Effect of Cooking Methods on the Antioxidant Capacity of Plant Foods Submitted to In Vitro Digestion–Fermentation

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.1.1. In Vitro Digestion and Fermentation
2.1.2. Antioxidant Capacity
2.2. Plant Foods and Cooking Conditions
2.3. In Vitro Gastrointestinal Digestion and Fermentation
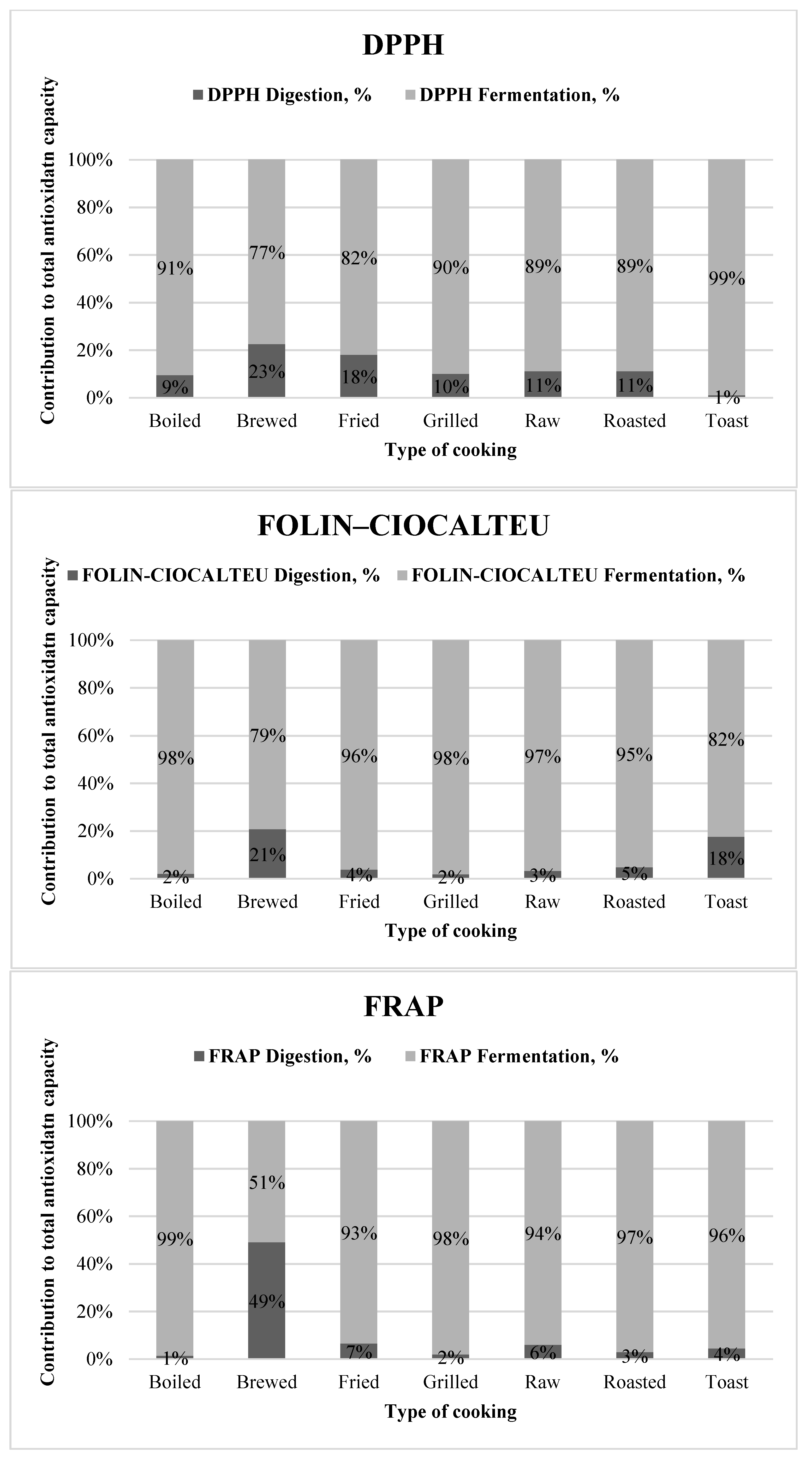
2.4. Antioxidant Assays
2.5. Calculations of Daily Antioxidant Intake
2.6. Statistical Analysis
3. Results
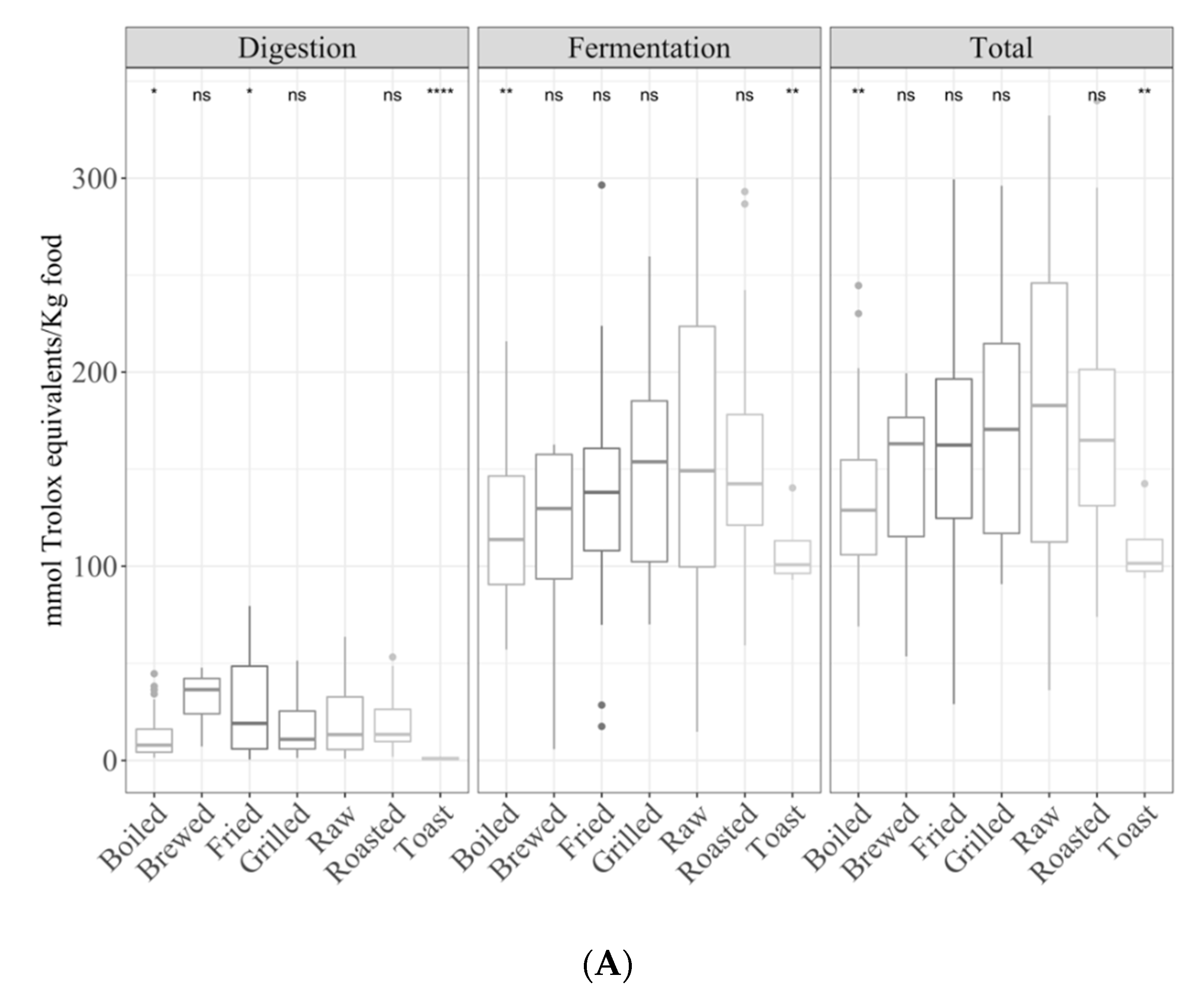
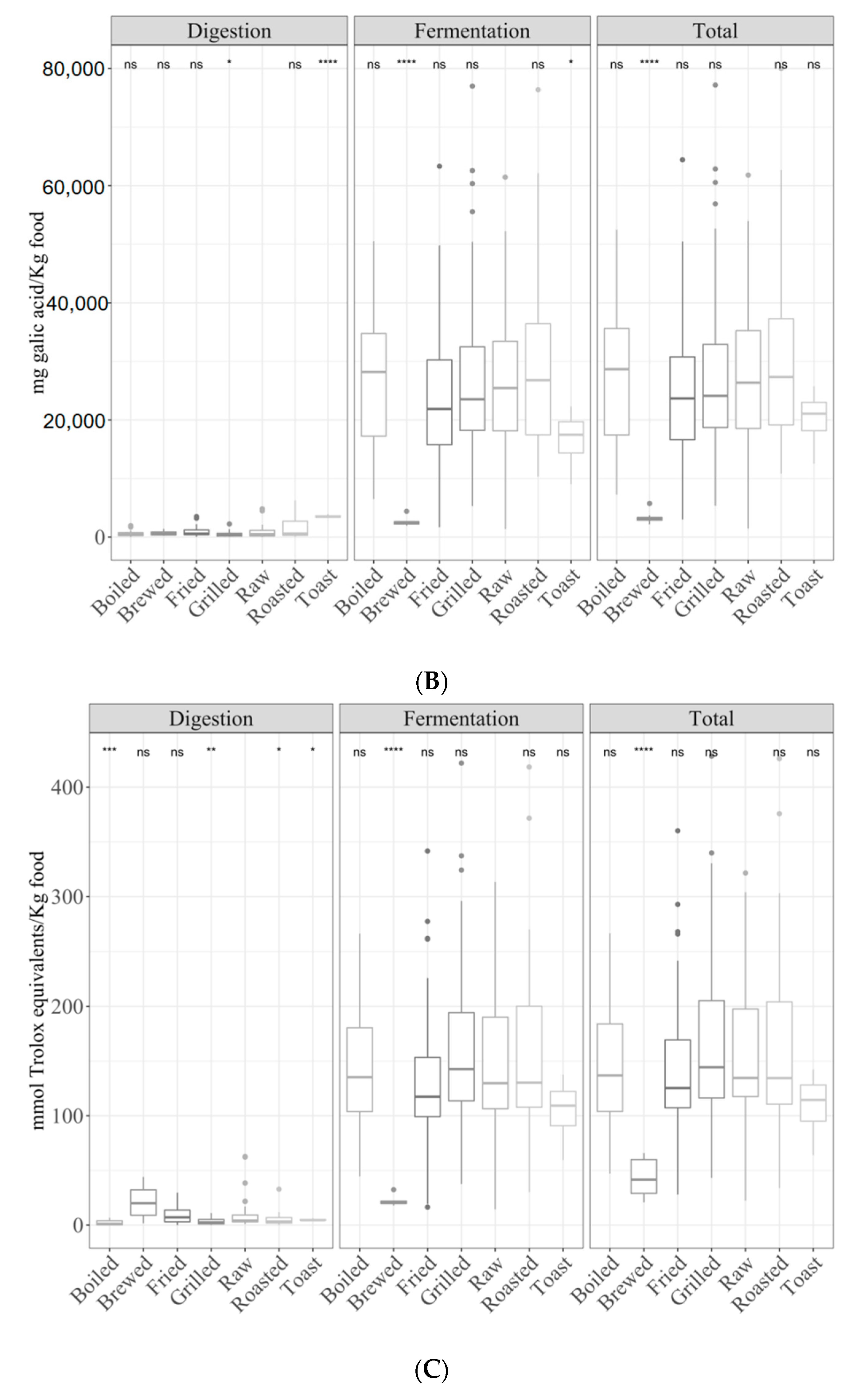
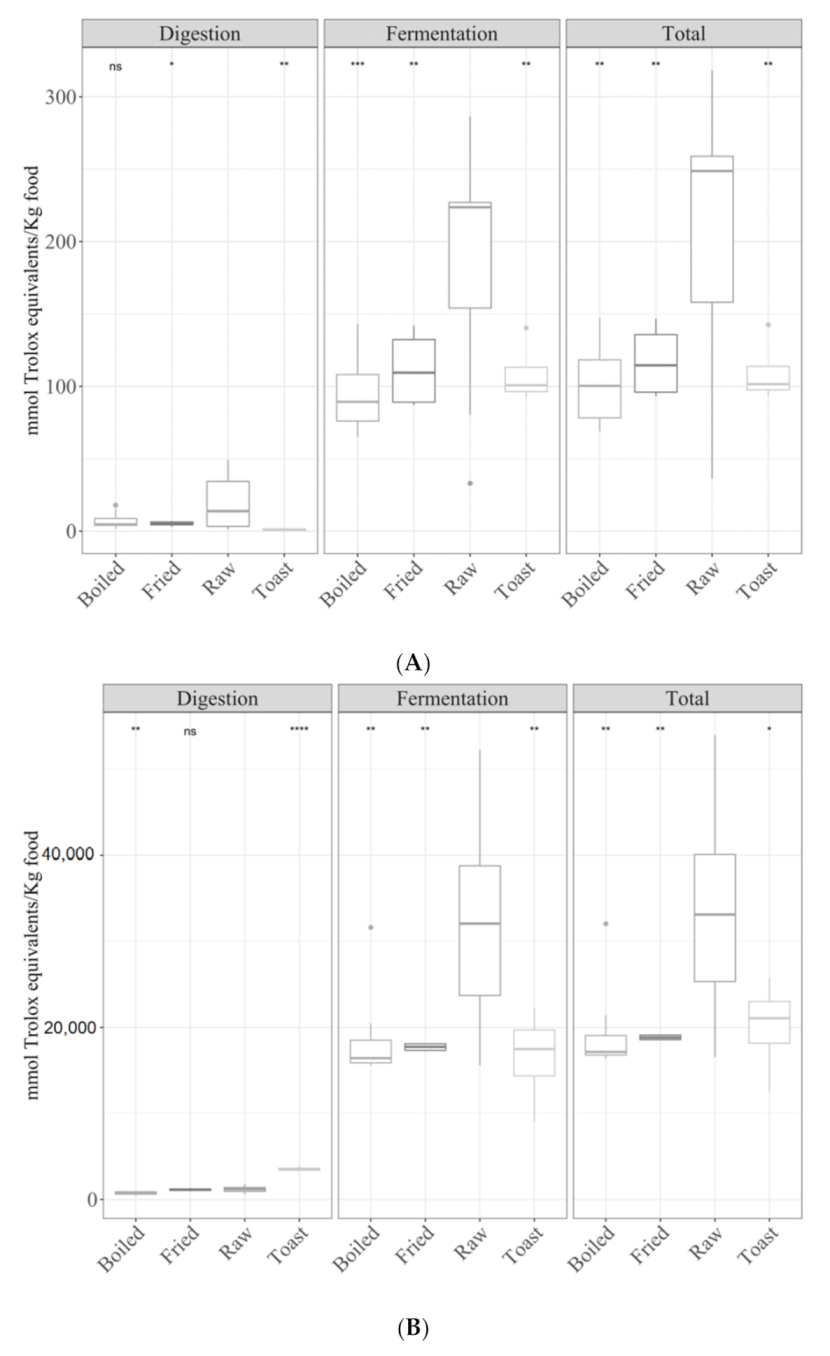
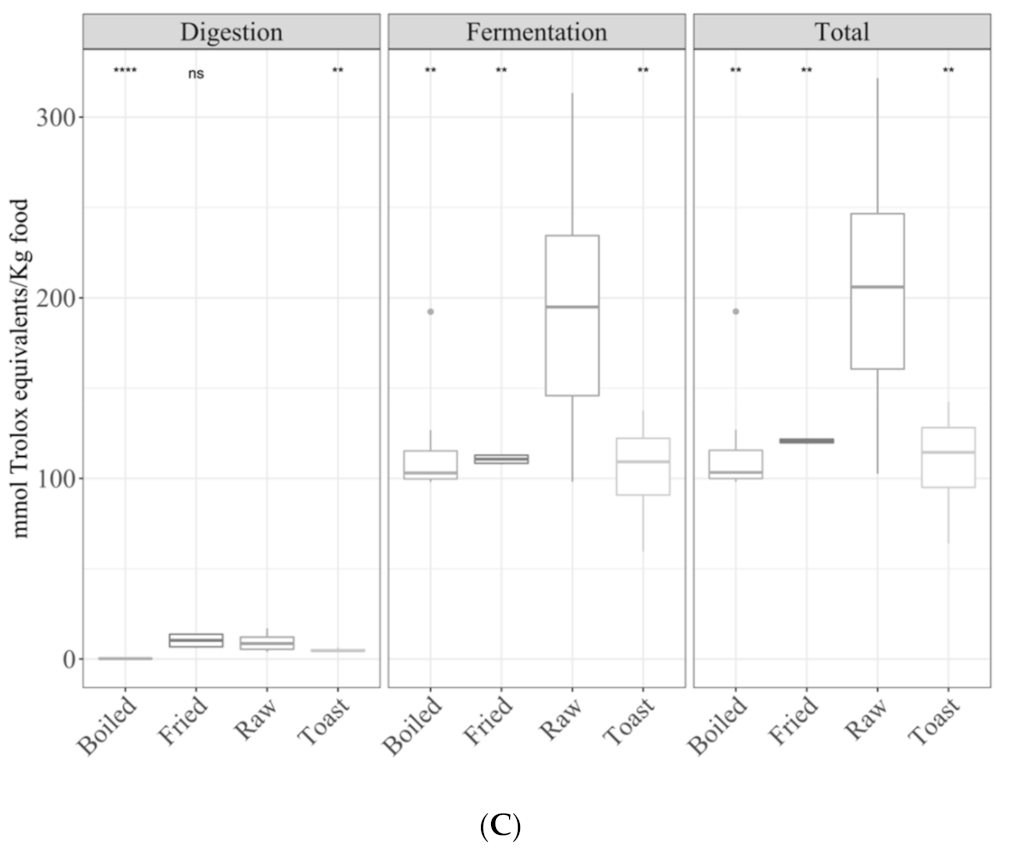
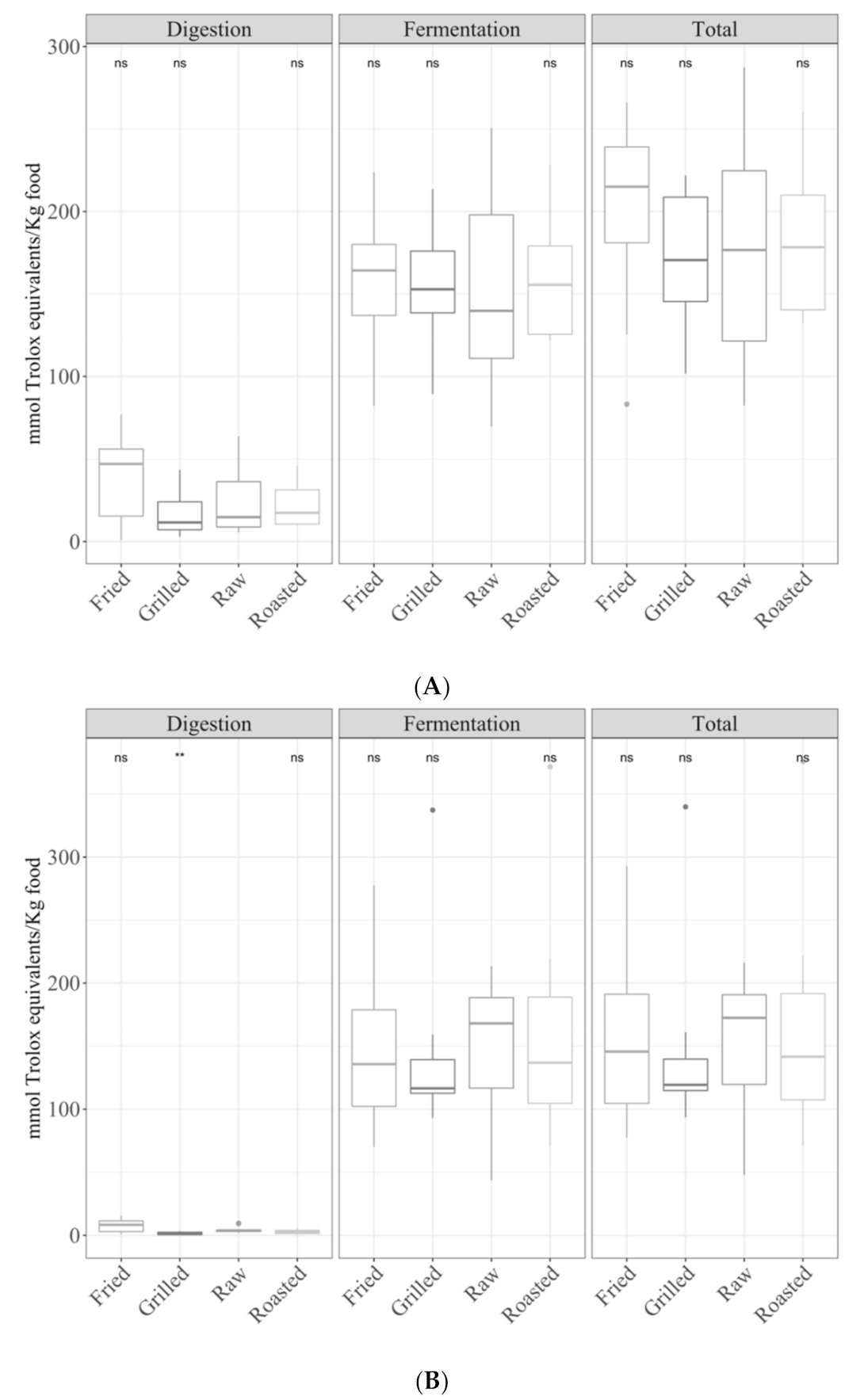
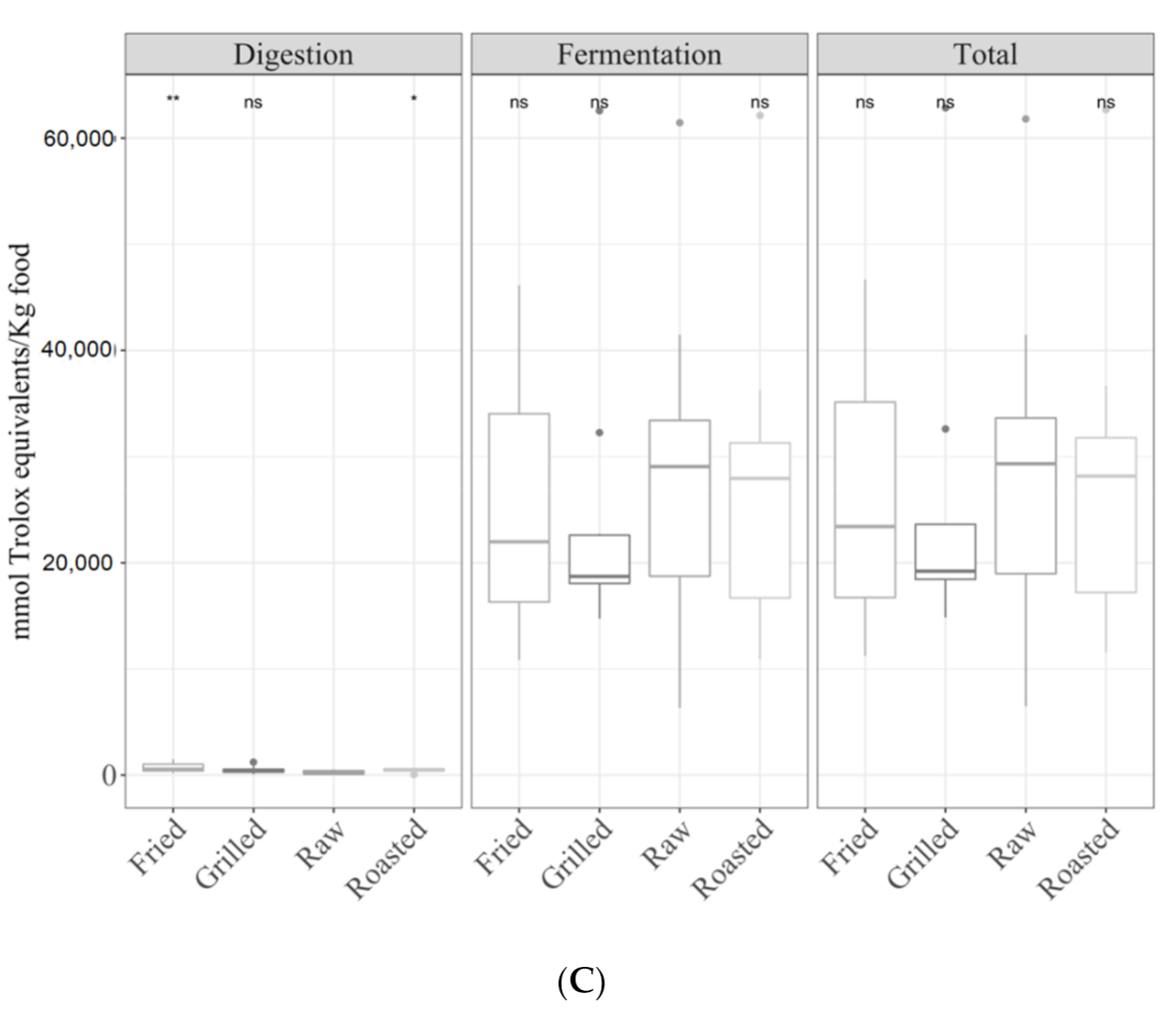
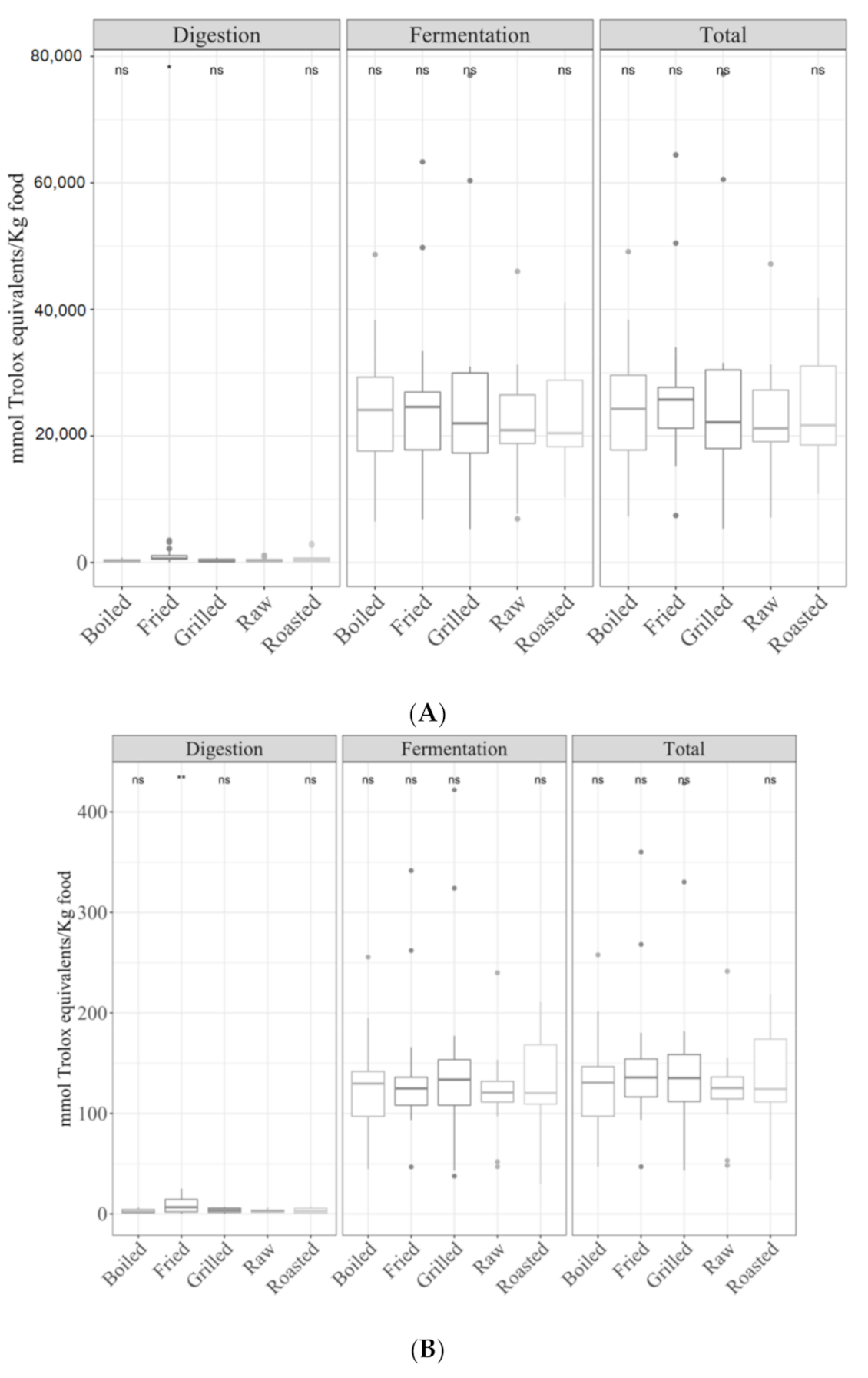
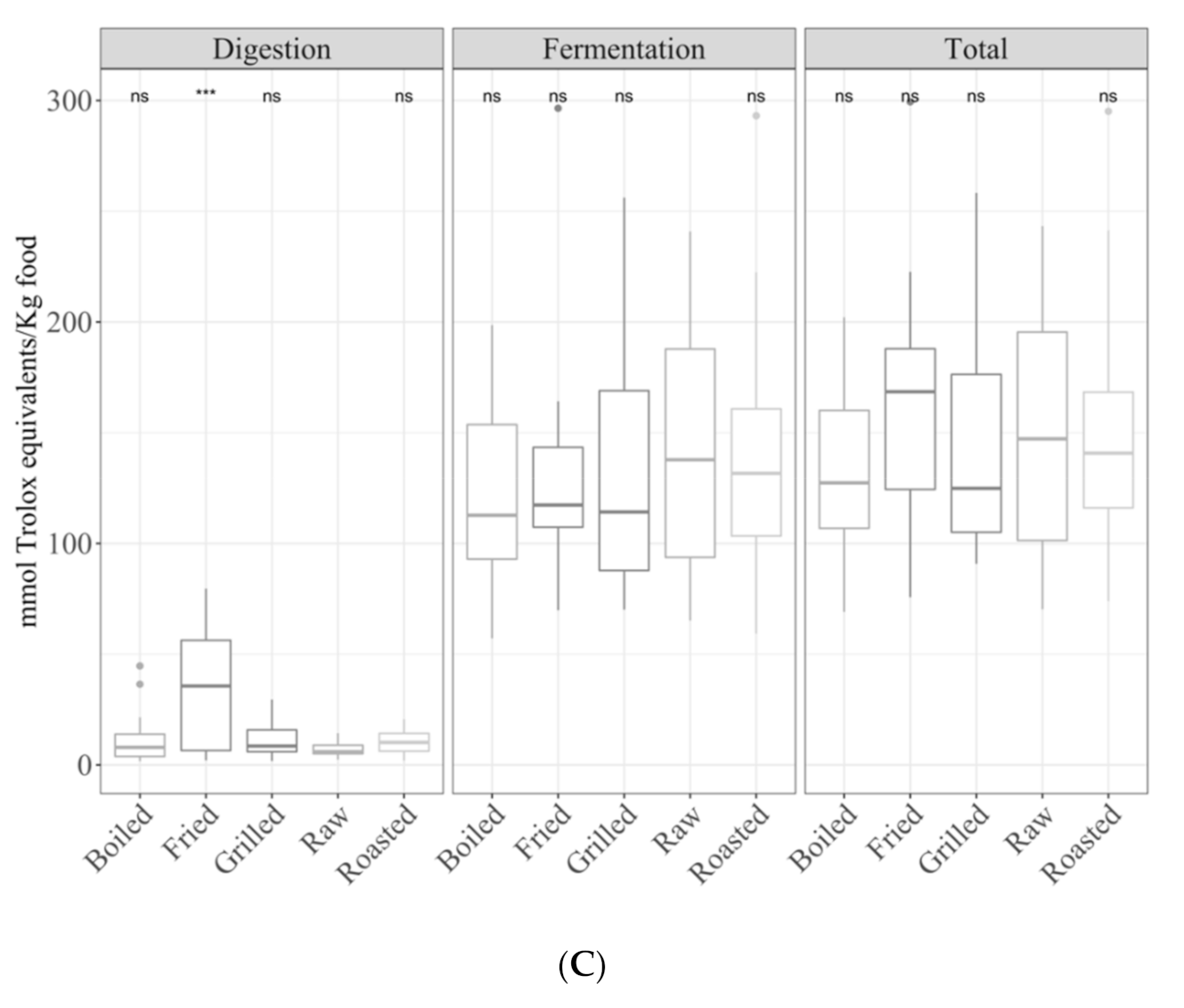
3.1. Samples by Type of Cooking
3.1.1. Gastrointestinal Digested Fraction
3.1.2. Gastrointestinal Fermented Fraction
3.1.3. Total Antioxidant Capacity
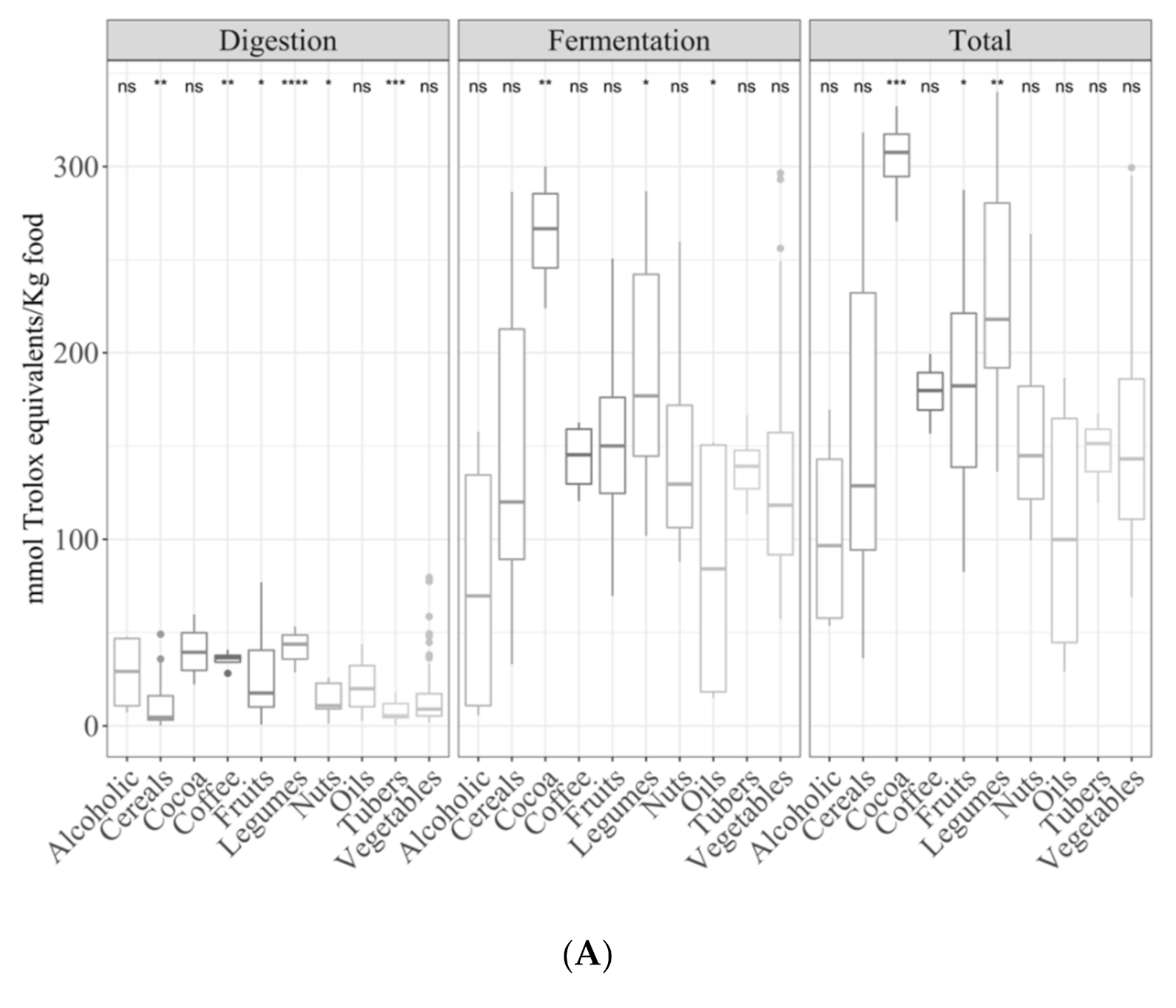
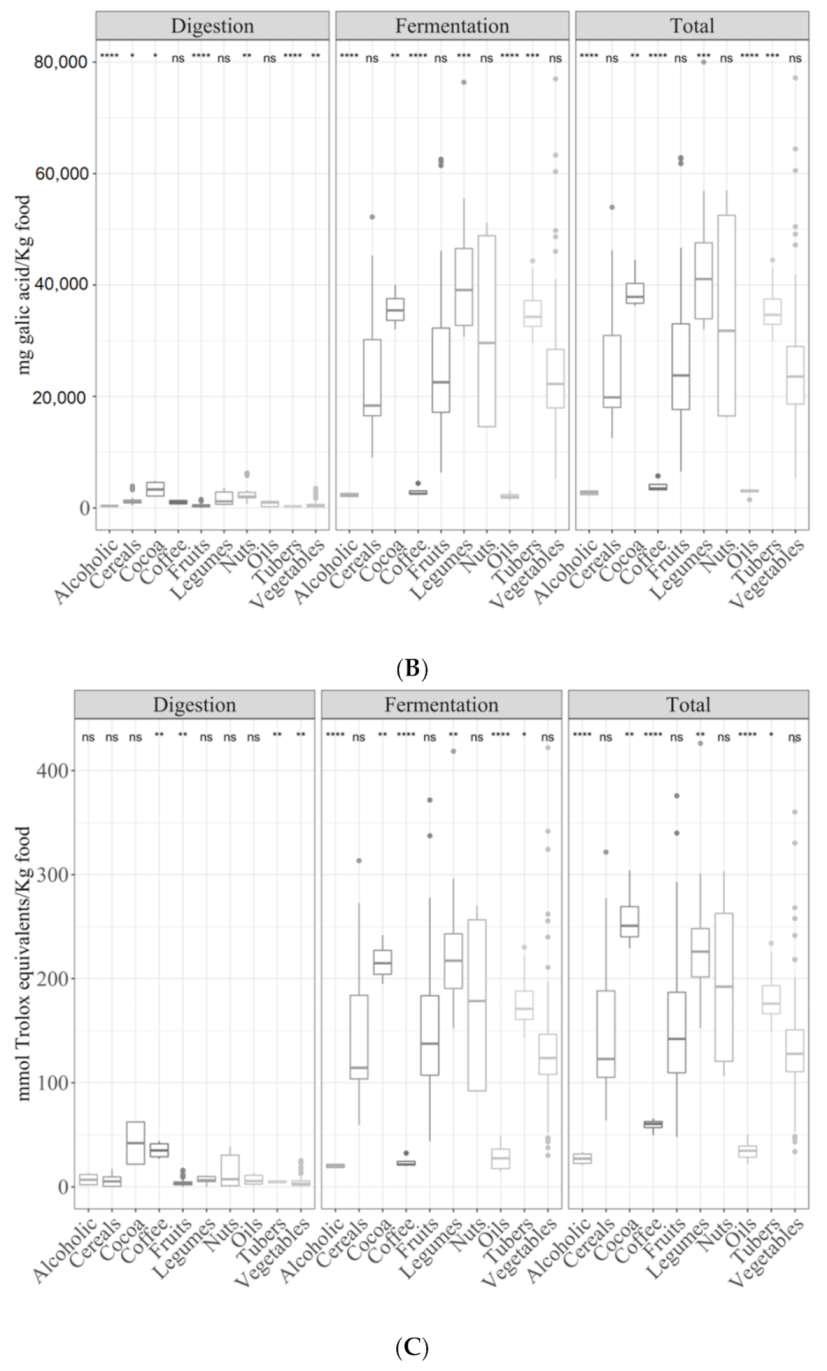
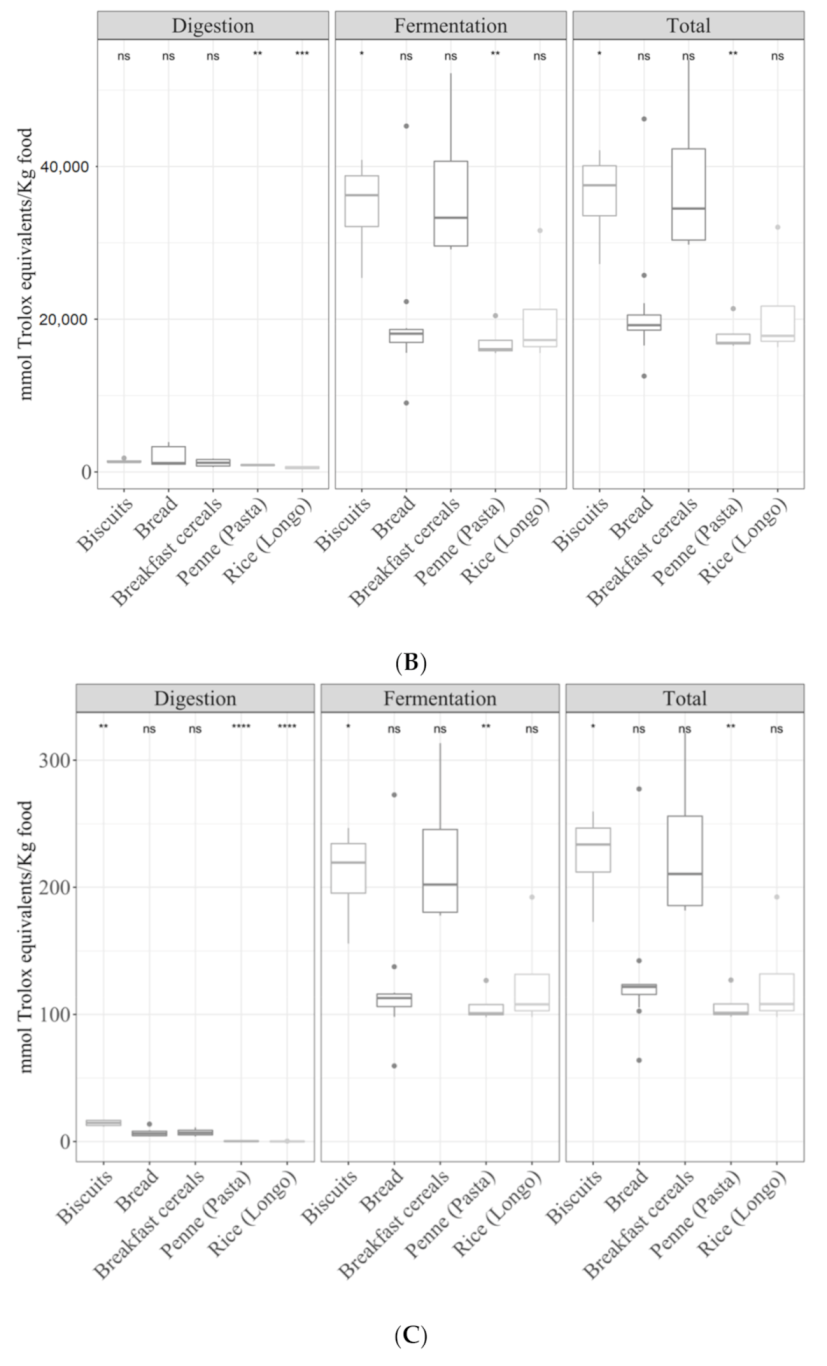
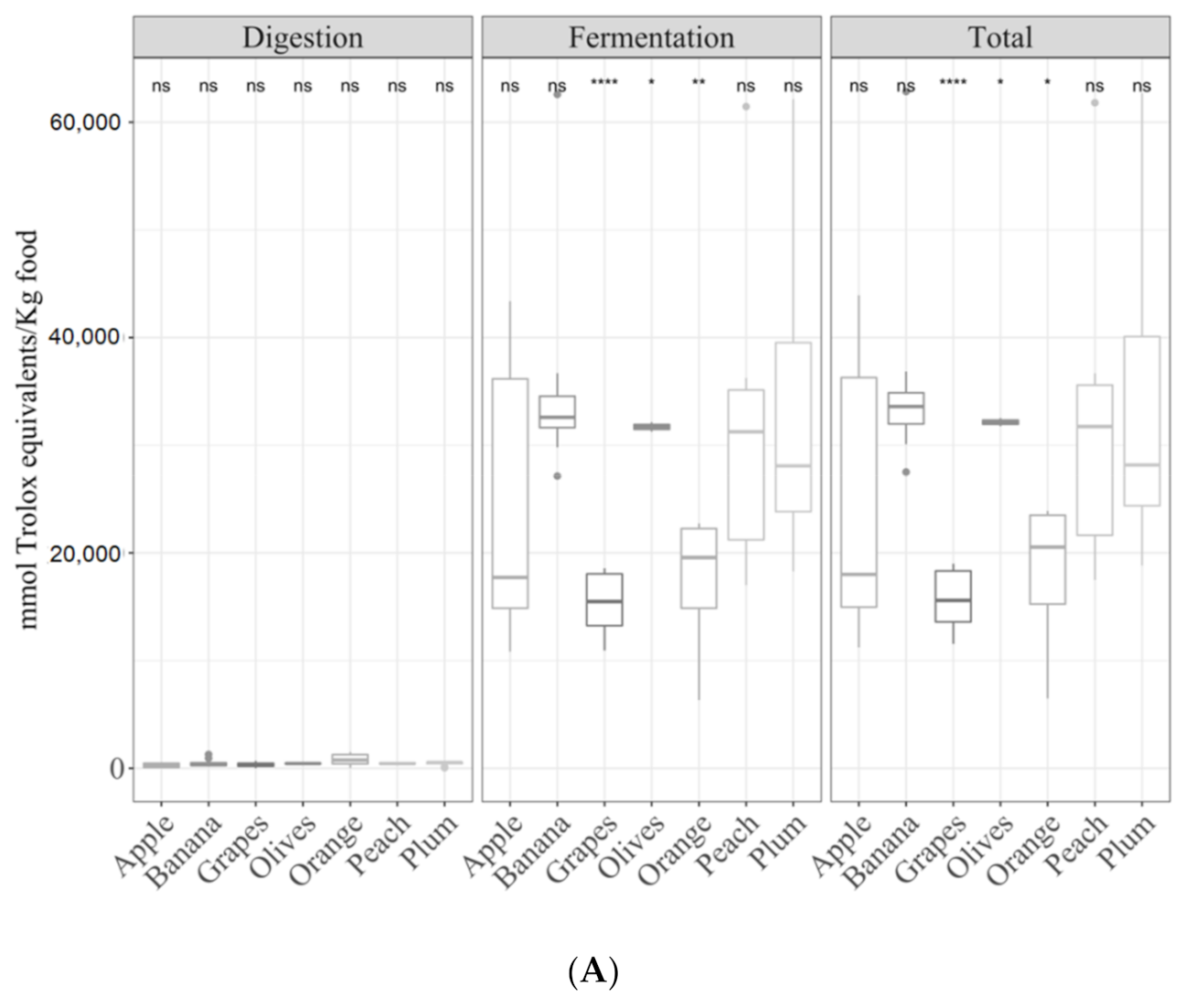
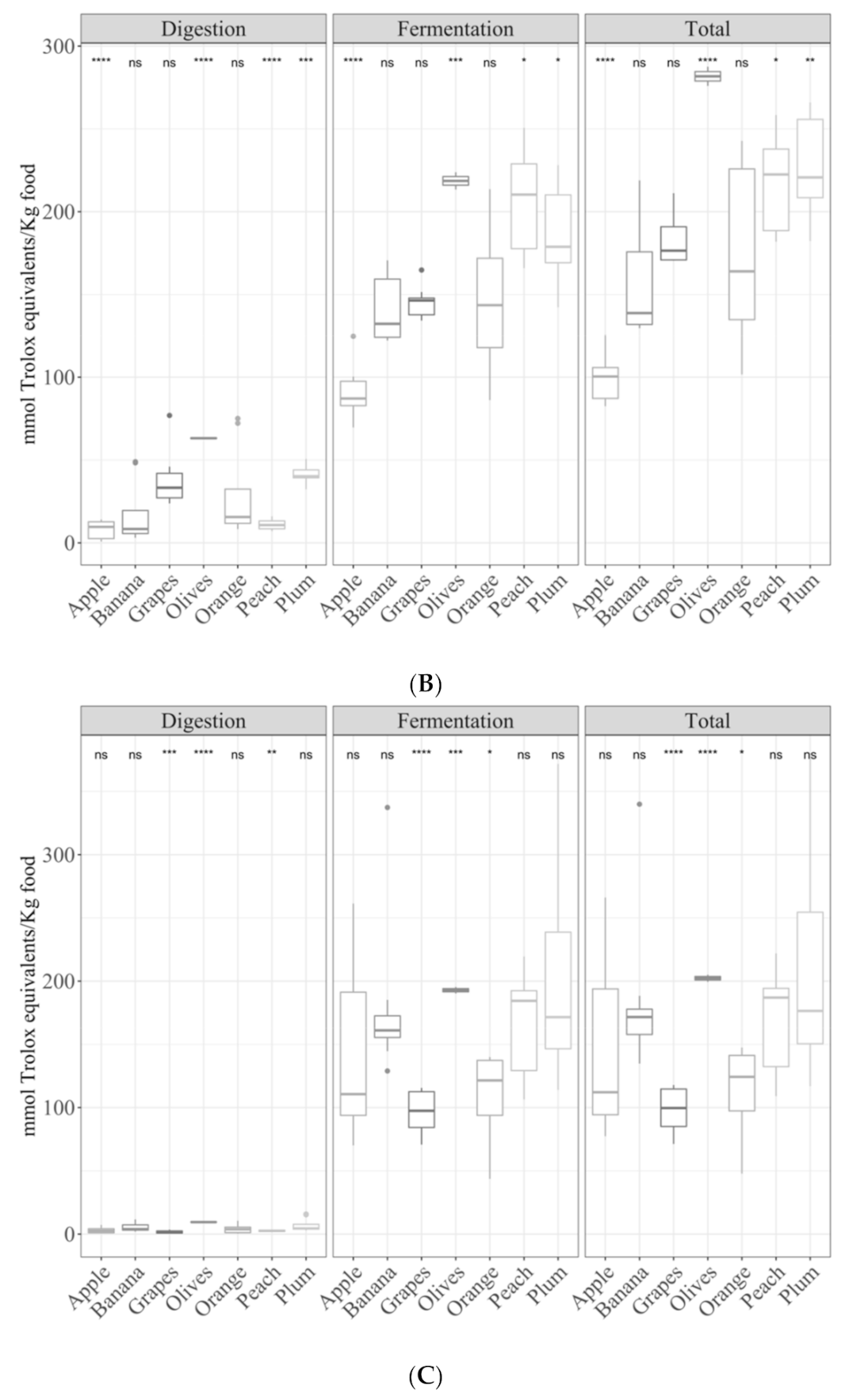
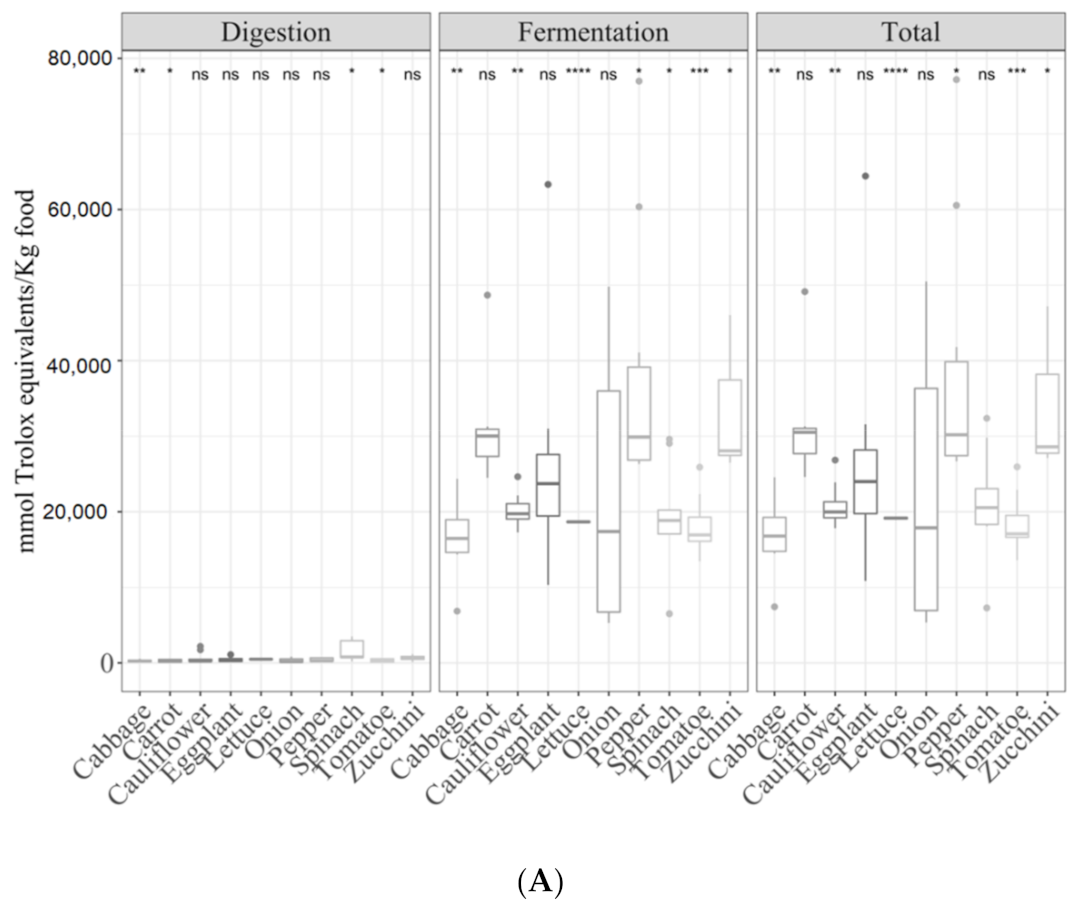
3.2. Samples by Type of Food
3.2.1. Gastrointestinal Digested Fraction
3.2.2. Gastrointestinal Fermented Fraction
3.2.3. Total Antioxidant Capacity
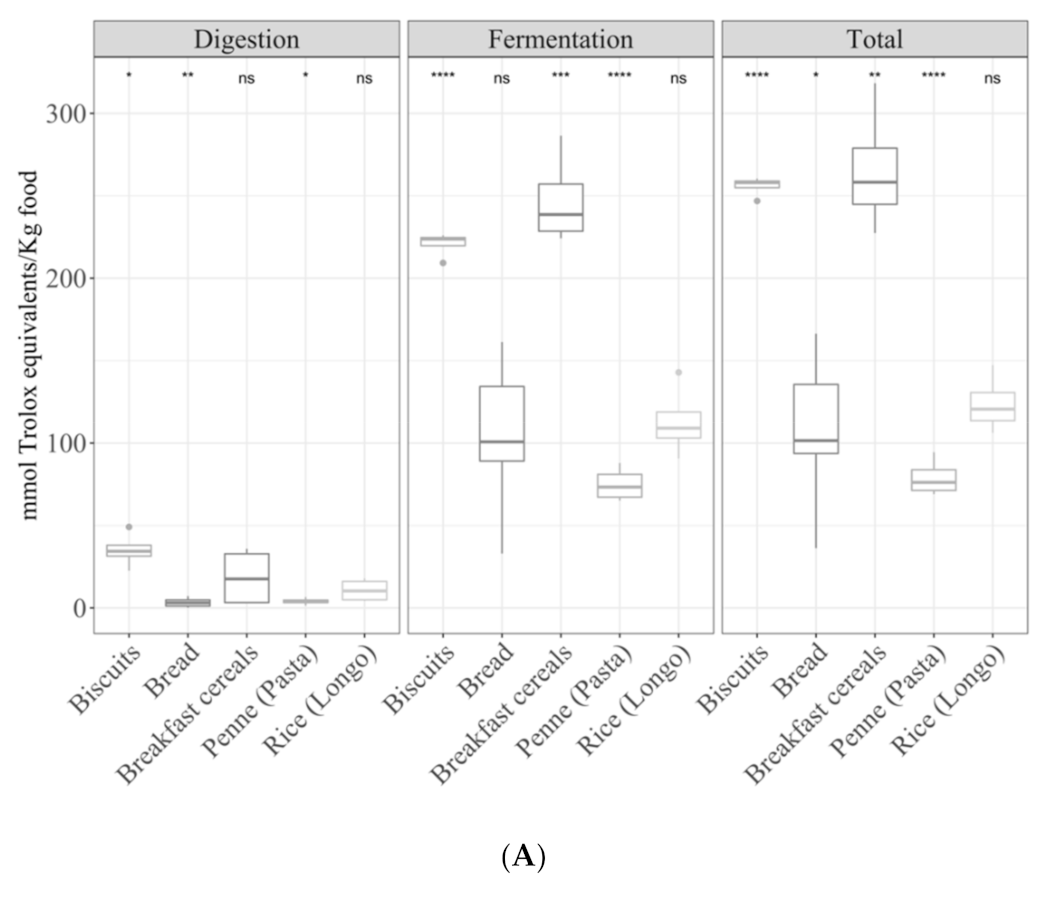
3.3. Detailed Analysis of Large Food Groups
3.3.1. Cereals
3.3.2. Fruits
3.3.3. Vegetables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abuajah, C.I.; Ogbonna, A.C.; Osuji, C.M. Functional components and medicinal properties of food: A review. J. Food Sci. Technol. 2015, 52, 2522–2529. [Google Scholar] [CrossRef] [Green Version]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.H. Dietary bioactive compounds and their health implications. J. Food Sci. 2013, 78, A18–A25. [Google Scholar] [CrossRef]
- Bello Gutierrez, J. Los Tipos de Coccion. In Ciencia y Tecnología Culinaria, 1st ed.; Bello Gutierrez, J., Ed.; Díaz de Santos S.A.: Madrid, Spain, 1998; pp. 129–190. [Google Scholar]
- del Ramírez-Anaya, J.P.; Samaniego-Sánchez, C.; Castañeda-Saucedo, M.C.; Villalón-Mir, M.; de la Serrana, H.L.-G. Phenols and the antioxidant capacity of Mediterranean vegetables prepared with extra virgin olive oil using different domestic cooking techniques. Food Chem. 2015, 188, 430–438. [Google Scholar] [CrossRef]
- Miglio, C.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effects of different cooking methods on nutritional and physicochemical characteristics of selected vegetables. J. Agric. Food Chem. 2008, 56, 139–147. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Guerra-Hernández, E.J.; García-Villanova, B. Colour measurement as indicator for controlling the manufacture and storage of enteral formulas. Food Cont. 2006, 17, 489–493. [Google Scholar] [CrossRef]
- Pastoriza de la Cueva, S.; Álvarez, J.; Végvári, Á.; Montilla-Gómez, J.; Cruz-López, O.; Delgado-Andrade, C.; Rufián-Henares, J.A. Relationship between HMF intake and SMF formation in vivo: An animal and human study. Mol. Nutr. Food Res. 2017, 61, 1600773. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Guerra-Hernández, E.; García-Villanova, B. Effect of red sweet pepper dehydration conditions on Maillard reaction, ascorbic acid and antioxidant activity. J. Food Eng. 2013, 118, 150–156. [Google Scholar] [CrossRef]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet–microbiota interactions and personalized nutrition. Nat. Rev. Microbiol. 2019, 17, 742–753. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Burillo, S.; Rufián-Henares, J.A.; Pastoriza, S. Towards an improved global antioxidant response method (GAR+): Physiological-resembling in vitro digestion-fermentation method. Food Chem. 2018, 239, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Pastoriza, S.; Delgado-Andrade, C.; Haro, A.; Rufián-Henares, J.A. A physiologic approach to test the global antioxidant response of foods. The GAR method. Food Chem. 2011, 129, 1926–1932. [Google Scholar] [CrossRef]
- Yen, G.-C.; Chen, H.-Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Moreno-Montoro, M.; Olalla-Herrera, M.; Gimenez-Martinez, R.; Navarro-Alarcon, M.; Rufián-Henares, J.A. Phenolic compounds and antioxidant activity of Spanish commercial grape juices. J. Food Comp. Anal. 2015, 38, 19–26. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a measure of “Antioxidant Power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Mercasa. La Alimentación en España. 2020. Available online: https://www.mercasa.es/media/publicaciones/281/AEE_2020_web.pdf (accessed on 12 November 2020).
- Salvador i Castells, G. Tabla de Medidas Caseras de Alimentos. In Nutrición y Dietética Clínica; Salas-Salvadó, J., Bonada, A., Trallero, R., Saló, M.E., Eds.; Elsevier-Masson: Barcelona, Spain, 2000; pp. 557–570. [Google Scholar]
- Saura-Calixto, F.; Goñi, I. Antioxidant capacity of the Spanish Mediterranean diet. Food Chem. 2006, 94, 442–447. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Rufián-Henares, J.A.; Pastoriza, S. Towards an improved Global Antioxidant Response method (GAR+): Physiological-resembling in vitro antioxidant capacity methods. Food Chem. 2018, 239, 1263–1272. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Correia, E.; Lopes, L.; Guido, L.F. Further insights into the role of melanoidins on the antioxidant potential of barley malt. Food Chem. 2014, 160, 127–133. [Google Scholar] [CrossRef]
- Pastoriza, S.; Seiquer, I.; Mesías, M.; Rufián Henares, J.A.; Delgado Andrade, C. Evaluation of the availability and antioxidant capacity of Maillard compounds present in bread crust: Studies in Caco-2 cells. Foods 2017, 6, 5. [Google Scholar] [CrossRef]
- Dittrich, R.; Dragonas, C.; Kannenkeril, D.; Hoffmann, I.; Mueller, A.; Beckmann, M.W.; Pischetsrieder, M. A diet rich in Maillard reaction products protects LDL against copper induced oxidation ex vivo, a human intervention trial. Food Res. Int. 2009, 42, 1315–1322. [Google Scholar] [CrossRef]
- Martín, M.Á.; Ramos, S.; Mateos, R.; Rufián-Henares, J.A.; Morales, F.J.; Bravo, L.; Goya, L. Biscuit melanoidins of different molecular masses protect human HepG2 cells against oxidative stress. J. Agric. Food Chem. 2009, 57, 7250–7258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliazucchi, D.; Verzelloni, E.; Conte, A. Antioxidant properties of traditional balsamic vinegar and boiled must model systems. Eur. Food Res. Technol. 2007, 227, 835. [Google Scholar] [CrossRef]
- Ahmad-Qasem, M.H.; Cánovas, C.; Barrajó-Catalá, E.; Carreres, J.E.; Micol, V.; García-Peŕez, J.V. Influence of olive leaf processing on the bioaccessibility of bioactive polyphenols. J. Agric. Food Chem. 2014, 62, 6190–6198. [Google Scholar] [CrossRef] [PubMed]
- Plengmuankhae, W.; Tantitadapitak, C. Low temperature and water dehydration increase the levels of asiaticoside and madecassoside in Centella asiatica (L.) Urban. S. Afric. J. Bot. 2015, 97, 196–203. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Rufián-Henares, J.A.; Pastoriza, S. Effect of home cooking on the antioxidant capacity of vegetables: Relationship with Maillard reaction indicators. Food Res. Int. 2019, 121, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Bellail, A.A.; Shaltout, O.E.; Youssef, M.M.; Gamal, A.M.A.E. Effect of home-cooking methods on phenolic composition and antioxidant activity of sweetpotato (Ipomoea batatas (L.) Lam.) cultivars grown in Egypt. Food Nutr. Sci. 2012, 3, 720–726. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, F.A.; Nitzke, J.A.; Klipel, C.B.; Jong, E.V. de Chocolate and red wine—A comparison between flavonoids content. Food Chem. 2010, 120, 109–112. [Google Scholar] [CrossRef]
- Montreau, F.R. Sur le dosage des composés phénoliques totaux dans les vins par la méthode Folin-Ciocalteu. OENO ONE 1972, 6, 397–404. [Google Scholar] [CrossRef]
- Lee, K.W.; Kim, Y.J.; Lee, H.J.; Lee, C.Y. Cocoa has more phenolic phytochemicals and a higher antioxidant capacity than teas and red wine. J. Agric. Food Chem. 2003, 51, 7292–7295. [Google Scholar] [CrossRef]
- Pastoriza, S.; Rufián-Henares, J.A. Contribution of melanoidins to the antioxidant capacity of the Spanish diet. Food Chem. 2014, 164, 438–445. [Google Scholar] [CrossRef]
- Durante, M.; Tufariello, M.; Tomassi, L.; Lenucci, M.L.; Bleve, G.; Mita, G. Evaluation of bioactive compounds in black table olives fermented with selected microbial starters. J. Sci. Food Agric. 2018, 98, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Lenucci, M.S.; Gazza, L.; Taddei, F.; Nocente, F.; De Benedetto, G.E.; De Caroli, M.; Piro, G.; Mita, G. Bioactive composition and sensory evaluation of innovative spaghetti supplemented with free or α-cyclodextrin chlatrated pumpkin oil extracted by supercritical CO2. Food Chem. 2019, 294, 112–122. [Google Scholar] [CrossRef] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food type | Analytical Assay | AOX Daily Intake 1 (µmol Trolox/d) | AOX Serving Intake 2 (µmol Trolox/serving) | Mean Contribution to Daily Antioxidant Capacity Intake (%) 3 | Mean Contribution to Daily Antioxidant Capacity Intake Per Serving (%) 3 |
|---|---|---|---|---|---|
| Alcoholic drinks | FRAP | 1228 | 3649 | 20.4 | 60.7 |
| Cereals | FRAP | 22,423 | 6343 | 373 | 106 |
| Cocoa | FRAP | 909 | 6888 | 15.1 | 115 |
| Coffee | FRAP | 296 | 1426 | 4.92 | 23.7 |
| Fruits | FRAP | 19,130 | 22,230 | 318 | 370 |
| Legumes | FRAP | 1250 | 16,655 | 20.8 | 277 |
| Nuts | FRAP | 1257 | 5808 | 20.9 | 96.6 |
| Oils | FRAP | 1157 | 348 | 19.2 | 5.8 |
| Tubers | FRAP | 14,804 | 31,253 | 246 | 520 |
| Vegetables | FRAP | 15,336 | 16,001 | 255 | 266 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navajas-Porras, B.; Pérez-Burillo, S.; Valverde-Moya, Á.J.; Hinojosa-Nogueira, D.; Pastoriza, S.; Rufián-Henares, J.Á. Effect of Cooking Methods on the Antioxidant Capacity of Plant Foods Submitted to In Vitro Digestion–Fermentation. Antioxidants 2020, 9, 1312. https://doi.org/10.3390/antiox9121312
Navajas-Porras B, Pérez-Burillo S, Valverde-Moya ÁJ, Hinojosa-Nogueira D, Pastoriza S, Rufián-Henares JÁ. Effect of Cooking Methods on the Antioxidant Capacity of Plant Foods Submitted to In Vitro Digestion–Fermentation. Antioxidants. 2020; 9(12):1312. https://doi.org/10.3390/antiox9121312
Chicago/Turabian StyleNavajas-Porras, Beatriz, Sergio Pérez-Burillo, Álvaro Jesús Valverde-Moya, Daniel Hinojosa-Nogueira, Silvia Pastoriza, and José Ángel Rufián-Henares. 2020. "Effect of Cooking Methods on the Antioxidant Capacity of Plant Foods Submitted to In Vitro Digestion–Fermentation" Antioxidants 9, no. 12: 1312. https://doi.org/10.3390/antiox9121312
APA StyleNavajas-Porras, B., Pérez-Burillo, S., Valverde-Moya, Á. J., Hinojosa-Nogueira, D., Pastoriza, S., & Rufián-Henares, J. Á. (2020). Effect of Cooking Methods on the Antioxidant Capacity of Plant Foods Submitted to In Vitro Digestion–Fermentation. Antioxidants, 9(12), 1312. https://doi.org/10.3390/antiox9121312