thus, the physiological mechanism by which EEP prevents obesity in mice is activated by at least ten consecutive doses of EEP treatment. Confirmation of that physiological observation was found in the EEP treated group where, on the 10th day, the weight gain decreased in comparison to all other groups and remained rather uniformly unchanged until the end of the experimental period. To normalize weight gain change to food intake, we estimated the FER, which was calculated by dividing the weight increase by the food intake. In the HFD group, the FER was 3.1× greater in relation to the control group, which is a typical symptom of obesity, while in the HFD + EEP group, the FER was 2.3×. Food intake of these treatment groups was lower because dietary fat is a strong suppressor of food intake due to the fat induced secretion of satiety peptides. Our findings are similar to those in Hong et al. and Shin et al. [
51,
52]. These data collectively suggest that the supplementation of EEP attenuates HFD induced body weight gain, which is attributable to fat mass reduction possibly by inhibiting adipogenesis and increasing energy expenditure [
51,
52]. Further experiments will show what biochemical changes in lipid physiology occur during that period and whether they attribute to the overall body weight and lipid metabolism (
Table 4). There are only a few works dealing with high EEP consumption and long-term consumption of this honeybee product [
53].
Liver enzyme profiles, which are sometimes used as screening marker of hepatotoxicity, measured in the EEP treated group in this experiment remained similar to the control group, indicating that no unfavorable effects were noticed in the experimental animals. EEP showed a hepatoprotective role. It reduced the lipid content in the liver (
Table 4) and improved the liver function, as indicated by liver enzymes such as aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), and lactate dehydrogenase (LDH) in mice fed with the HFD and in healthy mice (
Table 6). These enzymes are liver-specific enzymes that provide a deep insight about hepatic functioning; they elevate in cases of hepatic injuries or malfunctioning.
Renal function was also improved after EEP treatment in normolipidemic and hyperlipidemic mice (
Table 5). Therefore, Oršolić et al. [
3] demonstrated that the antioxidant and the anti-inflammatory effects of EEP were able to attenuate hypertension and structural hepatorenal damage in alloxan-induced diabetes mice models. Similar results were found in Koya-Miyata and co-workers that used Brazilian propolis, but there are differences between Croatian and Brazilian propolis [
50,
53]. The plant sources represent different ecosystems that have a great impact on the physical structure and the most important chemical composition of propolis, thus the chemical composition of propolis depends on the site of collection, th egeographic zone, the local flora, and the bee species [
17,
18,
19,
20]. Flavonoid-aglycones (flavones, flavonols, flavanones) and phenolic acids and their esters are the main biologically active components in propolis from the Northern Hemisphere, whereas in Brazilian propolis, the main biologically active components are prenylated phenylpropanoids (prenylated cinnamic acids and their esters), cinnamic acid derivatives, chromane, naphthalene, and anthracene derivatives, triterpens and steroids, and di- and sesquiterpenes. Also, in Europe, Northern America, and non-tropical regions in Asia, the main plant sources of propolis are cracks in the bark of trees and leaf buds from poplars (
Populus spp., mainly from
P. nigra L.), followed by birches (
Betula spp.), elms (
Ulmus spp.), pine trees (
Pinus spp.), oaks (
Quercus spp.), willows (
Salix spp.), horse chestnut trees (
Aesculus hyppocastanum L.), spruce (
Picea spp.), and the ash (
Fraxinus spp.). The Brazilian sources are
Baccharis spp. (mainly
B. dracunculifolia), but bees collect resins from other plant species as well, such as
Araucaria spp.,
Clusia spp.,
Eucalyptus spp.,
Vernonia spp.,
Diclenia spp.,
Hyptis spp.,
Myrcia spp.,
Schinus spp., and
Weinmania spp. Bankova [
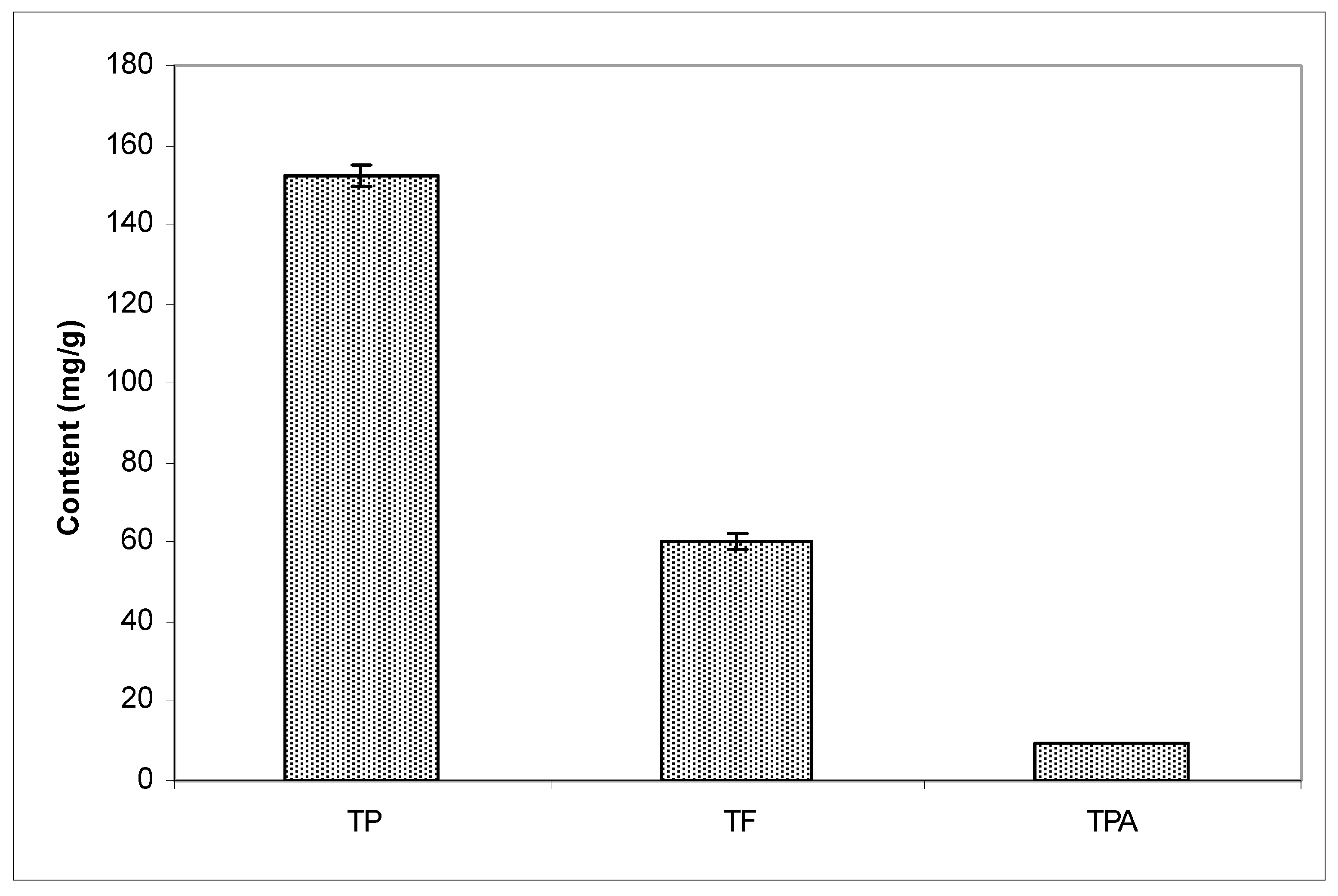
54] determined by plant origin recognizes several chemotypes, but if we compare Croatian and Brazilian propolis, then we distinguish poplar- and alecrim-chemotypes of propolis. As already mentioned, differences in the biological activity of propolis have been found in samples gathered from different races of honeybees. Therefore, it is very important to define the biological activity of propolis. Chemical analysis shows that Croatian propolis used in this study had a higher content of bioactive components than the ones described in references [
3,
4,
5,
6,
7,
8,
9,
10,
11,
16,
17,
18,
19,
20]. European propolis is known to have a high flavonoid content, often surpassing 20% in comparison to Brazilian propolis [
55]. According to the results of Kosalec et al., the Croatian sample of propolis was richer in extractable substances than the Brazilian sample (27.53% and 13.15%, respectively) using ethanol. Additionally, Croatian ethanolic propolis extracts demonstrated more potent antibacterial, antitumor, antimetastatic, and immunomodulatory activity than Brazilian ethanolic propolis extracts [
14,
15,
20]. It seems that bergamot, an endemic plant of the Calabria region (Italy), has a similar content of flavonoids as Croatian propolis and has an effect on weight gain, lipid clearance, and blood triglycerides [
56].
Summarizing all the results, we can conclude that oral administration of EEP suppresses overall weight gain, reduces lipid profiles in serum and liver, increases hepatorenal function, and reduces atherogenic indices in mice fed with a HFD.

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}