A Red Fluorescent Protein-Based Probe for Detection of Intracellular Reactive Sulfane Sulfur


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Compounds
2.2. Protein Mutation, Expression, and Purification
2.3. Characterization of psRFP
2.4. Liquid Chromatography Tandem Mass Spectrum (LC-MS/MS) Analysis of psRFP
2.5. Analysis of Intracellualr Sulfane Sulfur with psRFP
3. Results
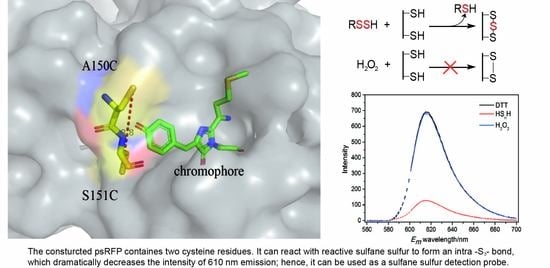
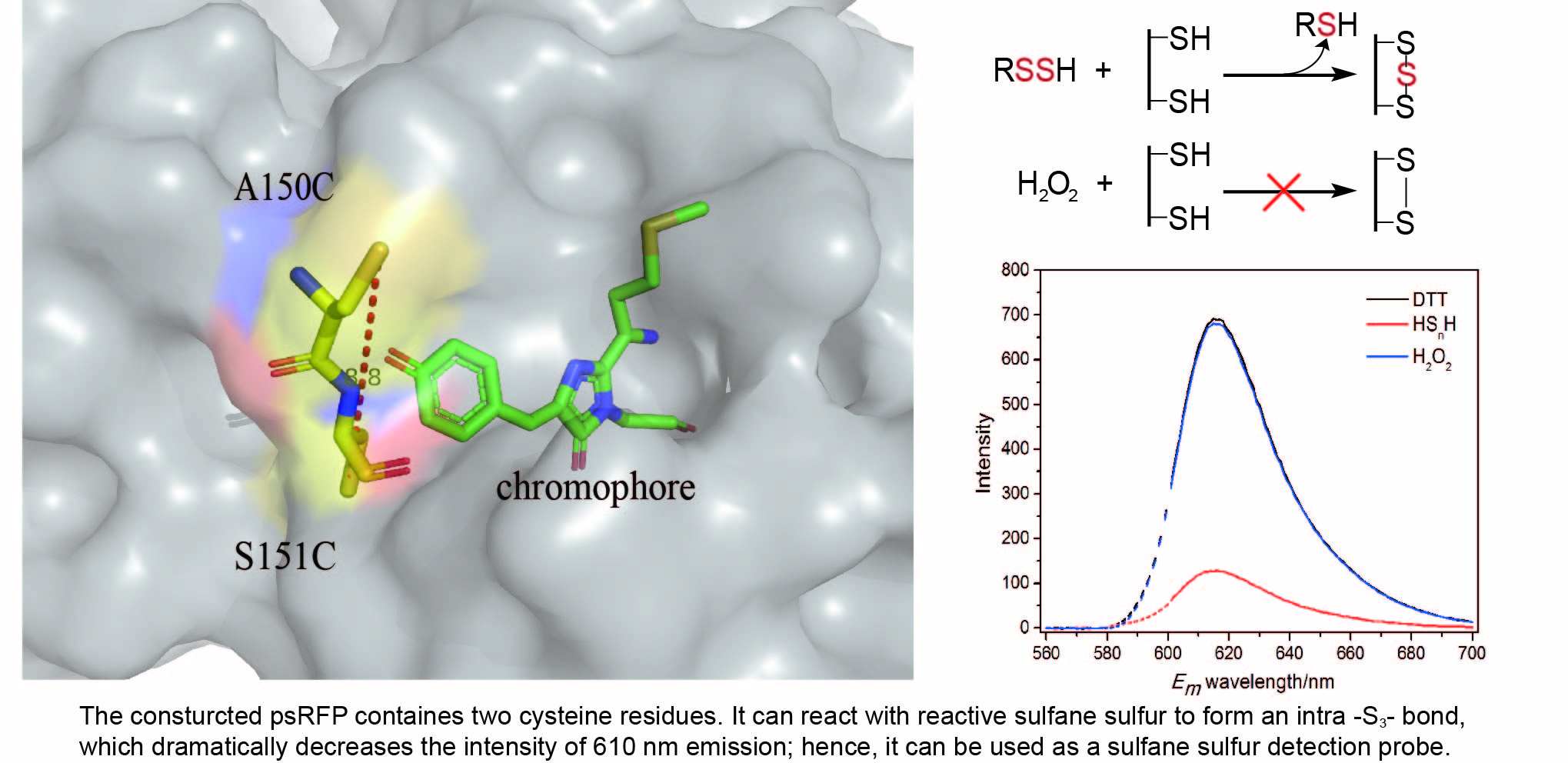
3.1. Design of the Reactive Sulfane Sulfur-Sensitive mCherry
3.2. Construction of the Hydrogen Polysulfide-Sensitive mCherry
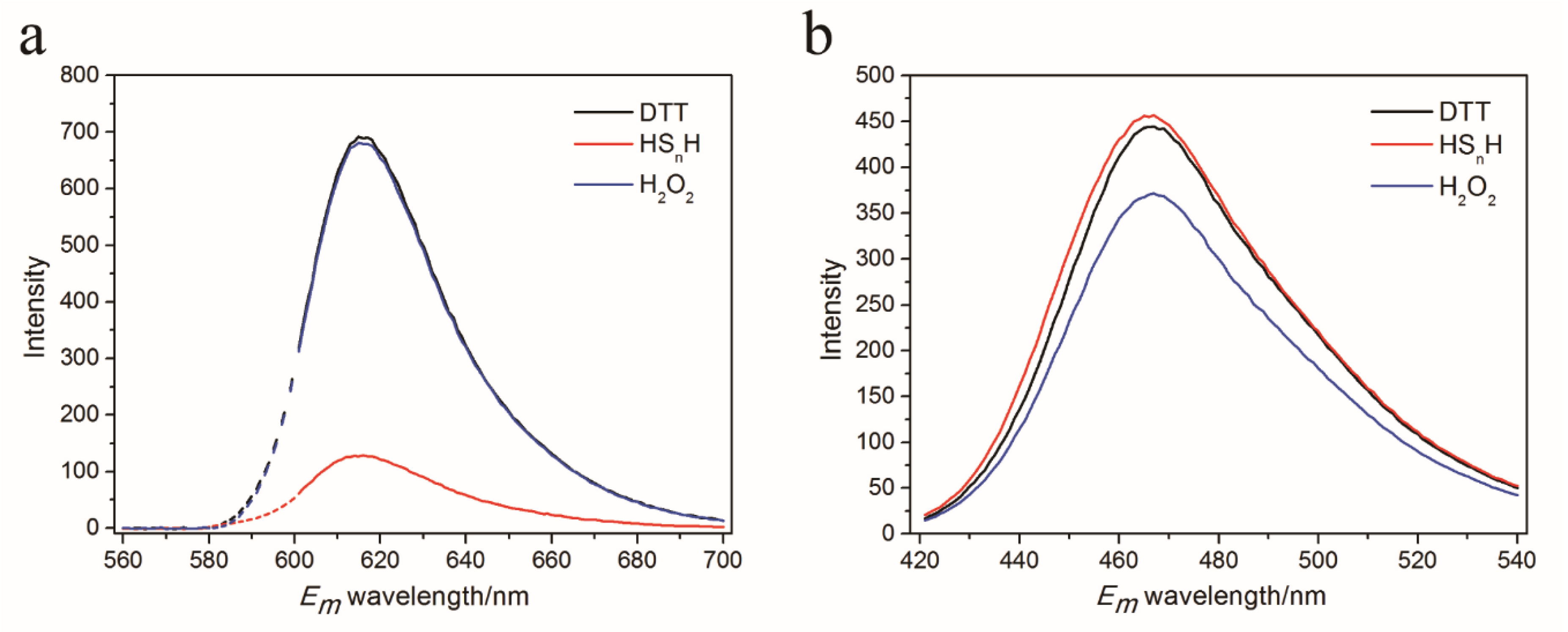
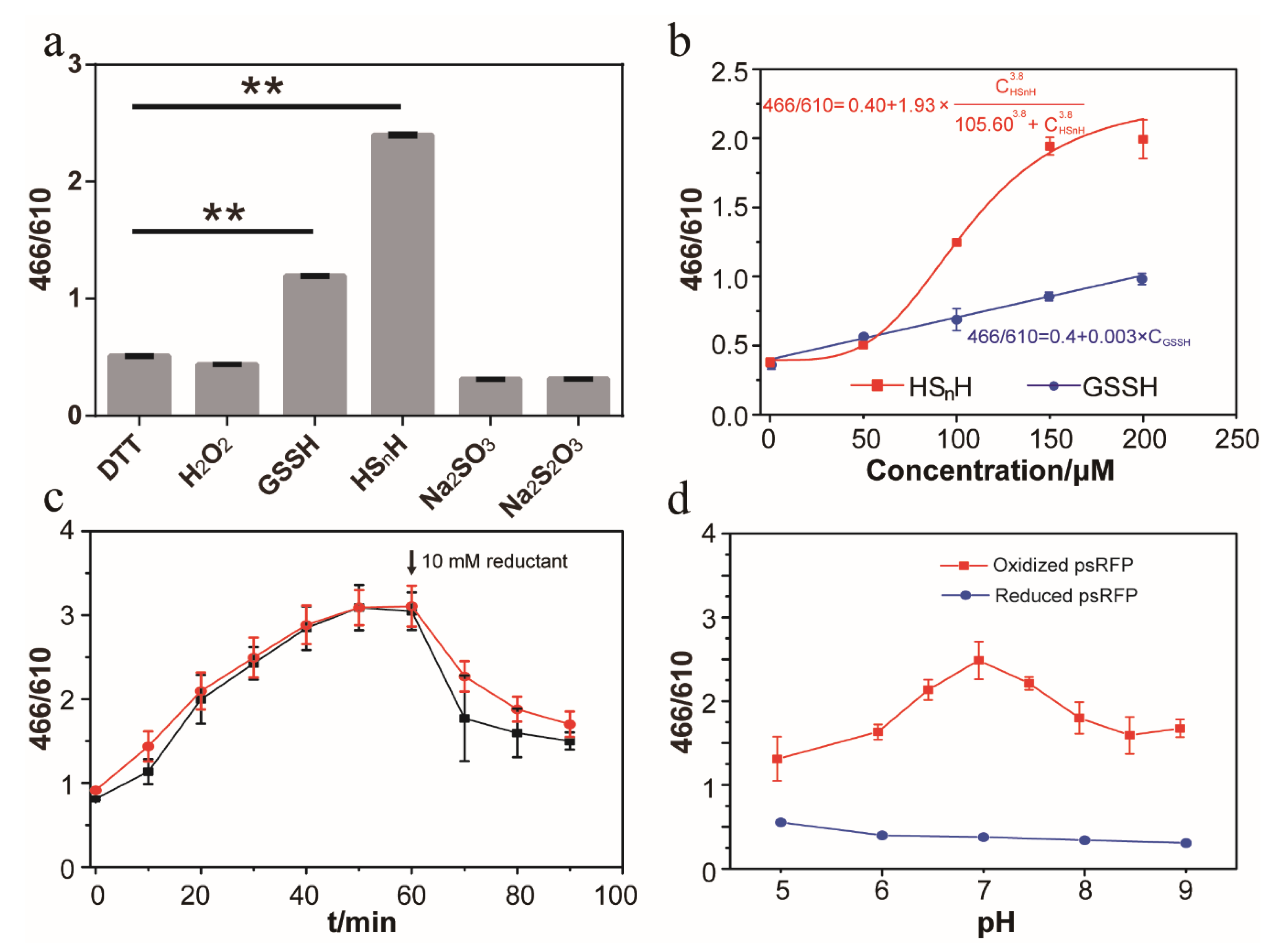
3.3. In Vitro Test of psRFP
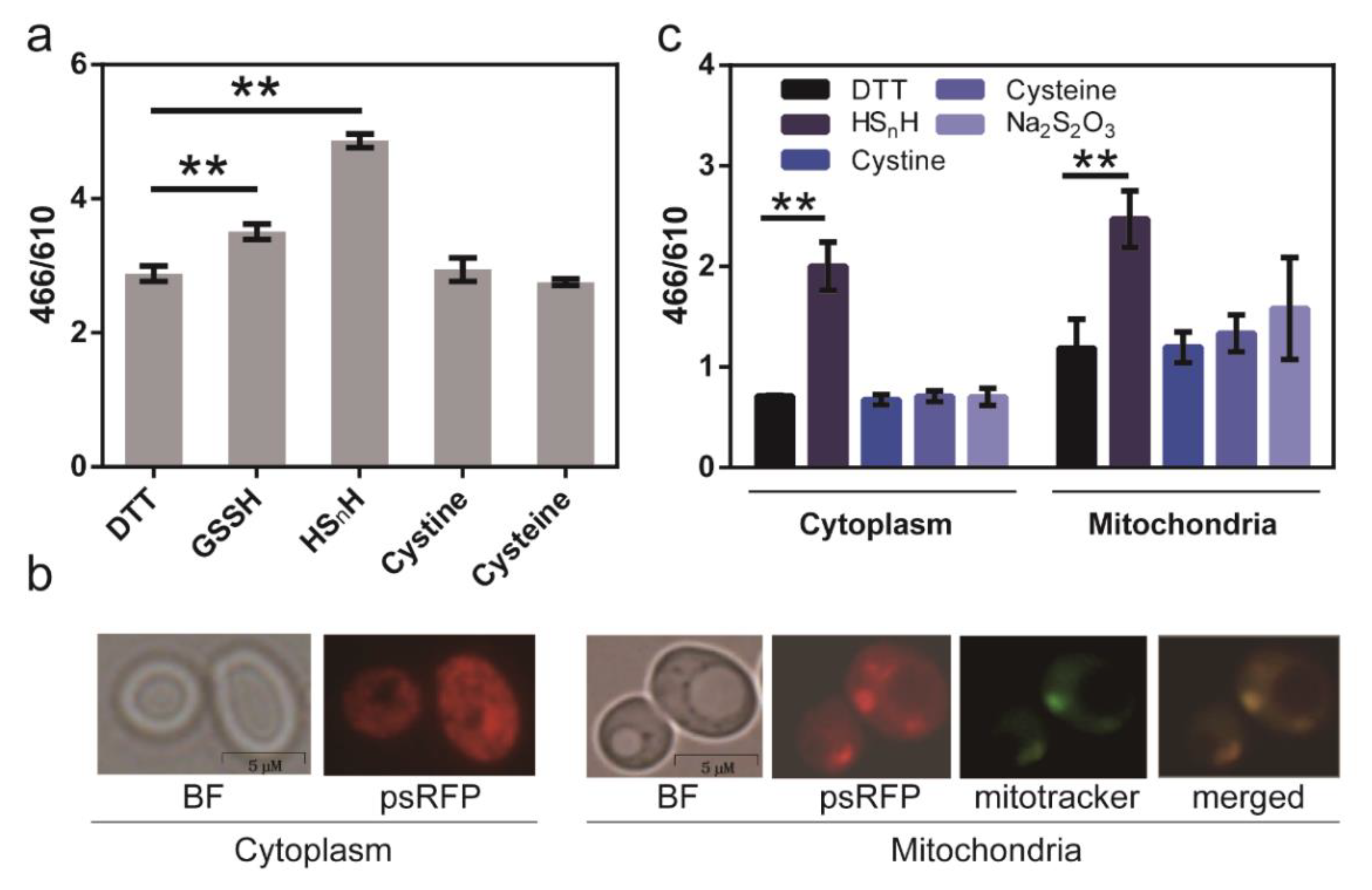
3.4. In Vivo Test of psRFP
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Giles, G.I.; Nasim, M.J.; Ali, W.; Jacob, C. The reactive sulfur species concept: 15 years on. Antioxidants 2017, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Fukuto, J.M.; Ignarro, L.J.; Nagy, P.; Wink, D.A.; Kevil, C.G.; Feelisch, M.; Cortese-Krott, M.M.; Bianco, C.L.; Kumagai, Y.; Hobbs, A.J.; et al. Biological hydropersulfides and related polysulfides–a new concept and perspective in redox biology. FEBS Lett. 2018, 592, 2140–2152. [Google Scholar] [CrossRef]
- Yadav, P.K.; Martinov, M.; Vitvitsky, V.; Seravalli, J.; Wedmann, R.; Filipovic, M.R.; Banerjee, R. Biosynthesis and reactivity of cysteine persulfides in signaling. J. Am. Chem. Soc. 2016, 138, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Yadav, V.; Banerjee, R. Heme-dependent metabolite switching regulates H2S synthesis in response to endoplasmic reticulum (ER) stress. J. Biol. Chem. 2016, 291, 16418–16423. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Enzymology of H2S biogenesis, decay and signaling. Antioxid. Redox Sign. 2014, 20, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Walsh, B.J.C.; Giedroc, D.P. H2S and reactive sulfur signaling at the host bacterial pathogen interface. J. Biol. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, H.; Chen, Z.; Zhao, R.; Wang, Q.; Ran, M.; Xia, Y.; Hu, X.; Liu, J.; Xian, M.; et al. Using resonance synchronous spectroscopy to characterize the reactivity and electrophilicity of biologically relevant sulfane sulfur. Redox Biol. 2019, 24. [Google Scholar] [CrossRef]
- Ono, K.; Akaike, T.; Sawa, T.; Kumagai, Y.; Wink, D.A.; Tantillo, D.J.; Hobbs, A.J.; Nagy, P.; Xian, M.; Lin, J.; et al. Redox chemistry and chemical biology of H2S, hydropersulfides, and derived species: Implications of their possible biological activity and utility. Free Radic. Biol. Med. 2014, 77, 82–94. [Google Scholar] [CrossRef]
- Kimura, H. Signaling molecules: Hydrogen sulfide and polysulfide. Antioxid. Redox Sign. 2015, 22, 362–376. [Google Scholar] [CrossRef]
- Mishanina, T.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef]
- Kimura, H. Signalling by hydrogen sulfide and polysulfides via protein S-sulfuration. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M.; Kuhnle, G.G.C.; Dyson, A.; Fernandez, B.O.; Grman, M.; DuMond, J.F.; Barrow, M.P.; McLeod, G.; Nakagawa, H.; Ondrias, K.; et al. Key bioactive reaction products of the NO/H2S interaction are S/N-hybrid species, polysulfides, and nitroxyl. Proc. Natl. Acad. Sci. USA. 2015, 112, E4651–E4660. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, H.; Zhang, X.; Chen, Z.; Zhao, R.; Hou, N.; Liu, J.; Xun, L.; Liu, H. Developing polysulfide-sensitive gfps for real-time analysis of polysulfides in live cells and subcellular organelles. Anal. Chem. 2019, 91, 3893–3901. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A. Peroxisome Metabolism in cancer. Cells 2020, 9, 1692. [Google Scholar] [CrossRef] [PubMed]
- Brady, N.R.; Hamacher-Brady, A.; Westerhoff, H.V.; Gottlieb, R.A. A wave of reactive oxygen species (ROS)-induced ROS release in a sea of excitable mitochondria. Antioxid. Redox Sign. 2006, 8, 1651–1665. [Google Scholar] [CrossRef]
- Akaike, T.; Ida, T.; Wei, F.Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S.; et al. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Hussain, S.; Malik, A.H.; Iyer, P.K. FRET-assisted selective detection of flavins via cationic conjugated polyelectrolyte under physiological conditions. J. Mater. Chem. B 2016, 4, 4439–4446. [Google Scholar] [CrossRef]
- Bergman, L.W. Growth and maintenance of yeast. Methods Mol. Biol. 2001, 177, 9–14. [Google Scholar] [CrossRef]
- Xin, Y.; Liu, H.; Cui, F.; Liu, H.; Xun, L. Recombinant Escherichia coli with sulfide:quinone oxidoreductase and persulfide dioxygenase rapidly oxidises sulfide to sulfite and thiosulfate via a new pathway. Environ. Microbiol. 2016, 18, 5123–5136. [Google Scholar] [CrossRef]
- Luebke, J.L.; Shen, J.; Bruce, K.E.; Kehl-Fie, T.E.; Peng, H.; Skaar, E.P.; Giedroc, D.P. The CsoR-like sulfurtransferase repressor (CstR) is a persulfide sensor in Staphylococcus aureus. Mol. Microbiol. 2014, 94, 1343–1360. [Google Scholar] [CrossRef]
- Xia, Y.; Chu, W.; Qi, Q.; Xun, L. New insights into the QuikChangeTM process guide the use of Phusion DNA polymerase for site-directed mutagenesis. Nucl. Acids Res. 2015, 43, e12. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Lü, C.; Xia, Y.; Xin, Y.; Liu, H.; Xun, L.; Liu, H. FisR activates σ54-dependent transcription of sulfide-oxidizing genes in Cupriavidus pinatubonensis JMP134. Mol. Microbiol. 2017, 105, 373–384. [Google Scholar] [CrossRef]
- Bošnjak, I.; Bojović, V.; Šegvić-Bubić, T.S.; Bielen, A. Occurrence of protein disulfide bonds in different domains of life: A comparison of proteins from the Protein Data Bank. Protein Eng. Des. Sel. 2014, 27, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Merzlyak, E.M.; Goedhart, J.; Shcherbo, D.; Bulina, M.E.; Shcheglov, A.S.; Fradkov, A.F.; Gaintzeva, A.; Lukyanov, K.A.; Lukyanov, S.; Gadella, T.W.J.; et al. Bright monomeric red fluorescent protein with an extended fluorescence lifetime. Nat. Methods 2007, 4, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.G.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Mou, Z.; Wang, H.; Tang, X.; Dong, Z.; Wang, L.; Dong, X.; Liu, W. Highly selective and sensitive one- and two-photon ratiometric fluorescent probe for intracellular hydrogen polysulfide sensing. Anal. Chem. 2016, 88, 7206–7212. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, C.; Peng, B.; Zhao, Y.; Pacheco, A.; Xian, M. New fluorescent probes for sulfane sulfurs and the application in bioimaging. Chem. Sci. 2013, 4, 2892–2896. [Google Scholar] [CrossRef]
- Umezawa, K.; Kamiya, M.; Urano, Y. A reversible fluorescent probe for real-time live-cell imaging and quantification of endogenous hydropolysulfides. Angew. Chem. Int. Ed. Engl. 2018, 57, 9346–9350. [Google Scholar] [CrossRef]
- Takano, Y.; Hanaoka, K.; Shimamoto, K.; Miyamotoet, R.; Komatsu, T.; Ueno, T.; Terai, T.; Kimura, H.; Nagano, T.; Urano, Y. Development of a reversible fluorescent probe for reactive sulfur species, sulfane sulfur, and its biological application. Chem. Commun. 2017, 53, 1064–1067. [Google Scholar] [CrossRef]
- Gao, M.; Wang, R.; Yu, F.; You, J.; Chen, L. Imaging and evaluation of sulfane sulfur in acute brain ischemia using a mitochondria-targeted near-infrared fluorescent probe. J. Mater. Chem. B. 2018, 6, 2608–2619. [Google Scholar] [CrossRef]
- Han, X.; Song, X.; Li, B.; Yu, F.; Chen, L. A near-infrared fluorescent probe for sensitive detection and imaging of sulfane sulfur in living cells and in vivo. Biomater. Sci. 2018, 6, 672–682. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Characteristics/Purposes | References |
|---|---|---|
| Strains | ||
| S. cerevisiae BY4742 | MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 | A gift from Prof. Jin Hou |
| E. coli DH5a | supE44, AlacU169 (q80lacZAM15), hsdR17, recA1, endA1, gyrA96,thi-1, relA1. For cloning and plasmid construction | Lab stock |
| E. coli BL21(DE3) | F-ompT hsdSB (rB-mB-) gal ( λ1857 ind1 Sam7 nin5 lacUV5 T7gene1) dcm. For expression and protein purification | Lab stock |
| Plasmids | ||
| YEplac195-psGFPcyt | YEplac195 containing psRFP. Under control of TEF1 promoter | This study |
| YEplac195-psGFPmit | YEplac195 containing an N-ternimus sigal peptide a-psRFP. Under control of TEF1 promoter | This study |
| pET30a-psRFP | pET30a containing psRFP. Under control of T7 promoter for expression and protein purification | This study |
| pET30a-mCherry | pET30a containing mCherry. Under control of T7 promoter for expression and protein purification | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Wang, Q.; Xia, Y.; Xun, L.; Liu, H. A Red Fluorescent Protein-Based Probe for Detection of Intracellular Reactive Sulfane Sulfur. Antioxidants 2020, 9, 985. https://doi.org/10.3390/antiox9100985
Li Z, Wang Q, Xia Y, Xun L, Liu H. A Red Fluorescent Protein-Based Probe for Detection of Intracellular Reactive Sulfane Sulfur. Antioxidants. 2020; 9(10):985. https://doi.org/10.3390/antiox9100985
Chicago/Turabian StyleLi, Zimai, Qingda Wang, Yongzhen Xia, Luying Xun, and Huaiwei Liu. 2020. "A Red Fluorescent Protein-Based Probe for Detection of Intracellular Reactive Sulfane Sulfur" Antioxidants 9, no. 10: 985. https://doi.org/10.3390/antiox9100985
APA StyleLi, Z., Wang, Q., Xia, Y., Xun, L., & Liu, H. (2020). A Red Fluorescent Protein-Based Probe for Detection of Intracellular Reactive Sulfane Sulfur. Antioxidants, 9(10), 985. https://doi.org/10.3390/antiox9100985