BG-4 from Bitter Gourd (Momordica charantia) Differentially Affects Inflammation In Vitro and In Vivo


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of BG-4
2.2. In Vitro Antioxidant Activity of BG-4
2.3. Measurement of Pro-Inflammatory Markers In Vitro
2.4. Dosage Information and In Vivo Experimental Procedure
2.5. Myeloperoxidase Assay in the Colon
2.6. Measurement of Hemoglobin Content in the Feces
2.7. Measurement of Cytokines by ELISA
2.8. Statistical Analysis
3. Results and Discussion
3.1. BG-4 Exert Antioxidant and Antiradical Activity
3.2. BG-4 Decreased Expression of Pro-Inflammatory Markers In Vitro
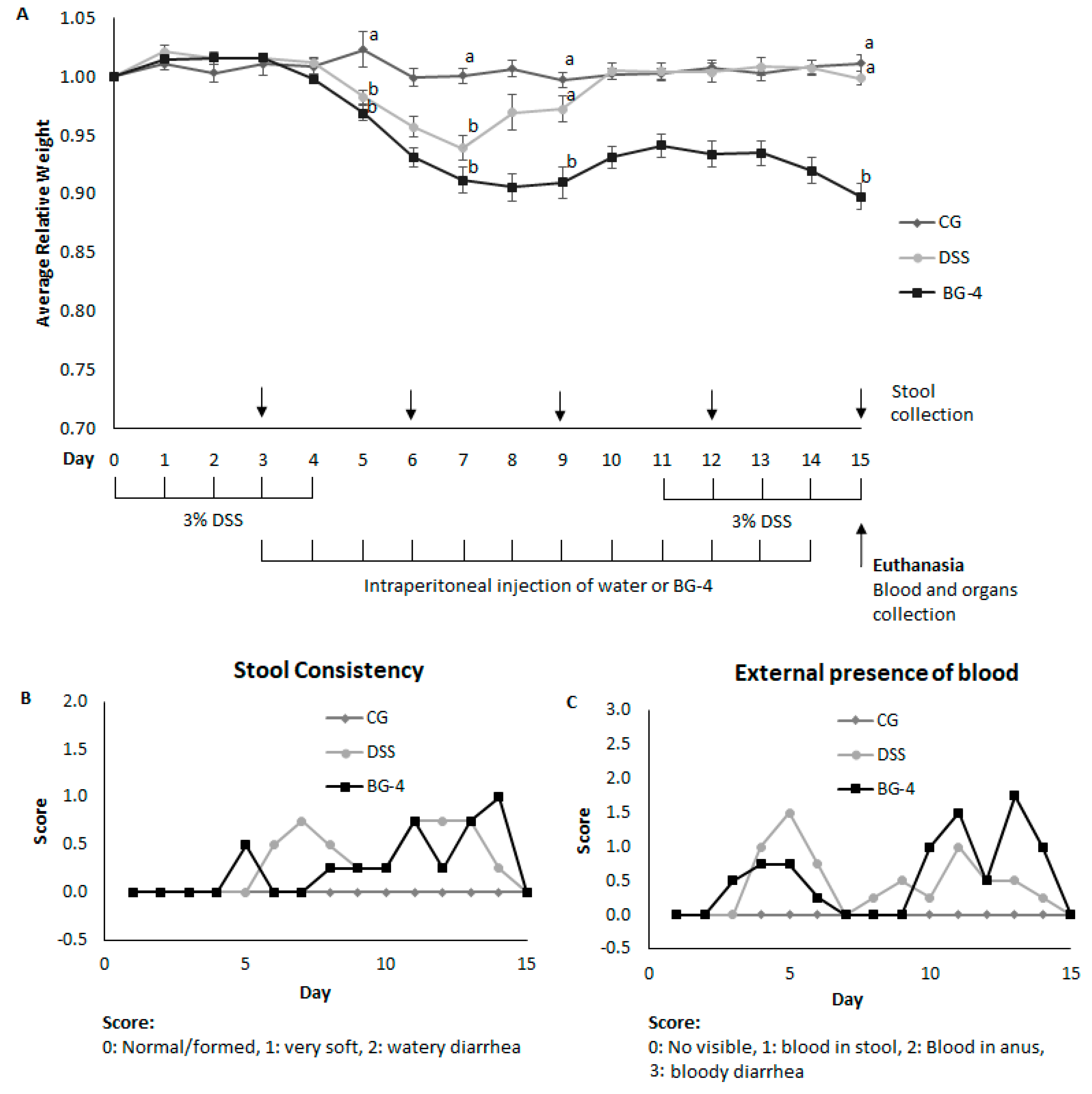
3.3. IP Administration of BG-4 did not Alleviate Indicators of Colitis in Vivo
3.4. IP Administration of BG-4 Aggravated DSS-Induced Colitis in Mice
3.5. IP Administration of BG-4 Exerted a Differential Systemic and Localized Effect in Pro-Inflammatory Cytokines In Vivo
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Egberg, M.D.; Kappelman, M.D.; Gulati, A.S. Improving Care in Pediatric Inflammatory Bowel Disease. Gastroenterol. Clin. North Am. 2018, 47, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Crowley, E.; Muise, A. Inflammatory Bowel Disease: What Very Early Onset Disease Teaches Us. Gastroenterol. Clin. North Am. 2018, 47, 755–772. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Chang, L.; Chang, H.M.; Chang, F. Intestinal and Extraintestinal Cancers Associated With Inflammatory Bowel Disease. Clin. Colorectal Cancer 2018, 17, e29–e37. [Google Scholar] [CrossRef]
- Sairenji, T.; Collins, K.L.; Evans, D.V. An Update on Inflammatory Bowel Disease. Prim. Care Clin. Off. Pract. 2017, 44, 673–692. [Google Scholar] [CrossRef] [PubMed]
- Palamthodi, S.; Lele, S.S. Nutraceutical applications of gourd family vegetables: Benincasa hispida, Lagenaria siceraria and Momordica charantia. Biomed. Prev. Nutr. 2014, 4, 15–21. [Google Scholar] [CrossRef]
- Lii, C.K.; Chen, H.W.; Yun, W.T.; Liu, K.L. Suppressive effects of wild bitter gourd (Momordica charantia Linn. var. abbreviata ser.) fruit extracts on inflammatory responses in RAW 264.7 macrophages. J. Ethnopharmacol. 2009, 122, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Dandawate, P.R.; Subramaniam, D.; Padhye, S.B.; Anant, S. Bitter melon: A panacea for inflammation and cancer. Chin. J. Nat. Med. 2016, 14, 81–100. [Google Scholar] [CrossRef]
- Xu, J.; Cao, K.; Feng, Z.; Liu, J. Benefits of the soluble and insoluble fractions of bitter gourd in mice fed a high-fat diet. J. Funct. Foods. 2018, 42, 216–223. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Khalid, S.; Tahir, F.; Sabitaliyevich, U.Y.; Yaylim, I.; Attar, R.; Xu, B. Bitter gourd (Momordica charantia) as a rich source of bioactive components to combat cancer naturally: Are we on the right track to fully unlock its potential as inhibitor of deregulated signaling pathways. Food Chem. Toxicol. 2018, 119, 98–105. [Google Scholar] [CrossRef]
- Raish, M.; Ahmad, A.; Ansari, M.A.; Alkharfy, K.M.; Aljenobi, F.I.; Jan, B.L.; Al-Mohizea, A.M.; Khan, A.; Ali, N. Momordica charantia polysaccharides ameliorate oxidative stress, inflammation, and apoptosis in ethanol-induced gastritis in mucosa through NF-kB signaling pathway inhibition. Int. J. Biol. Macromol. 2018, 111, 193–199. [Google Scholar] [CrossRef]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 110, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Raina, K.; Kumar, D.; Agarwal, R. Promise of bitter melon (Momordica charantia) bioactives in cancer prevention and therapy. Semin. Cancer Biol. 2016, 40–41, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Dia, V.P.; Krishnan, H.B. BG-4, a novel anticancer peptide from bitter gourd (Momordica charantia), promotes apoptosis in human colon cancer cells. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Jones, L.D.; Pangloli, P.; Krishnan, H.B.; Dia, V.P. BG-4, a novel bioactive peptide from Momordica charantia, inhibits lipopolysaccharide-induced inflammation in THP-1 human macrophages. Phytomedicine 2018, 42, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Vernaza, M.G.; Dia, V.P.; Gonzalez de Mejia, E.; Chang, Y.K. Antioxidant and Antiinflammatory Properties of Germinated and Hydrolysed Brazilian Soybean Flours. Food Chem. 2012, 134, 2217–2225. [Google Scholar] [CrossRef]
- Richard-Eaglin, A.; Smallheer, B.A. Immunosuppressive/Autoimmune Disorders. Nurs. Clin. North Am. 2018, 53, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Shajib, M.S.; Manocha, M.M.; Khan, W.I. Investigating Intestinal Inflammation in DSS-induced Model of IBD. J. Vis. Exp. 2012, 1–6. [Google Scholar] [CrossRef]
- Dai, Z.; Feng, S.; Liu, A.; Wang, H.; Zeng, X.; Yang, C.S. Anti-inflammatory effects of newly synthesized α-galacto-oligosaccharides on dextran sulfate sodium-induced colitis in C57BL/6J mice. Food Res. Int. 2018, 109, 350–357. [Google Scholar] [CrossRef]
- Welch, C.L.; Young, D.S. Spectrophotometry of occult blood in feces. Clin. Chem. 1983, 29, 2022–2025. [Google Scholar]
- Grajeda-Iglesias, C.; Salas, E.; Barouh, N.; Baréa, B.; Panya, A.; Figueroa-Espinoza, M.C. Antioxidant Activity of Protocatechuates Evaluated by DPPH, ORAC, and CAT Methods. Food Chem. 2016, 194, 749–757. [Google Scholar] [CrossRef]
- Foss, C.A.; Sanchez-Bautista, J.; Jain, S.K. Imaging Macrophage-associated Inflammation. Semin. Nucl. Med. 2018, 48, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Van den Bossche, J.; Saraber, D.L. Metabolic regulation of macrophages in tissues. Cell. Immunol. 2018, 330, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Fang, S.C.; Liu, C.W.; Chen, Y.F. Inhibitory effects of new varieties of bitter melon on lipopolysaccharide-stimulated inflammatory response in RAW 264.7 cells. J. Funct. Foods. 2013, 5, 1829–1837. [Google Scholar] [CrossRef]
- Dia, V.P.; Bringe, N.A.; de Mejia, E.G. Peptides in pepsin-pancreatin hydrolysates from commercially available soy products that inhibit lipopolysaccharide-induced inflammation in macrophages. Food Chem. 2014, 152, 423–431. [Google Scholar] [CrossRef] [PubMed]
- de Mejia, E.G.; Dia, V.P. Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides 2009, 30, 2388–2398. [Google Scholar] [CrossRef] [PubMed]
- An, J.Y.; Lee, H.H.; Shin, J.S.; Yoo, H.S.; Park, J.S.; Son, S.H.; Kim, S.W.; Yu, J.; Lee, J.; Lee, K.T.; et al. Identification and structure activity relationship of novel flavone derivatives that inhibit the production of nitric oxide and PGE2in LPS-induced RAW 264.7 cells. Bioorganic Med. Chem. Lett. 2017, 27, 2613–2616. [Google Scholar] [CrossRef] [PubMed]
- Sinha, C.K.; Coran, A.G. Ulcerative colitis. Handb. Pediatr. Surg. 2010, 201–206. [Google Scholar] [CrossRef]
- Wu, X.; Song, M.; Cai, X.; Neto, C.; Tata, A.; Han, Y.; Wang, Q.; Tang, Z.; Xiao, H. Chemopreventive Effects of Whole Cranberry (Vaccinium macrocarpon) on Colitis-Associated Colon Tumorigenesis. Mol. Nutr. Food Res. 2018, 1800942. [Google Scholar] [CrossRef]
- Stettner, N.; Rosen, C.; Bernshtein, B.; Gur-Cohen, S.; Frug, J.; Silberman, A.; Sarver, A.; Carmel-Neiderman, N.N.; Eilam, R.; et al. Induction of Nitric-Oxide Metabolism in Enterocytes Alleviates Colitis and Inflammation-Associated Colon Cancer. Cell Rep. 2018, 23, 1962–1976. [Google Scholar] [CrossRef]
- Huang, Y.; Guo, J.; Gui, S. Orally targeted galactosylated chitosan poly(lactic-co-glycolic acid) nanoparticles loaded with TNF-α siRNA provide a novel strategy for the experimental treatment of ulcerative colitis. Eur. J. Pharm. Sci. 2018, 125, 232–243. [Google Scholar] [CrossRef]
- Strzepa, A.; Pritchard, K.A.; Dittel, B.N. Myeloperoxidase: A new player in autoimmunity. Cell. Immunol. 2017, 317, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Hong Li, Y.; Adam, R.; Colombel, J.F.; Xiang Bian, Z. A characterization of pro-inflammatory cytokines in dextran sulfate sodium-induced chronic relapsing colitis mice model. Int. Immunopharmacol. 2018, 60, 194–201. [Google Scholar] [CrossRef]
- Wang, Y.; Ning, Z.H.; Tai, H.W.; Long, S.; Qin, W.C.; Su, L.M.; Zhao, Y.H. Relationship between lethal toxicity in oral administration and injection to mice: Effect of exposure routes. Regul. Toxicol. Pharmacol. 2015, 71, 205–212. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nieto-Veloza, A.; Wang, Z.; Zhong, Q.; Krishnan, H.B.; Dia, V.P. BG-4 from Bitter Gourd (Momordica charantia) Differentially Affects Inflammation In Vitro and In Vivo. Antioxidants 2019, 8, 175. https://doi.org/10.3390/antiox8060175
Nieto-Veloza A, Wang Z, Zhong Q, Krishnan HB, Dia VP. BG-4 from Bitter Gourd (Momordica charantia) Differentially Affects Inflammation In Vitro and In Vivo. Antioxidants. 2019; 8(6):175. https://doi.org/10.3390/antiox8060175
Chicago/Turabian StyleNieto-Veloza, Andrea, Zhihong Wang, Qixin Zhong, Hari B. Krishnan, and Vermont P. Dia. 2019. "BG-4 from Bitter Gourd (Momordica charantia) Differentially Affects Inflammation In Vitro and In Vivo" Antioxidants 8, no. 6: 175. https://doi.org/10.3390/antiox8060175
APA StyleNieto-Veloza, A., Wang, Z., Zhong, Q., Krishnan, H. B., & Dia, V. P. (2019). BG-4 from Bitter Gourd (Momordica charantia) Differentially Affects Inflammation In Vitro and In Vivo. Antioxidants, 8(6), 175. https://doi.org/10.3390/antiox8060175